
Bioremediation of Petroleum Hydrocarbons in Seawater: Prospects of Using Lyophilized Native Hydrocarbon-Degrading Bacteria


 , , , and
, , , and 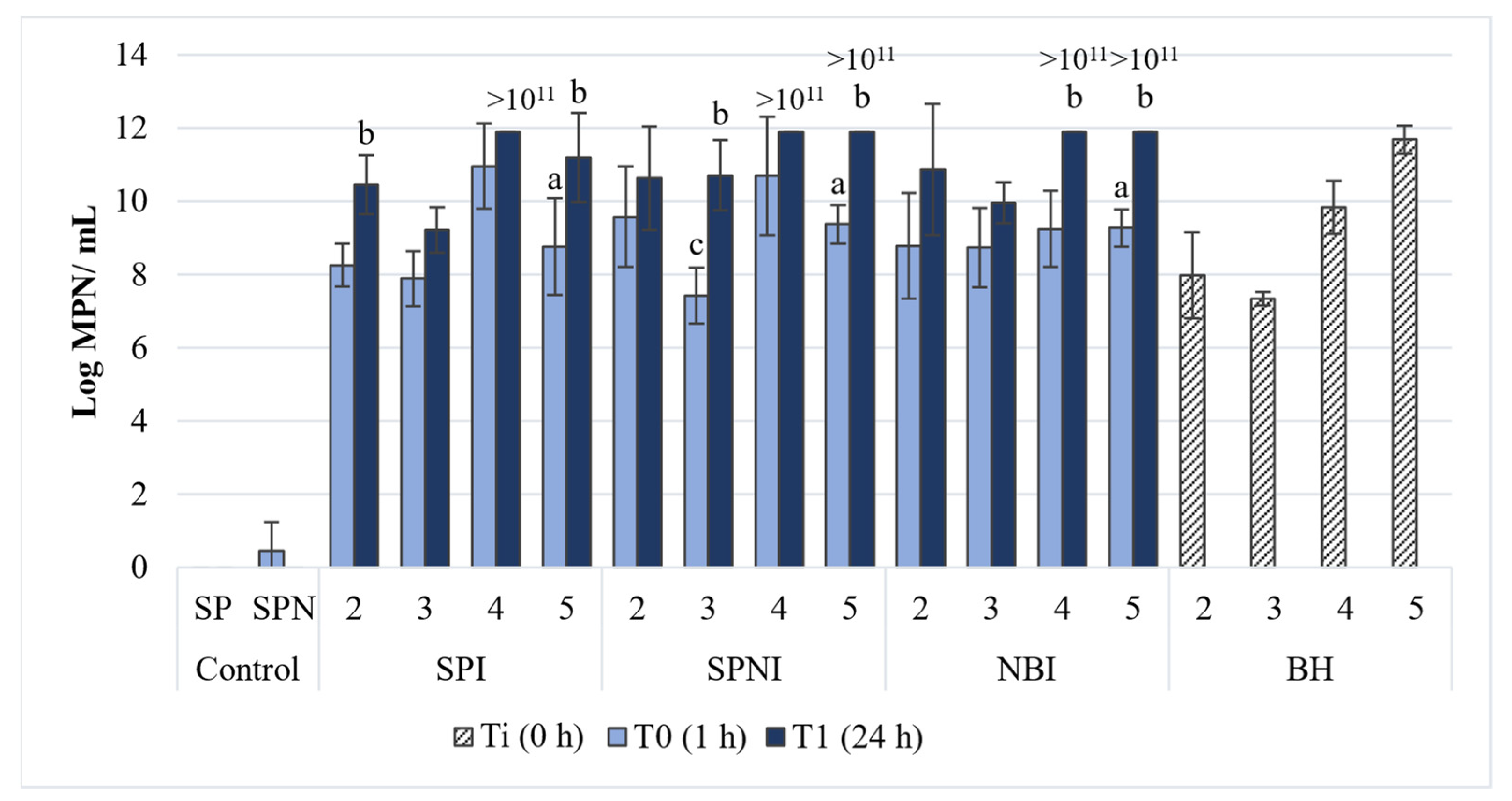{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Optimization of Hydrocarbon-Degrading Bacteria for Lyophilization
2.2. Evaluation of the Viability and Hydrocarbon-Degradation Potential of the Lyophilized Bacteria
2.2.1. Experiments with the Lyophilized Strains CPN2, CPN3, 1.7 L
2.2.2. Experiments with Different Concentrations of the Lyophilized Consortium
2.3. Microcosms Bioremediation Experiment in Natural Seawater
2.4. Abundance of Hydrocarbon-Degrading Bacteria by the MPN Method
2.5. Analyses of the Total Petroleum Hydrocarbons (TPHs)
2.6. Isolation of Potential Hydrocarbon-Degrading Bacteria
2.7. Identification of Bacterial Strains
2.8. Data Analysis
3. Results
3.1. Abundance of Hydrocarbon-Degraders in the Experiments with Lyophilized Strains
3.1.1. Experiments with the Strains CPN2, CPN3, 1.7 L and Their Consortium
3.1.2. Experiments with Different Concentrations of the Consortium of the Three Lyophilized Strains
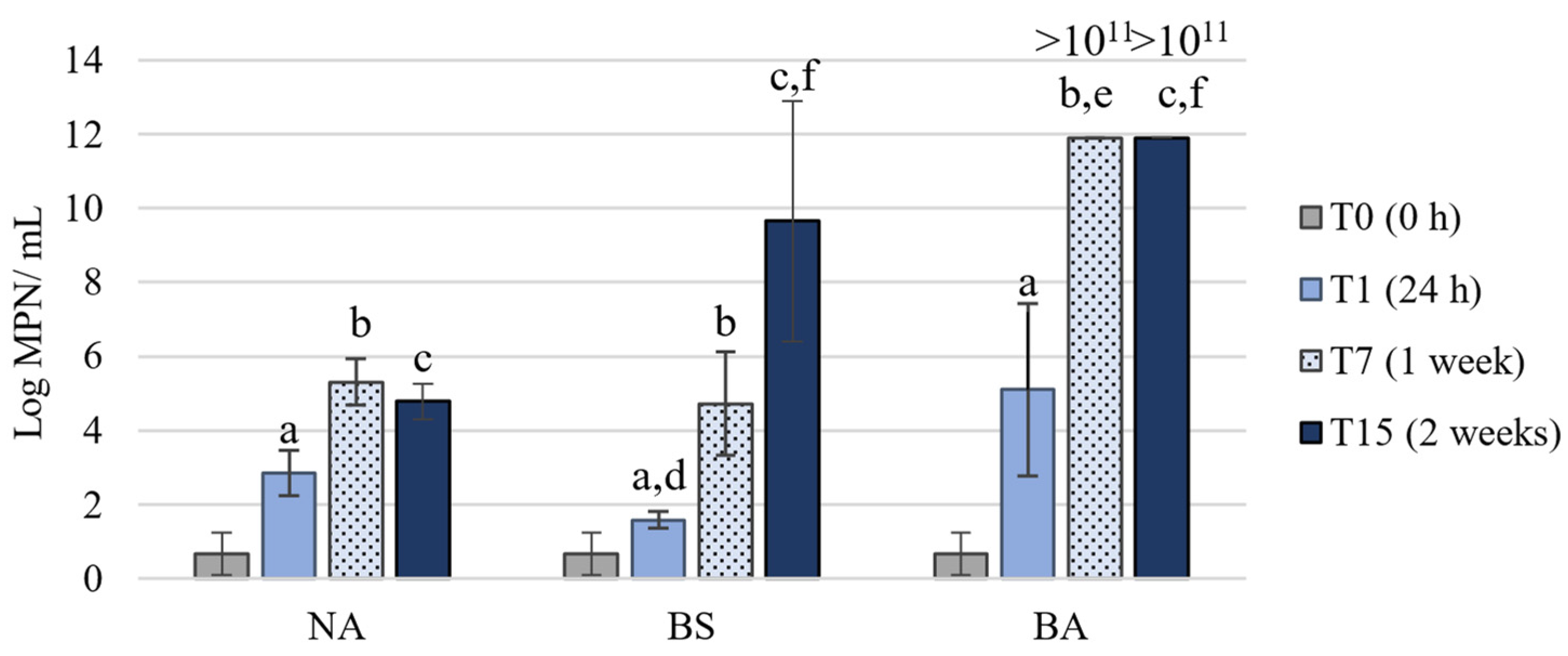
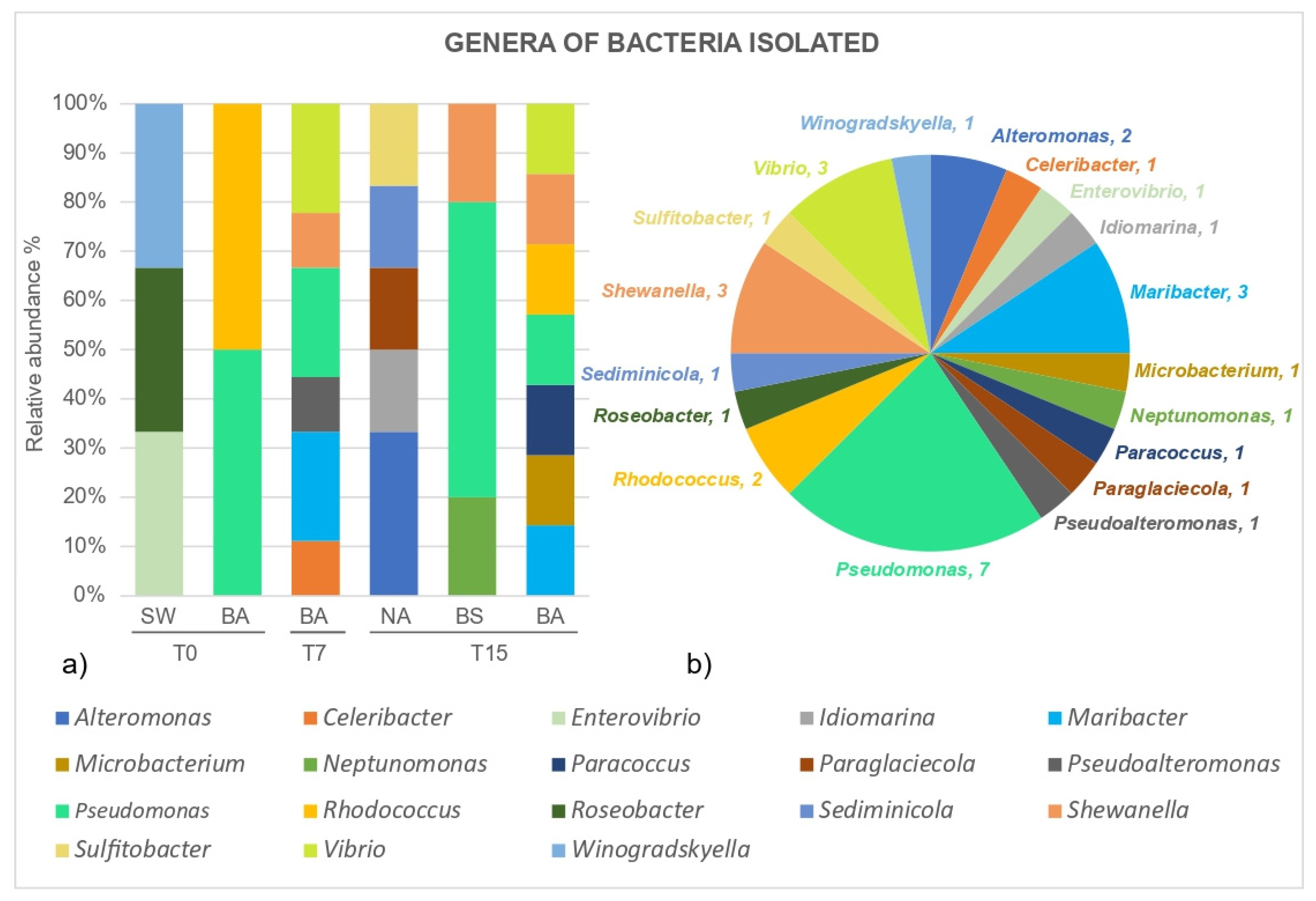
3.2. Microcosm Bioremediation Experiment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crisafi, F.; Genovese, M.; Smedile, F.; Russo, D.; Catalfamo, M.; Yakimov, M.; Giuliano, L.; Denaro, R. Bioremediation technologies for polluted seawater sampled after an oil-spill in Taranto Gulf (Italy): A comparison of biostimulation, bioaugmentation and use of a washing agent in microcosm studies. Mar. Pollut. Bull. 2016, 106, 119–126. [Google Scholar] [CrossRef]
- Barron, M.G. Ecological Impacts of the Deepwater Horizon Oil Spill: Implications for Immunotoxicity. Toxicol. Pathol. 2012, 40, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Atlas, R.M.; Hazen, T.C. Oil biodegradation and bioremediation: A tale of the two worst spills in U.S. history. Environ. Sci Technol. 2011, 45, 6709–6715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, C.M.; Reis, I.; Couto, M.N.; Bordalo, A.A.; Mucha, A.P. Potential of the microbial community present in an unimpacted beach sediment to remediate petroleum hydrocarbons. Env. Sci. Pollut. Res. Int. 2013, 20, 3176–3184. [Google Scholar] [CrossRef]
- Li, X.; Zhao, L.; Adam, M. Biodegradation of marine crude oil pollution using a salt-tolerant bacterial consortium isolated from Bohai Bay, China. Mar. Pollut. Bull. 2016, 105, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Priya, A.; Manab Sarma, P.; Lal, B. Isolation and characterization of Candida vishwanathiistrain TERI MS1 for degradation of petroleum hydrocarbons in marine environment. Desalination Water Treat. 2016, 57, 22099–22106. [Google Scholar] [CrossRef]
- Nikolopoulou, M.; Pasadakis, N.; Kalogerakis, N. Evaluation of autochthonous bioaugmentation and biostimulation during microcosm-simulated oil spills. Mar. Pollut. Bull. 2013, 72, 165–173. [Google Scholar] [CrossRef]
- Perdigão, R.; Almeida, C.M.R.; Santos, F.; Carvalho, M.F.; Mucha, A.P. Optimization of an Autochthonous Bacterial Consortium Obtained from Beach Sediments for Bioremediation of Petroleum Hydrocarbons. Water 2021, 13, 66. [Google Scholar] [CrossRef]
- Maamar, A.; Lucchesi, M.-E.; Debaets, S.; Nguyen van Long, N.; Quemener, M.; Coton, E.; Bouderbala, M.; Burgaud, G.; Matallah-Boutiba, A. Highlighting the Crude Oil Bioremediation Potential of Marine Fungi Isolated from the Port of Oran (Algeria). Diversity 2020, 12, 196. [Google Scholar] [CrossRef]
- Zinjarde, S.; Pant, A. Hydrocarbon degraders from tropical marine environments. Mar. Pollut. Bull. 2002, 44, 118–121. [Google Scholar] [CrossRef]
- Leahy, J.G.; Colwell, R.R. Microbial degradation of hydrocarbons in the environment. Microbiol. Rev. 1990, 54, 305–315. [Google Scholar] [CrossRef]
- Djahnit, N.; Chernai, S.; Catania, V.; Hamdi, B.; China, B.; Cappello, S.; Quatrini, P. Isolation, characterization and determination of biotechnological potential of oil-degrading bacteria from Algerian centre coast. J. Appl. Microbiol. 2019, 126, 780–795. [Google Scholar] [CrossRef]
- Harwati, T.U.; Kasai, Y.; Kodama, Y.; Susilaningsih, D.; Watanabe, K. Characterization of diverse hydrocarbon-degrading bacteria isolated from Indonesian seawater. Microbes Environ. 2007, 22, 412–415. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Zhong, R.; Shan, D.; Shao, Z. Indigenous oil-degrading bacteria in crude oil-contaminated seawater of the Yellow sea, China. Appl. Microbiol. Biotechnol. 2014, 98, 7253–7269. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, R.; Nagai, M.; Morikawa, M.; Okuyama, H. Autochthonous bioaugmentation and its possible application to oil spills. World J. Microbiol. Biotechnol. 2009, 25, 1519–1528. [Google Scholar] [CrossRef] [Green Version]
- Varjani, S.J. Microbial degradation of petroleum hydrocarbons. Bioresour. Technol. 2017, 223, 277–286. [Google Scholar] [CrossRef]
- Fuentes, S.; Mendez, V.; Aguila, P.; Seeger, M. Bioremediation of petroleum hydrocarbons: Catabolic genes, microbial communities, and applications. Appl. Microbiol. Biotechnol. 2014, 98, 4781–4794. [Google Scholar] [CrossRef] [PubMed]
- Souza, E.C.; Vessoni-Penna, T.C.; de Souza Oliveira, R.P. Biosurfactant-enhanced hydrocarbon bioremediation: An overview. Int. Biodeterior. Biodegrad. 2014, 89, 88–94. [Google Scholar] [CrossRef]
- Dehvari, M.; Ghafari, S.; Haghighifard, N.J.; Jorfi, S. Petroleum Contaminated Seawater Detoxification in Microcosm by Halotolerant Consortium Isolated from Persian Gulf. Curr. Microbiol. 2021, 78, 95–106. [Google Scholar] [CrossRef]
- Pereira, E.; Napp, A.P.; Allebrandt, S.; Barbosa, R.; Reuwsaat, J.; Lopes, W.; Kmetzsch, L.; Staats, C.C.; Schrank, A.; Dallegrave, A. Biodegradation of aliphatic and polycyclic aromatic hydrocarbons in seawater by autochthonous microorganisms. Int. Biodeterior. Biodegrad. 2019, 145, 104789. [Google Scholar] [CrossRef]
- Fodelianakis, S.; Antoniou, E.; Mapelli, F.; Magagnini, M.; Nikolopoulou, M.; Marasco, R.; Barbato, M.; Tsiola, A.; Tsikopoulou, I.; Giaccaglia, L.; et al. Allochthonous bioaugmentation in ex situ treatment of crude oil-polluted sediments in the presence of an effective degrading indigenous microbiome. J. Hazard. Mater. 2015, 287, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Tao, K.; Liu, X.; Chen, X.; Hu, X.; Cao, L.; Yuan, X. Biodegradation of crude oil by a defined co-culture of indigenous bacterial consortium and exogenous Bacillus subtilis. Bioresour. Technol. 2017, 224, 327–332. [Google Scholar] [CrossRef]
- Das, N.; Chandran, P. Microbial degradation of petroleum hydrocarbon contaminants: An overview. Biotechnol. Res. Int. 2011, 2011, 941810. [Google Scholar] [CrossRef] [Green Version]
- Ivshina, I.B.; Kuyukina, M.S.; Krivoruchko, A.V.; Elkin, A.A.; Makarov, S.O.; Cunningham, C.J.; Peshkur, T.A.; Atlas, R.M.; Philp, J.C. Oil spill problems and sustainable response strategies through new technologies. Environ. Sci. Process. Impacts 2015, 17, 1201–1219. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Liu, W.; Tian, S.; Wang, W.; Qi, Q.; Jiang, P.; Gao, X.; Li, F.; Li, H.; Yu, H. Petroleum Hydrocarbon-Degrading Bacteria for the Remediation of Oil Pollution Under Aerobic Conditions: A Perspective Analysis. Front. Microbiol. 2018, 9, 2885. [Google Scholar] [CrossRef]
- Partovinia, A.; Rasekh, B. Review of the immobilized microbial cell systems for bioremediation of petroleum hydrocarbons polluted environments. Crit. Rev. Environ. Sci. Technol. 2018, 48, 1–38. [Google Scholar] [CrossRef]
- Dai, X.; Lv, J.; Yan, G.; Chen, C.; Guo, S.; Fu, P. Bioremediation of intertidal zones polluted by heavy oil spilling using immobilized laccase-bacteria consortium. Bioresour. Technol. 2020, 309, 123305. [Google Scholar] [CrossRef]
- Wang, X.; Liu, Y.; Song, C.; Yuan, X.; Zhang, Q.; Miao, Y. Application Analysis of Immobilized Bioremediation Preparation in Oil Spill Contaminated Shore. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2020. [Google Scholar]
- Alessandrello, M.J.; Tomás, M.S.J.; Raimondo, E.E.; Vullo, D.L.; Ferrero, M.A. Petroleum oil removal by immobilized bacterial cells on polyurethane foam under different temperature conditions. Mar. Pollut. Bull. 2017, 122, 156–160. [Google Scholar] [CrossRef]
- Chen, C.-H.; Whang, L.-M.; Pan, C.-L.; Yang, C.-L.; Liu, P.-W.G. Immobilization of diesel-degrading consortia for bioremediation of diesel-contaminated groundwater and seawater. Int. Biodeterior. Biodegrad. 2017, 124, 62–72. [Google Scholar] [CrossRef]
- Hou, D.; Shen, X.; Luo, Q.; He, Y.; Wang, Q.; Liu, Q. Enhancement of the diesel oil degradation ability of a marine bacterial strain by immobilization on a novel compound carrier material. Mar. Pollut. Bull. 2013, 67, 146–151. [Google Scholar] [CrossRef]
- Tsutsumi, H.; Kono, M.; Takai, K.; Manabe, T.; Haraguchi, M.; Yamamoto, I.; Oppenheimer, C. Bioremediation on the shore after an oil spill from the Nakhodka in the Sea of Japan. III. Field tests of a bioremediation agent with microbiological cultures for the treatment of an oil spill. Mar. Pollut. Bull. 2000, 40, 320–324. [Google Scholar] [CrossRef]
- Villela, H.D.M.; Peixoto, R.S.; Soriano, A.U.; Carmo, F.L. Microbial bioremediation of oil contaminated seawater: A survey of patent deposits and the characterization of the top genera applied. Sci. Total Environ. 2019, 666, 743–758. [Google Scholar] [CrossRef] [PubMed]
- Elisabeth Helmke, B.D.; Birte Gerdes, B.D.J.; Juergens, B.D.K.; Reuter, L.D. Bioremediation Method for Accelerated Biological Decomposition of Petroleum Hydrocarbons in Sea-Iced Covered Polar Regions, and Bacteria and Enzyme Mixtures as Agents for Carrying out Said Method. U.S. Patent 8,444,962, 21 May 2013. [Google Scholar]
- Adams, G. The Principles of Freeze-Drying. In Cryopreservation and Freeze-Drying Protocols; Day, J.G., Stacey, G.N., Eds.; Humana Press: Totowa, NJ, USA, 2007; pp. 15–38. [Google Scholar]
- Kupletskaya, M.; Netrusov, A. Viability of lyophilized microorganisms after 50-year storage. Microbiology 2011, 80, 850–853. [Google Scholar] [CrossRef]
- Gouveia, V. Bioremediation of Oil Along the Nw Portuguese Coast-the Role of Autochthonous Microorganisms. Master’s Thesis, School of Medicine and Biomedical Sciences, University of Porto, Porto, Portugal, 2015. [Google Scholar]
- Wrenn, B.A.; Venosa, A.D. Selective enumeration of aromatic and aliphatic hydrocarbon degrading bacteria by a most-probable-number procedure. Can. J. Microbiol. 1996, 42, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Head, I.M.; Jones, D.M.; Röling, W.F. Marine microorganisms make a meal of oil. Nat. Rev. Microbiol. 2006, 4, 173–182. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Dutta, A.; Pal, S.; Gupta, A.; Sarkar, J.; Chatterjee, A.; Saha, A.; Sarkar, P.; Sar, P.; Kazy, S.K. Biostimulation and bioaugmentation of native microbial community accelerated bioremediation of oil refinery sludge. Bioresour. Technol. 2018, 253, 22–32. [Google Scholar] [CrossRef]
- Tyagi, M.; da Fonseca, M.M.R.; de Carvalho, C. Bioaugmentation and biostimulation strategies to improve the effectiveness of bioremediation processes. Biodegradation 2011, 22, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Reis, I.; Almeida, C.M.; Magalhaes, C.M.; Cochofel, J.; Guedes, P.; Basto, M.C.; Bordalo, A.A.; Mucha, A.P. Bioremediation potential of microorganisms from a sandy beach affected by a major oil spill. Environ. Sci. Pollut. Res. 2014, 21, 3634–3645. [Google Scholar] [CrossRef] [PubMed]
- Santisi, S.; Catalfamo, M.; Bonsignore, M.; Gentile, G.; Di Salvo, E.; Genovese, M.; Mahjoubi, M.; Cherif, A.; Mancini, G.; Hassanshahian, M. Biodegradation ability of two selected microbial autochthonous consortia from a chronically polluted marine coastal area (Priolo Gargallo, Italy). J. Appl. Microbiol. 2019, 127, 618–629. [Google Scholar] [CrossRef]
- Hassanshahian, M.; Emtiazi, G.; Caruso, G.; Cappello, S. Bioremediation (bioaugmentation/biostimulation) trials of oil polluted seawater: A mesocosm simulation study. Mar. Environ. Res. 2014, 95, 28–38. [Google Scholar] [CrossRef]
- Alexandrino, D.A.M.; Ribeiro, I.; Pinto, L.M.; Cambra, R.; Oliveira, R.S.; Pereira, F.; Carvalho, M.F. Biodegradation of mono-, di- and trifluoroacetate by microbial cultures with different origins. New Biotechnol. 2018, 43, 23–29. [Google Scholar] [CrossRef]
- Santos, F.; Mucha, A.P.; Alexandrino, D.A.M.; Almeida, C.M.R.; Carvalho, M.F. Biodegradation of enrofloxacin by microbial consortia obtained from rhizosediments of two estuarine plants. J. Environ. Manag. 2019, 231, 1145–1153. [Google Scholar] [CrossRef] [PubMed]
- Duarte, P.; Almeida, C.M.R.; Fernandes, J.P.; Morais, D.; Lino, M.; Gomes, C.R.; Carvalho, M.F.; Mucha, A.P. Bioremediation of bezafibrate and paroxetine by microorganisms from estuarine sediment and activated sludge of an associated wastewater treatment plant. Sci. Total Environ. 2019, 655, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Poblete-Castro, I.; Wittmann, C.; Nikel, P.I. Biochemistry, genetics and biotechnology of glycerol utilization in Pseudomonas species. Microb. Biotechnol. 2020, 13, 32–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sriwongchai, S.; Pokethitiyook, P.; Pugkaew, W.; Kruatrachue, M.; Lee, H. Optimization of lipid production in the oleaginous bacterium Rhodococcus erythropolis growing on glycerol as the sole carbon source. Afr. J. Biotechnol. 2012, 11, 14440–14447. [Google Scholar] [CrossRef]
- Li, H.; Li, Y.; Bao, M.; Li, S. Solid inoculants as a practice for bioaugmentation to enhance bioremediation of hydrocarbon contaminated areas. Chemosphere 2020, 263, 128175. [Google Scholar] [CrossRef]
- Yan, F.; Wang, Y. Remediation of marine pollution by microorganisms in the comprehensive management of coastal zones. J. Coast. Res. 2018, 239–244. [Google Scholar] [CrossRef]
- Peng, F.; Liu, Z.; Wang, L.; Shao, Z. An oil-degrading bacterium: Rhodococcus erythropolis strain 3C-9 and its biosurfactants. J. Appl. Microbiol. 2007, 102, 1603–1611. [Google Scholar] [CrossRef]
- Luong, T.M.; Ponamoreva, O.N.; Nechaeva, I.A.; Petrikov, K.V.; Delegan, Y.A.; Surin, A.K.; Linklater, D.; Filonov, A.E. Characterization of biosurfactants produced by the oil-degrading bacterium Rhodococcus erythropolis S67 at low temperature. World J. Microbiol. Biotechnol. 2018, 34, 20. [Google Scholar] [CrossRef]
- Mohanram, R.; Jagtap, C.; Kumar, P. Isolation, screening, and characterization of surface-active agent-producing, oil-degrading marine bacteria of Mumbai Harbor. Mar. Pollut. Bull. 2016, 105, 131–138. [Google Scholar] [CrossRef]
- Shi, K.; Zhang, Q.; Xue, J.; Chen, X.; Chen, Y.; Qiao, Y.; Yang, Y.; Sun, J. Study on the degradation performance and bacterial community of bioaugmentation in petroleum-pollution seawater. J. Environ. Chem. Eng. 2020, 8, 103900. [Google Scholar] [CrossRef]
- Imron, M.F.; Titah, H.S. Optimization of diesel biodegradation by Vibrio alginolyticus using Box-Behnken design. Environ. Eng. Res. 2018, 23, 374–382. [Google Scholar] [CrossRef]
- Feknous, N.; Branes, Z.; Batisson, I.; Amblard, C. Growth of indigenous bacteria Vibrio alginolyticus and Dietzia sp. isolated from the east coast of Algeria in the presence of monoaromatic hydrocarbons. Environ. Prot. Eng. 2019, 45, 127–137. [Google Scholar] [CrossRef]
- Hedlund, B.P.; Staley, J.T. Isolation and characterization of Pseudoalteromonas strains with divergent polycyclic aromatic hydrocarbon catabolic properties. Environ. Microbiol. 2006, 8, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.M.; Kim, J.M.; Lee, H.J.; Madsen, E.L.; Jeon, C.O. Alteromonas as a key agent of polycyclic aromatic hydrocarbon biodegradation in crude oil-contaminated coastal sediment. Environ. Sci. Technol. 2012, 46, 7731–7740. [Google Scholar] [CrossRef]
- Diéguez, A.L.; Pichon, P.; Balboa, S.; Magnesen, T.; Romalde, J.L. Complete characterization of new isolates of Neptunomonas phycophila leads to emend its description and opens possibilities of biotechnological applications. MicrobiologyOpen 2017, 6, e00519. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Yang, Q.Q.; Ren, Y.; Zhang, W.W.; Zheng, G.; Sun, C.; Pan, J.; Zhu, X.F.; Zhang, X.Q.; Wu, M. Maribacter thermophilus sp. nov., isolated from an algal bloom in an intertidal zone, and emended description of the genus Maribacter. Int. J. Syst. Evol. Microbiol. 2015, 65 Pt 1, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Venosa, A.D.; Zhu, X. Biodegradation of Crude Oil Contaminating Marine Shorelines and Freshwater Wetlands. Spill Sci. Technol. Bull. 2003, 8, 163–178. [Google Scholar] [CrossRef]
- Junusmin, K.I.; Manurung, B.S.; Darmayati, Y. Bioremediation of Oil-Contaminated Sediment by Hydrocarbonoclastic Bacterial Consortium Immobilized in Different Types of Carrier. In Proceedings of the 5th International Symposium on Applied Chemistry (ISAC), Tangerang, Indonesia, 23–24 October 2019; Available online: https://aip.scitation.org/doi/abs/10.1063/1.5134620 (accessed on 10 September 2021).
- Luo, Q.; Hou, D.; Jiang, D.; Chen, W. Bioremediation of marine oil spills by immobilized oil-degrading bacteria and nutrition emulsion. Biodegradation 2021, 32, 165–177. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perdigão, R.; Almeida, C.M.R.; Magalhães, C.; Ramos, S.; Carolas, A.L.; Ferreira, B.S.; Carvalho, M.F.; Mucha, A.P. Bioremediation of Petroleum Hydrocarbons in Seawater: Prospects of Using Lyophilized Native Hydrocarbon-Degrading Bacteria. Microorganisms 2021, 9, 2285. https://doi.org/10.3390/microorganisms9112285
Perdigão R, Almeida CMR, Magalhães C, Ramos S, Carolas AL, Ferreira BS, Carvalho MF, Mucha AP. Bioremediation of Petroleum Hydrocarbons in Seawater: Prospects of Using Lyophilized Native Hydrocarbon-Degrading Bacteria. Microorganisms. 2021; 9(11):2285. https://doi.org/10.3390/microorganisms9112285
Chicago/Turabian StylePerdigão, Rafaela, C. Marisa R. Almeida, Catarina Magalhães, Sandra Ramos, Ana L. Carolas, Bruno S. Ferreira, Maria F. Carvalho, and Ana P. Mucha. 2021. "Bioremediation of Petroleum Hydrocarbons in Seawater: Prospects of Using Lyophilized Native Hydrocarbon-Degrading Bacteria" Microorganisms 9, no. 11: 2285. https://doi.org/10.3390/microorganisms9112285
APA StylePerdigão, R., Almeida, C. M. R., Magalhães, C., Ramos, S., Carolas, A. L., Ferreira, B. S., Carvalho, M. F., & Mucha, A. P. (2021). Bioremediation of Petroleum Hydrocarbons in Seawater: Prospects of Using Lyophilized Native Hydrocarbon-Degrading Bacteria. Microorganisms, 9(11), 2285. https://doi.org/10.3390/microorganisms9112285