
Biochemical Characterization of Recombinant Isocitrate Dehydrogenase and Its Putative Role in the Physiology of an Acidophilic Micrarchaeon


Abstract
:1. Introduction
2. Materials and Methods
2.1. Database Research and Bioinformatic Sequence and Structure Analyses
2.2. Cloning and Recombinant Expression of icd26x His
2.3. Isolation and Affinity Purification of MhIDH6x His
2.4. Protein Quantification, SDS-PAGE & Western Blot
2.5. Spectrophotometric IDH-Activity Assays and Determination of Kinetic Properties
3. Results & Discussion
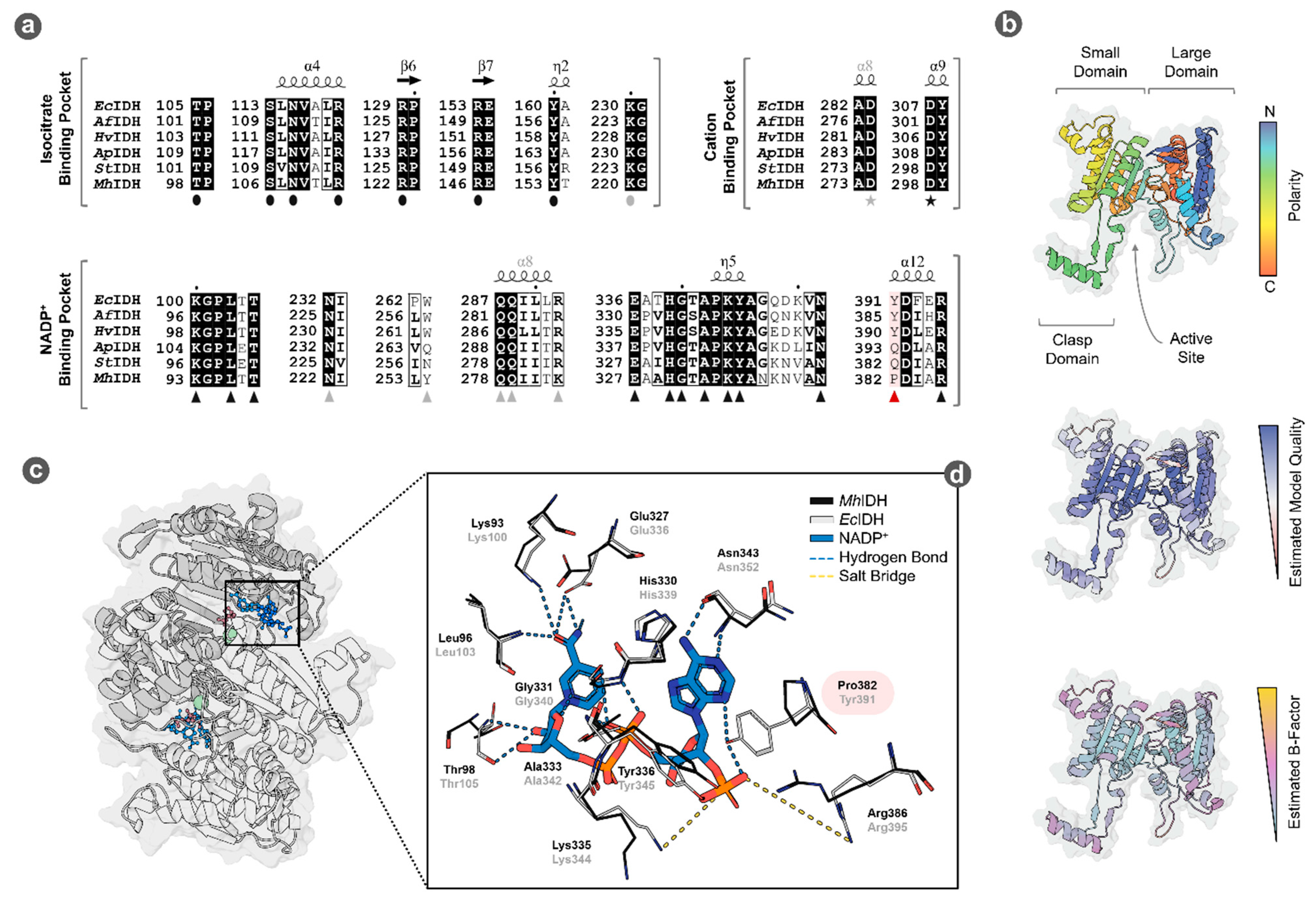
3.1. MhIDH Shows Conserved Characteristics of Prokaryotic, NADP-Dependent IDHs
3.2. MhIDH6x His Can Be Produced in E. coli
3.3. Biochemical Properties of MhIDH6x His
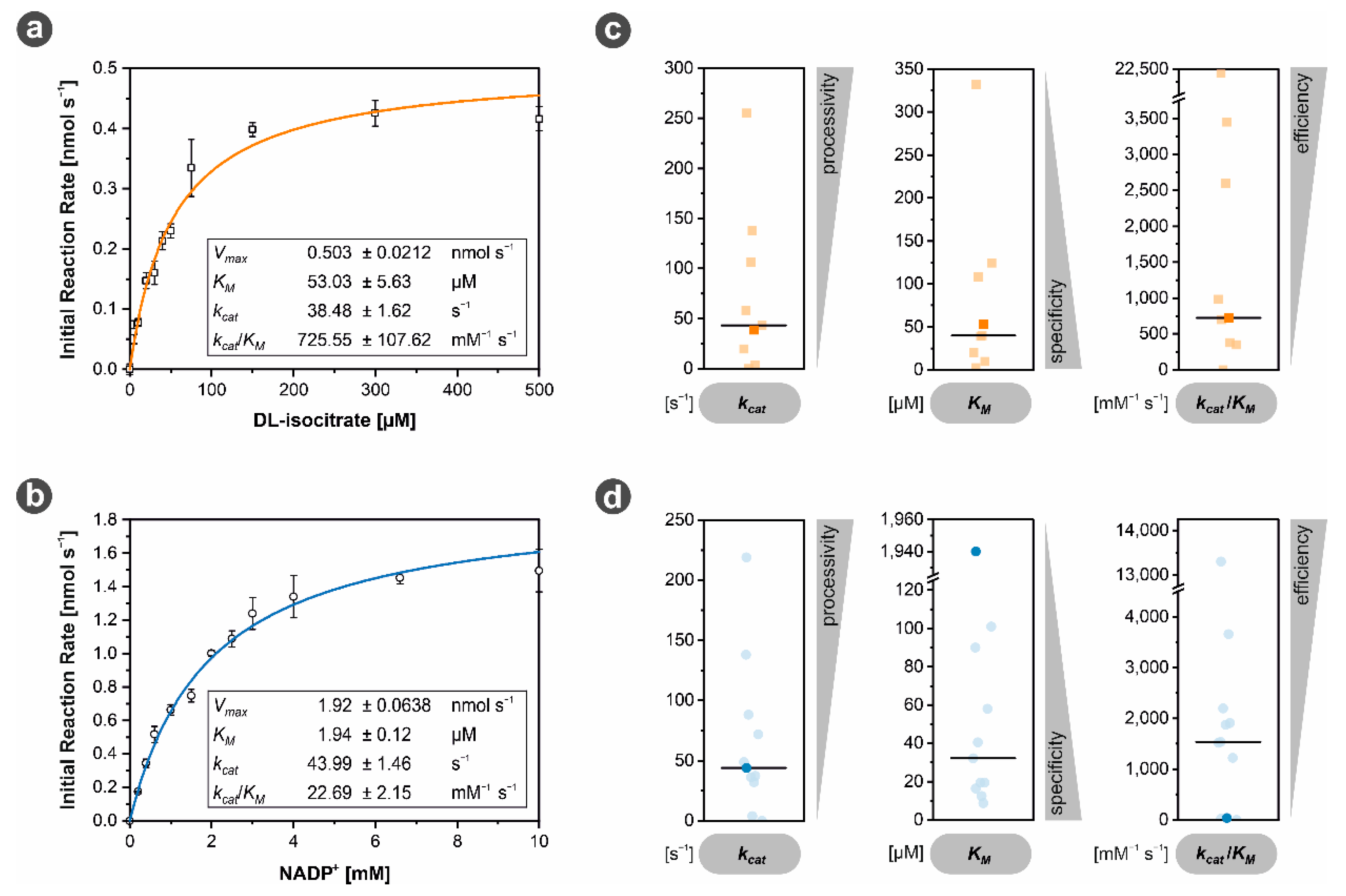
3.3.1. MhIDH6x His Activity Is Dependent on NADP+ and Divalent Cations
3.3.2. MhIDH6x His Shows Highest Activity at Slightly Alkaline pH
3.3.3. MhIDH6x His Is Characterized by Low NADP+ Affinity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NCBI Accession | Description | e-Value | Identity [%] |
|---|---|---|---|
| O29610.1 | Isocitrate dehydrogenase [NADP]; [Archaeoglobus fulgidus DSM 4304] | 3.54 × 10−150 | 55.47 |
| P96318.2 | Isocitrate dehydrogenase [NADP]; [Caldococcus noboribetus] | 9.487 × 10−147 | 54.61 |
| D4GU92.1 | Isocitrate dehydrogenase [NADP]; [Haloferax volcanii DS2] | 1.2741 × 10−134 | 48.05 |
| P08200.1 | Isocitrate dehydrogenase [NADP]; [Escherichia coli K-12] | 9.9176 × 10−128 | 49.75 |
| P39126.1 | Isocitrate dehydrogenase [NADP]; [Bacillus subtilis subsp. subtilis str. 168] | 5.42 × 10−127 | 49.40 |
| Q9ZH99.1 | Isocitrate dehydrogenase [NADP]; [Coxiella burnetii RSA 493] | 2.2534 × 10−126 | 49.26 |
| P65099.1 | Isocitrate dehydrogenase [NADP]; [Staphylococcus aureus subsp. aureus Mu50] | 5.4508 × 10−125 | 49.14 |
| Q6G8N2.1 | Isocitrate dehydrogenase [NADP]; [Staphylococcus aureus subsp. aureus MSSA476] | 6.9239 × 10−125 | 49.14 |
| Q5HNL1.1 | Isocitrate dehydrogenase [NADP]; [Staphylococcus epidermidis RP62A] | 9.4905 × 10−125 | 49.14 |
| Q6GG12.1 | Isocitrate dehydrogenase [NADP]; [Staphylococcus aureus subsp. aureus MRSA252] | 1.2055 × 10−124 | 49.63 |
| P56063.1 | Isocitrate dehydrogenase [NADP]; [Helicobacter pylori 26695] | 2.7939 × 10−122 | 49.27 |
| Q9ZN36.1 | Isocitrate dehydrogenase [NADP]; [Helicobacter pylori J99] | 6.5213 × 10−122 | 48.66 |
| Q02NB5.1 | Isocitrate dehydrogenase [NADP]; [Pseudomonas aeruginosa UCBPP-PA14] | 7.6852 × 10−122 | 49.23 |
| Q59940.2 | Isocitrate dehydrogenase [NADP]; [Streptococcus mutans UA159] | 3.6802 × 10−120 | 46.02 |
| P41560.2 | Isocitrate dehydrogenase [NADP] 1; [Colwellia maris] | 1.0682 × 10−119 | 48.65 |
| Q59985.1 | Isocitrate dehydrogenase [NADP]; [Streptococcus salivarius] | 9.7535 × 10−116 | 46.27 |
| P50214.1 | Isocitrate dehydrogenase [NADP]; [Nostoc sp. PCC 7120 = FACHB-418] | 1.995 × 10−113 | 42.58 |
| P80046.2 | Isocitrate dehydrogenase [NADP]; [Synechocystis sp. PCC 6803 substr. Kazusa] | 3.0758 × 10−109 | 42.45 |
| O67480.1 | Isocitrate dehydrogenase [NADP]; [Aquifex aeolicus VF5] | 4.518 × 10−109 | 46.62 |
| Q4UKR1.2 | Isocitrate dehydrogenase [NADP]; [Rickettsia felis URRWXCal2] | 3.03035 × 10−59 | 35.88 |
| Q1RJU4.1 | Isocitrate dehydrogenase [NADP]; [Rickettsia bellii RML369-C] | 6.19516 × 10−58 | 34.82 |
| Q92IR7.1 | Isocitrate dehydrogenase [NADP]; [Rickettsia conorii str. Malish 7] | 4.10157 × 10−57 | 35.09 |
| Q9ZDR0.1 | Isocitrate dehydrogenase [NADP]; [Rickettsia prowazekii str. Madrid E] | 5.79905 × 10−57 | 35.45 |
| … | … | … | … |
| O27441.1 | 3-isopropylmalate dehydrogenase; [Methanothermobacter thermautotrophicus str. Delta H] | 4.50989 × 10−51 | 30.97 |
| … | … | … | … |
| O29627.1 | 3-isopropylmalate dehydrogenase; [Archaeoglobus fulgidus DSM 4304] | 8.66687 × 10−47 | 32.41 |
| P50455.3 | 3-isopropylmalate dehydrogenase; [Sulfurisphaera tokodaii str. 7] | 1.07086 × 10−46 | 30.81 |
| Organism | pH Optimum | DL-Isocitric Acid | NADP+ | Refs | ||||
|---|---|---|---|---|---|---|---|---|
| kcat [s−1] | KM [mM] | kcat/KM [mM−1 s−1] | kcat [s−1] | KM [mM] | kcat/KM [mM−1 s−1] | |||
| Archaeoglobus fulgidus | 8.6 | 255 | 0.332 | 700 | 219 | 0.0165 | 13 300 | [40,53] |
| Bifidobacterium longum subsp. infantis | 8.0 | NA | NA | NA | 36.4 | 0.01945 | 1 871 * | [49] |
| Escherichia coli | 8.0 | 106.4 | 0.0405 | 2 600 | 88.1 | 0.0392 | 2 200 | [53] |
| Haloferax volcanii | 8.0 | 0.0023 | 0.108 | 0.021 * | 0.003 | 0.101 | 0.030 * | [18,54,55] |
| Microcystis aeruginosa | 7.5 | 43.21 | 0.1243 | 347.63 * | 48.88 | 0.0322 | 1 518 * | [56] |
| Mycobacterium tuberculosis 1 | 7.5 | 3.8 | 0.01 | 380 * | 4 | 0.125 | 32 * | [57] |
| Mycobacterium tuberculosis 2 | 7.5 | 19.6 | 0.02 | 980 * | 37.4 | 0.0196 | 1 908 * | [57] |
| Plasmodium falciparum | 8.0 | 138 | 0.04 | 3 450 * | 138 | 0.09 | 1 533 * | [58] |
| Saccharolobus solfataricus | 8.0 | NA | NA | NA | NA | NA | NA | [18] |
| Sus scrofa | 7.4 | 58.3 | 0.0026 | 22 423 * | 32.2 | 0.0088 | 3 659 * | [59,60] |
| Thermoplasma acidophilum | 7.5 | NA | NA | NA | NA | NA | NA | [61] |
| Yarrowia lipolytica | 8.5 | NA | NA | NA | 72 | 0.058 | 1 220 | [62] |
| “Ca. Micrarchaeum harzensis A_DKE” | 8.0 | 38.48 ± 1.62 | 0.0530 ± 0.0056 | 725 ± 107.62 | 43.99 ± 1.46 | 1.94 ± 0.12 | 22.69 ± 2.15 | This study |
References
- Kristjánsson, J.K.; Hreggvidsson, G.O. Ecology and habitats of extremophiles. World J. Microbiol. Biotechnol. 1995, 11, 17–25. [Google Scholar] [CrossRef]
- Pikuta, E.V.; Hoover, R.B.; Tang, J. Microbial extremophiles at the limits of life. Crit. Rev. Microbiol. 2007, 33, 183–209. [Google Scholar] [CrossRef] [PubMed]
- Horikoshi, K.; Antranikian, G.; Bull, A.T.; Robb, F.T.; Stetter, K.O. Extremophiles Handbook Volume 1. J. Chem. Inf. Model. 1981, 53, 1689–1699. [Google Scholar] [CrossRef]
- Charlesworth, J.; Burns, B.P. Extremophilic adaptations and biotechnological applications in diverse environments. AIMS Microbiol. 2016, 2, 251–261. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Dopson, M. Life in acid: PH homeostasis in acidophiles. Trends Microbiol. 2007, 15, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Golyshina, O.V.; Golyshin, P.N.; Timmis, K.N.; Ferrer, M. The “pH optimum anomaly” of intracellular enzymes of Ferroplasma acidiphilum. Environ. Microbiol. 2006, 8, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Slonczewski, J.L.; Fujisawa, M.; Dopson, M.; Krulwich, T.A. Cytoplasmic pH Measurement and Homeostasis in Bacteria and Archaea. Adv. Microb. Physiol. 2009, 55, 1–79, 317. [Google Scholar]
- Matin, A. pH Homeostasis in Acidophiles. In Novartis Foundation Symposium 221—Bacterial Responses to pH; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2007; pp. 152–166. ISBN 9780470515631. [Google Scholar]
- Kadnikov, V.V.; Savvichev, A.S.; Mardanov, A.V.; Beletsky, A.V.; Chupakov, A.V.; Kokryatskaya, N.M.; Pimenov, N.V.; Ravin, N.V. Metabolic Diversity and Evolutionary History of the Archaeal Phylum “ Candidatus Micrarchaeota” Uncovered from a Freshwater Lake Metagenome. Appl. Environ. Microbiol. 2020, 86, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.X.; Méndez-García, C.; Dombrowski, N.; Servín-Garcidueñas, L.E.; Eloe-Fadrosh, E.A.; Fang, B.Z.; Luo, Z.H.; Tan, S.; Zhi, X.Y.; Hua, Z.S.; et al. Metabolic versatility of small archaea Micrarchaeota and Parvarchaeota. ISME J. 2018, 12, 756–775. [Google Scholar] [CrossRef]
- Baker, B.J.; Comolli, L.R.; Dick, G.J.; Hauser, L.J.; Hyatt, D.; Dill, B.D.; Land, M.L.; VerBerkmoes, N.C.; Hettich, R.L.; Banfield, J.F. Enigmatic, ultrasmall, uncultivated Archaea. Proc. Natl. Acad. Sci. USA 2010, 107, 8806–8811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golyshina, O.V.; Toshchakov, S.V.; Makarova, K.S.; Gavrilov, S.N.; Korzhenkov, A.A.; La Cono, V.; Arcadi, E.; Nechitaylo, T.Y.; Ferrer, M.; Kublanov, I.V.; et al. ‘ARMAN’ archaea depend on association with euryarchaeal host in culture and in situ. Nat. Commun. 2017, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Krause, S.; Gfrerer, S.; Reuse, C.; Dombrowski, N.; Villanueva, L.; Bunk, B.; Spröer, C.; Neu, T.R.; Kuhlicke, U.; Schmidt-Hohagen, K.; et al. Unraveling the critical growth factors for stable cultivation of (nano-sized) Micrarchaeota. bioRxiv 2021. [Google Scholar] [CrossRef]
- Krause, S.; Bremges, A.; Münch, P.C.; McHardy, A.C.; Gescher, J. Characterisation of a stable laboratory co-culture of acidophilic nanoorganisms. Sci. Rep. 2017, 7, 3289. [Google Scholar] [CrossRef] [Green Version]
- Golyshina, O.V.; Bargiela, R.; Toshchakov, S.V.; Chernyh, N.A.; Ramayah, S.; Korzhenkov, A.A.; Kublanov, I.V.; Golyshin, P.N. Diversity of “Ca. Micrarchaeota” in Two Distinct Types of Acidic Environments and Their Associations with Thermoplasmatales. Genes 2019, 10, 461. [Google Scholar] [CrossRef] [Green Version]
- Ziegler, S.; Dolch, K.; Geiger, K.; Krause, S.; Asskamp, M.; Eusterhues, K.; Kriews, M.; Wilhelms-Dick, D.; Goettlicher, J.; Majzlan, J.; et al. Oxygen-dependent niche formation of a pyrite-dependent acidophilic consortium built by archaea and bacteria. ISME J. 2013, 7, 1725–1737. [Google Scholar] [CrossRef] [PubMed]
- Gfrerer, S.; Winkler, D.; Novion Ducassou, J.; Couté, Y.; Rachel, R.; Gescher, J. Micrarchaeota are covered by a proteinaceous S-layer. bioRxiv 2021. [Google Scholar] [CrossRef]
- Camacho, M.L.; Brown, R.A.; Bonete, M.J.; Danson, J.M.; Hough, D.W. Isocitrate dehydrogenases from Haloferax volcanii and Sulfolobus solfataricus: Enzyme purification, characterisation and N-terminal sequence. FEMS Microbiol. Lett. 1995, 134, 85–90. [Google Scholar] [CrossRef]
- Agarwala, R.; Barrett, T.; Beck, J.; Benson, D.A.; Bollin, C.; Bolton, E.; Bourexis, D.; Brister, J.R.; Bryant, S.H.; Canese, K.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2018, 46, D8–D13. [Google Scholar] [CrossRef] [Green Version]
- Jeske, L.; Placzek, S.; Schomburg, I.; Chang, A.; Schomburg, D. BRENDA in 2019: A European ELIXIR core data resource. Nucleic Acids Res. 2019, 47, D542–D549. [Google Scholar] [CrossRef]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, 279–285. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Barton, G.J.; Higgins, D.G. Multiple Sequence Alignments. In Bioinformatics; Baxevanis, A.D., Bader, G.D., Wishart, D.S., Eds.; Wiley: Hoboken, NJ, USA, 2020; pp. 227–250. ISBN 9781119335580. [Google Scholar]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, 320–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, S.; Miller, S.P.; Carrondo, M.A.; Dean, A.M.; Matias, P.M. Induced fit and the catalytic mechanism of isocitrate dehydrogenase. Biochemistry 2012, 51, 7098–7115. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Wang, Y.; Zhang, Y. ResQ: An Approach to Unified Estimation of B-Factor and Residue-Specific Error in Protein Structure Prediction. J. Mol. Biol. 2016, 428, 693–701. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Roy, A.; Zhang, Y. BioLiP: A semi-manually curated database for biologically relevant ligand-protein interactions. Nucleic Acids Res. 2013, 41, 1096–1103. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Roy, A.; Zhang, Y. Protein-ligand binding site recognition using complementary binding-specific substructure comparison and sequence profile alignment. Bioinformatics 2013, 29, 2588–2595. [Google Scholar] [CrossRef]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully automated protein-ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Reeves, H.C.; Daumy, G.O.; Lin, C.C.; Houston, M. NADP+-Specific Isocitrate Dehydrogenase of Escherichia coli. Biochim. Biophys. Acta 1972, 258, 27–39. [Google Scholar] [CrossRef]
- Michaelis, L.; Menten, M.L. Die Kinetik der Invertinwirkung. Biochem. Z. 1913, 49, 333–369. [Google Scholar]
- Johnson, K.; Goody, R. The Original Michaelis Constant: Translation of the 1913 Michaelis-Menten Paper. Biochemistry 2012, 50, 8264–8269. [Google Scholar] [CrossRef] [Green Version]
- Steen, I.H.; Madern, D.; Karlström, M.; Lien, T.; Ladenstein, R.; Birkeland, N.-K. Comparison of Isocitrate Dehydrogenase from Three Hyperthermophiles Reveals Differences in Thermostability, Cofactor Specificity, Oligomeric State, and Phylogenetic Affiliation. J. Biol. Chem. 2001, 276, 43924–43931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, G.; Golding, G.B.; Dean, A.M. The selective cause of an ancient adaptation. Science 2005, 307, 1279–1282. [Google Scholar] [CrossRef]
- Steen, I.H.; Lien, T.; Birkeland, N.K. Biochemical and phylogenetic characterization of isocitrate dehydrogenase from a hyperthermophilic archaeon, Archaeoglobus fulgidus. Arch. Microbiol. 1997, 168, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Potter, S. Evidence for a dual-specificity isocitrate dehydrogenase in the euryarchaeotan Thermoplasma acidophilum. Can. J. Microbiol. 1993, 39, 262–264. [Google Scholar] [CrossRef]
- Danson, M.J.; Wood, P.A. Isocitrate dehydrogenase of the thermoacidophilic archaebacterium Sulpholobus acidocaldarius. FEBS Lett. 1984, 172, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Dean, A.M.; Koshland, D.E. Kinetic Mechanism of Escherichia coli Isocitrate Dehydrogenase. Biochemistry 1993, 32, 9302–9309. [Google Scholar] [CrossRef] [PubMed]
- Dean, A.M.; Golding, G.B. Protein engineering reveals ancient adaptive replacements in isocitrate dehydrogenase. Proc. Natl. Acad. Sci. USA 1997, 94, 3104–3109. [Google Scholar] [CrossRef] [Green Version]
- Hurley, J.H.; Dean, A.M.; Koshland, D.E.; Stroud, R.M. Catalytic Mechanism of NADP+-Dependent Isocitrate Dehydrogenase: Implications from the Structures of Magnesium-Isocitrate and NADP+ Complexes. Biochemistry 1991, 30, 8671–8678. [Google Scholar] [CrossRef]
- Kondo, H.; Murakami, M. Crystal Structures of the Putative Isocitrate Dehydrogenase from Sulfolobus tokodaii Strain 7 in the Apo and NADP + -Bound Forms. Archaea 2018, 2018, 7571984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlström, M.; Stokke, R.; Helene Steen, I.; Birkeland, N.K.; Ladenstein, R. Isocitrate dehydrogenase from the hyperthermophile Aeropyrum pernix: X-ray structure analysis of a ternary enzyme-substrate complex and thermal stability. J. Mol. Biol. 2005, 345, 559–577. [Google Scholar] [CrossRef] [PubMed]
- Colman, R.F. Role of Metal Ions in Reactions Catalyzed by Pig Heart Triphosphopyridine Nucleotide-dependent Isocitrate Dehydrogenase. J. Biol. Chem. 1972, 247, 215–223. [Google Scholar] [CrossRef]
- Huang, S.-P.; Cheng, H.-M.; Wang, P.; Zhu, G.-P. Biochemical Characterization and Complete Conversion of Coenzyme Specificity of Isocitrate Dehydrogenase from Bifidobacterium longum. Int. J. Mol. Sci. 2016, 17, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoddard, B.L.; Dean, A.; Koshland, D.E. Structure of Isocitrate Dehydrogenase with Isocitrate, Nicotinamide Adenine Dinucleotide Phosphate and Calcium at 2.5-Å Resolution: A Pseudo-Michaelis Ternary Complex. Biochemistry 1993, 32, 9310–9316. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Greer, A.; Dean, A.M. A highly active decarboxylating dehydrogenase with rationally inverted coenzyme specificity. Proc. Natl. Acad. Sci. USA 1995, 92, 11666–11670. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Greer, A.F.; Dean, A.M.; Hurley, J.H. Engineering secondary structure to invert coenzyme specificity in isopropylmalate dehydrogenase. Tech. Protein Chem. 1997, 8, 809–816. [Google Scholar] [CrossRef]
- Stokke, R.; Karlström, M.; Yang, N.; Leiros, I.; Ladenstein, R.; Birkeland, N.K.; Steen, I.H. Thermal stability of isocitrate dehydrogenase from Archaeoglobus fulgidus studied by crystal structure analysis and engineering of chimers. Extremophiles 2007, 11, 481–493. [Google Scholar] [CrossRef] [PubMed]
- Camacho, M.; Rodríguez-Arnedo, A.; Bonete, M.J. NADP-dependent isocitrate dehydrogenase from the halophilic archaeon Haloferax volcanii: Cloning, sequence determination and overexpression in Escherichia coli. FEMS Microbiol. Lett. 2002, 209, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Arnedo, A.; Camacho, M.; Llorca, F.; Bonete, M.J. Complete reversal of coenzyme specificity of isocitrate dehydrogenase from Haloferax volcanii. Protein J. 2005, 24, 259–266. [Google Scholar] [CrossRef]
- Jin, M.M.; Wang, P.; Li, X.; Zhao, X.Y.; Xu, L.; Song, P.; Zhu, G.P. Biochemical characterization of NADP+-dependent isocitrate dehydrogenase from Microcystis aeruginosa PCC7806. Mol. Biol. Rep. 2013, 40, 2995–3002. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Nandyala, A.; Podili, R.; Katoch, V.M.; Hasnain, S.E. Comparison of Mycobacterium tuberculosis isocitrate dehydrogenases (ICD-1 and ICD-2) reveals differences in coenzyme affinity, oligomeric state, pH tolerance and phylogenetic affiliation. BMC Biochem. 2005, 6, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrenger, C.; Müller, S. Isocitrate dehydrogenase of Plasmodium falciparum: Energy metabolism or redox control? Eur. J. Biochem. 2003, 270, 1775–1783. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.; Colman, R.F. Thr373, Asp375, and Lys260 are in the coenzyme site of porcine NADP-dependent isocitrate dehydrogenase. Arch. Biochem. Biophys. 2006, 450, 183–190. [Google Scholar] [CrossRef]
- Plaut, G.W.E. Isocitrate Dehydrogenase. In The Enzymes; Boyer, P.D., Lardy, H., Myrbäck, K., Eds.; Academic Press: New York, NY, USA, 1963; pp. 105–126. [Google Scholar]
- Stokke, R.; Birkeland, N.K.; Steen, I.H. Thermal stability and biochemical properties of isocitrate dehydrogenase from the thermoacidophilic archaeon Thermoplasma acidophilum. Extremophiles 2007, 11, 397–402. [Google Scholar] [CrossRef]
- Li, X.; Wang, P.; Ge, Y.; Wang, W.; Abbas, A.; Zhu, G. NADP+-specific isocitrate dehydrogenase from oleaginous yeast Yarrowia lipolytica CLIB122: Biochemical characterization and coenzyme sites evaluation. Appl. Biochem. Biotechnol. 2013, 171, 403–416. [Google Scholar] [CrossRef]





| No. | Orientation | Sequence (5′ → 3′) 1 |
|---|---|---|
| 1 | forward | GTT TAA CTT TAA GAA GGA GAT ATA CAT ACC ATG CAC CAT CAT CAC CAC CAT GAA GAA CAG AAA AAA GAA TCA ATA AG |
| 2 | reverse | CCG CCA AAA CAG CCA AGC TGG AGA CCG TTT TCA TGC TGA TTT TAT CGC |
| 3 | forward | AAA CGG TCT CCA GCT TG |
| 4 | reverse | GGT ATG TAT ATC TCC TTC TTA AAG TTA AAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Winkler, D.; Gfrerer, S.; Gescher, J. Biochemical Characterization of Recombinant Isocitrate Dehydrogenase and Its Putative Role in the Physiology of an Acidophilic Micrarchaeon. Microorganisms 2021, 9, 2318. https://doi.org/10.3390/microorganisms9112318
Winkler D, Gfrerer S, Gescher J. Biochemical Characterization of Recombinant Isocitrate Dehydrogenase and Its Putative Role in the Physiology of an Acidophilic Micrarchaeon. Microorganisms. 2021; 9(11):2318. https://doi.org/10.3390/microorganisms9112318
Chicago/Turabian StyleWinkler, Dennis, Sabrina Gfrerer, and Johannes Gescher. 2021. "Biochemical Characterization of Recombinant Isocitrate Dehydrogenase and Its Putative Role in the Physiology of an Acidophilic Micrarchaeon" Microorganisms 9, no. 11: 2318. https://doi.org/10.3390/microorganisms9112318
APA StyleWinkler, D., Gfrerer, S., & Gescher, J. (2021). Biochemical Characterization of Recombinant Isocitrate Dehydrogenase and Its Putative Role in the Physiology of an Acidophilic Micrarchaeon. Microorganisms, 9(11), 2318. https://doi.org/10.3390/microorganisms9112318