
Ecology of Powassan Virus in the United States



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
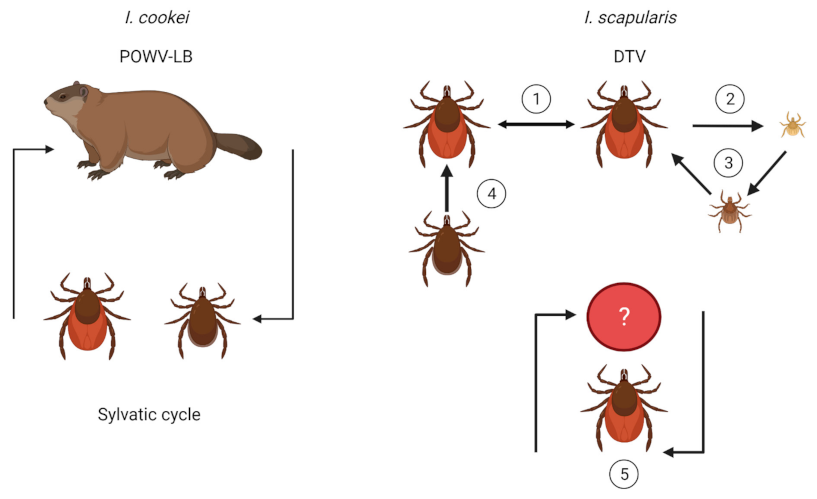
2. Vector and Host Associations
3. Transmission Dynamics
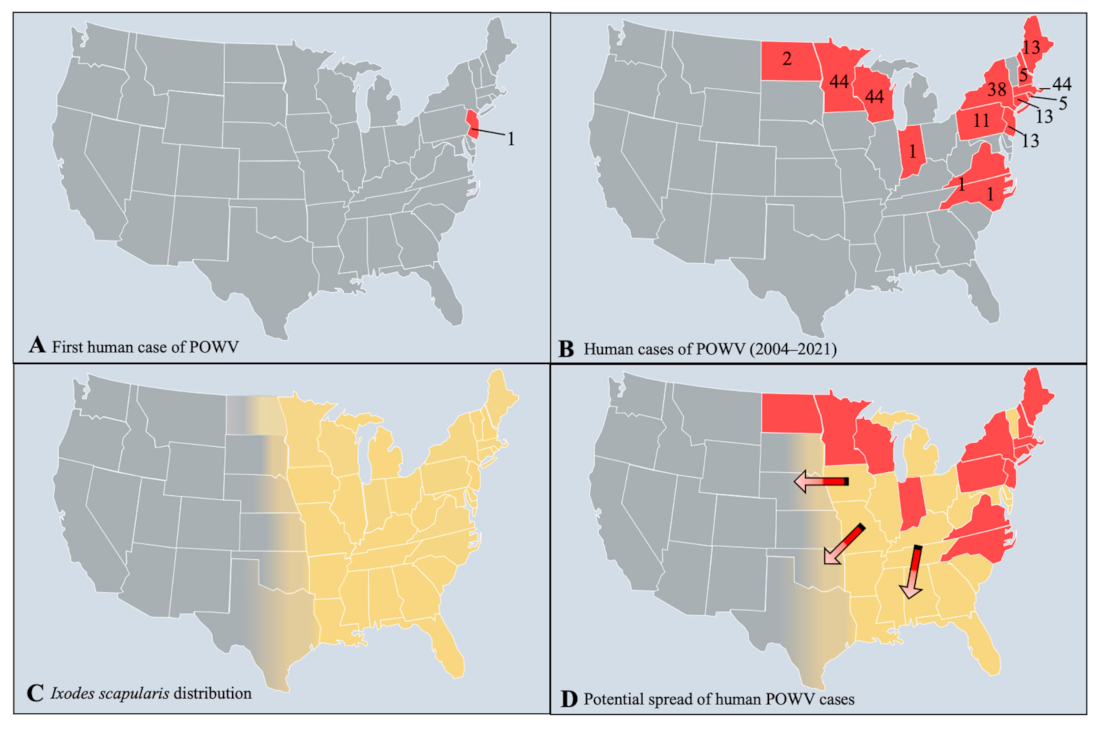
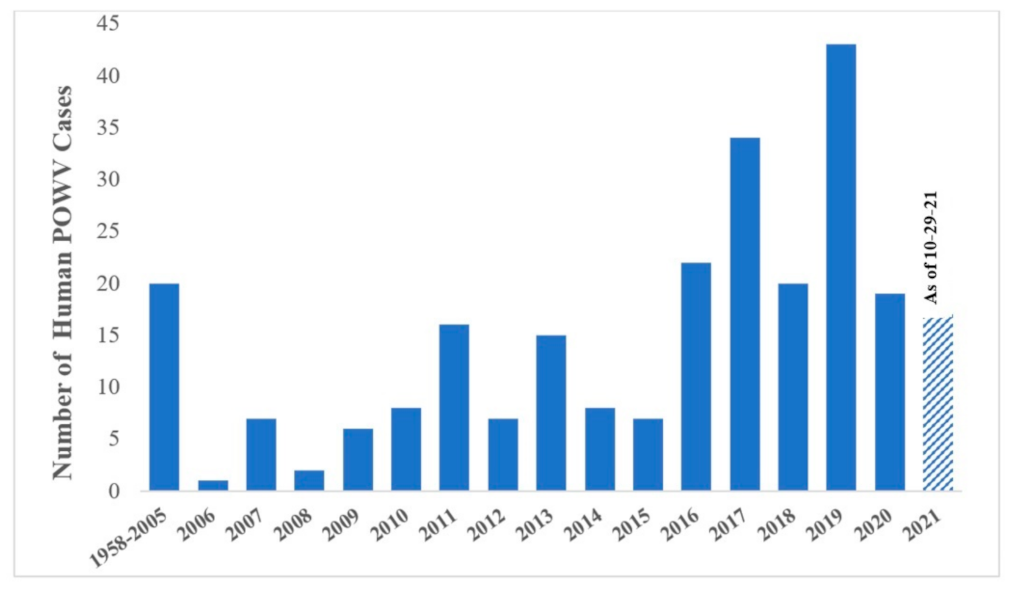
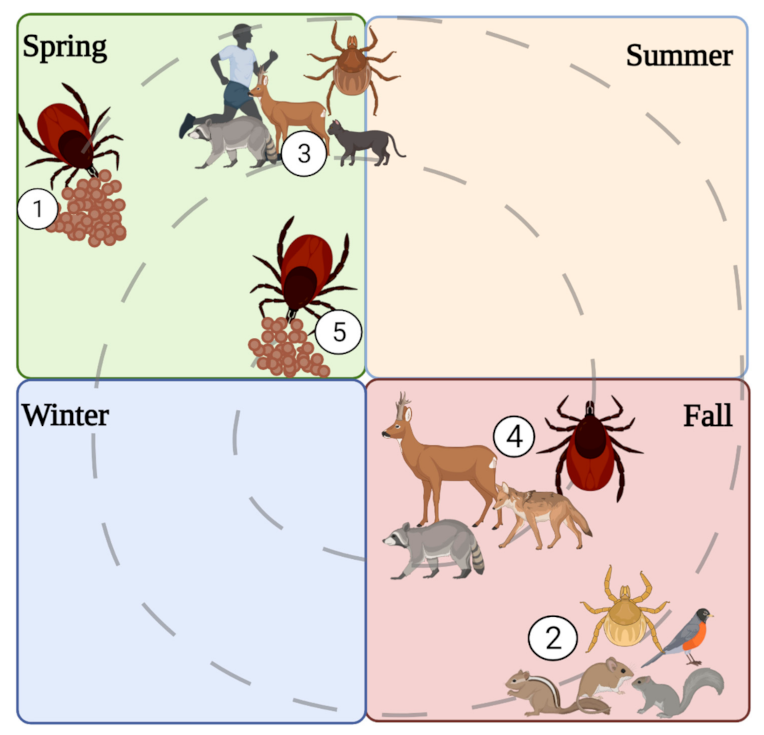
4. Spatial, Temporal, Habitat, and Meteorological Associations
5. Strain Variation and Stability
6. Climate Change and Anthropogenic Influence
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 21 September 2021).
- Sonenshine, D.E. Range expansion of tick disease vectors in north america: Implications for spread of tick-borne disease. Int. J. Environ. Res. Public Health 2018, 15, 478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diuk-Wasser, M.A.; VanAcker, M.C.; Fernandez, M.P. Impact of Land Use Changes and Habitat Fragmentation on the Eco-epidemiology of Tick-Borne Diseases. J. Med. Entomol. 2021, 58, 1546–1564. [Google Scholar] [CrossRef]
- Gould, E.A.; Higgs, S. Impact of climate change and other factors on emerging arbovirus diseases. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Paddock, C.; Lane, R.; Staples, J.; Labruna, M. Changing paradigms for tick-borne diseases in the Americas. In Global Health Impacts of Vector-Borne Diseases; National Academies Press (US): Washington, DC, USA, 2016. [Google Scholar] [CrossRef]
- CDC. How Many People Get Lyme Disease? 2021. Available online: https://www.cdc.gov/lyme/stats/humancases.html (accessed on 7 June 2021).
- Paules, C.; Marston, H.; Bloom, M.; Fauci, A. Tickborne Diseases—Confronting a Growing Threat. N. Engl. J. Med. 2018, 701–703. [Google Scholar] [CrossRef]
- McLean, D.M.; Donohue, W.L. Powassan virus: Isolation of virus from a fatal case of encephalitis. Can. Med. Assoc. J. 1959, 80, 708–711. [Google Scholar] [PubMed]
- Goldfield, M.; Austin, S.M.; Black, H.C.; Taylor, B.F.; Altman, R. A non-fatal human case of Powassan virus encephalitis. Am. J. Trop. Med. Hyg. 1973, 22, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Telford, S.R.; Armstrong, P.M.; Katavolos, P.; Foppa, I.; Garcia, A.S.O.; Wilson, M.L.; Spielman, A. A new tick-borne encephalitis-like virus infecting New England deer ticks, Ixodes dammini. Emerg. Infect. Dis. 1997, 10, 156–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuno, G.; Artsob, H.; Karabatsos, N.; Tsuchiya, K.R.; Chang, G.J.J. Genomic sequencing of deer tick virus and phylogeny of Powassan-related viruses of North America. Am. J. Trop. Med. Hyg. 2001, 65, 671–676. [Google Scholar] [CrossRef] [Green Version]
- Beasley, D.W.C.; Suderman, M.T.; Holbrook, M.R.; Barrett, A.D.T. Nucleotide sequencing and serological evidence that the recently recognized deer tick virus is a genotype of Powassan virus. Virus Res. 2001, 79, 81–89. [Google Scholar] [CrossRef]
- Krow-Lucal, E.R.; Lindsey, N.P.; Fischer, M.; Hills, S.L. Powassan virus disease in the United States, 2006–2016. Vector-Borne Zoonotic Dis. 2018, 18, 286–290. [Google Scholar] [CrossRef] [Green Version]
- Leonova, G.N.; Kondratov, I.G.; Ternovoi, V.A.; Romanova, E.V.; Protopopova, E.V.; Chausov, E.V.; Pavlenko, E.V.; Ryabchikova, E.I.; Belikov, S.I.; Loktev, V.B. Characterization of Powassan viruses from Far Eastern Russia. Arch. Virol. 2009, 154, 811–820. [Google Scholar] [CrossRef] [PubMed]
- CDC. Powassan Virus. Centers for Disease Control and Prevention. 2019. Available online: https://www.cdc.gov/powassan/statistics.html (accessed on 7 June 2021).
- CDC. Powassan Virus. 2021. Available online: https://wwwn.cdc.gov/arbonet/Maps/ADB_Diseases_Map/index.html (accessed on 7 June 2021).
- CDC. Arboviral Diseases, Neuroinvasive and Non-Neuroinvasive 2015 Case Definition. 2021. Available online: https://ndc.services.cdc.gov/case-definitions/arboviral-diseases-neuroinvasive-and-non-neuroinvasive-2015/ (accessed on 3 November 2021).
- Hinten, S.R.; Beckett, G.A.; Gensheimer, K.F.; Pritchard, E.; Courtney, T.M.; Sears, S.D.; Woytowicz, J.M.; Preston, D.G.; Smith, R.P.; Rand, P.W.; et al. Increased recognition of powassan encephalitis in the United States, 1999–2005. Vector-Borne Zoonotic Dis. 2008, 8, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Corrin, T.; Greig, J.; Harding, S.; Young, I.; Mascarenhas, M.; Waddell, L.A. Powassan virus, a scoping review of the global evidence. Zoonoses Public Health 2018, 65, 595–624. [Google Scholar] [CrossRef]
- Ebel, G.D. Update on Powassan virus: Emergence of a North American tick-borne flavivirus. Annu. Rev. Entomol. 2010, 55, 95–110. [Google Scholar] [CrossRef]
- Hermance, M.E.; Thangamani, S. Powassan virus: An emerging arbovirus of public health concern in North America. Vector-Borne Zoonotic Dis. 2017, 17, 453–462. [Google Scholar] [CrossRef]
- El Khoury, M.Y.; Camargo, J.F.; White, J.L.; Backenson, B.P.; Dupuis, A.P.; Escuyer, K.L.; Kramer, L.; George, K.S.; Chatterjee, D.; Prusinski, M.; et al. Potential Role of Deer Tick Virus in Powassan Encephalitis Cases in Lyme Disease–endemic Areas of New York, USA. Emerg. Infect. Dis. 2013, 19, 1926–1933. [Google Scholar] [CrossRef] [Green Version]
- Campbell, O.; Krause, P.J. The emergence of human Powassan virus infection in North America. Ticks Tick. Borne. Dis. 2020, 11, 101540. [Google Scholar] [CrossRef]
- Mclean, D.M.; Walker, S.J.; Macpherson, L.W.; Scholten, T.H.; Ronald, K.; Wyllie, J.C.; Mcqueen, E.J. Powassan Virus: Investigations of possible natural cycles of infection. J. Infect. Dis. 1961, 109, 19–23. [Google Scholar] [CrossRef]
- McLean, D.M.; Larke, R.P. Powassan and Silverwater viruses: Ecology of two Ontario arboviruses. Can. Med. Assoc. J. 1963, 88, 182–185. [Google Scholar]
- Mclean, D.M.; Best, J.M.; Sc, B.; Ass, J. Powassan virus: Summer infection cycle, 1964. Can. Med. Assoc. J. 1964, 91, 1360–1362. [Google Scholar]
- Main, A.J.; Carey, A.B.; Downs, W.G.; Haven, N. Powassan virus in Ixodes cookei and mustelidae in New England. J. Wildl. Dis. 1979, 15. [Google Scholar] [CrossRef] [Green Version]
- McLean, D.M.; Cobb, C.; Gooderham, S.E.; Smart, C.A.; Wilson, A.G.; Wilson, W.E. Powassan virus: Persistence of virus activity during 1966. Can. Med. Assoc. J. 1967, 96, 660–664. [Google Scholar]
- Mclean, D.M.; de Vos, A.; Quantz, J. Powassan virus: Field investigations of 1963. Am. J. Trop. Med. Hyg. 1964, 13, 747–753. [Google Scholar] [CrossRef]
- Ebel, G.D.; Campbell, E.N.; Goethert, H.K.; Spielman, A.; Telford, S.R. Enzootic transmission of deer tick virus in new England and Wisconsin sites. Am. J. Trop. Med. Hyg. 2000, 63, 36–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brackney, D.E.; Nofchissey, R.A.; Fitzpatrick, K.A.; Brown, I.K.; Ebel, G.D. Short report: Stable prevalence of Powassan virus in Ixodes scapularis in a Northern Wisconsin focus. Am. J. Trop. Med. Hyg. 2008, 79, 971–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokarz, R.; Tagliafierro, T.; Cucura, D.M.; Rochlin, I.; Sameroff, S.; Lipkin, W.I. Detection of Anaplasma phagocytophilum, Babesia microti, Borrelia burgdorferi, Borrelia miyamotoi, and Powassan Virus in Ticks by a Multiplex Real-Time Reverse Transcription-PCR Assay. mSphere 2017, 2, e00151-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitney, E.; Jamnback, H. The first isolations of Powassan virus in New York State. Proc. Soc. Exp. Biol. Med. 1965, 119, 432–435. [Google Scholar] [CrossRef] [PubMed]
- Thomas, L.A.; Kennedy, R.C.; Eklund, C.M. Isolation of a virus closely related to Powassan virus from Dermacentor andersoni collected along North Cache la Poudre River, Colorado. Proc. Soc. Exp. Biol. Med. 1960, 104, 355–359. [Google Scholar] [CrossRef]
- CDC. Geographic Distribution of Ticks that Bite Humans. 2019. Available online: https://www.cdc.gov/ticks/geographic_distribution.html (accessed on 3 November 2021).
- Johnson, H.N. Isolation of Powassan virus from a spotted skunk in California. J. Wildl. Dis. 1987, 23, 152–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupuis, A.P.; Peters, R.J.; Prusinski, M.A.; Falco, R.C.; Ostfeld, R.S.; Kramer, L.D. Isolation of deer tick virus (Powassan virus, lineage II) from Ixodes scapularis and detection of antibody in vertebrate hosts sampled in the Hudson Valley, New York State. Parasites Vectors 2013, 6, 185. [Google Scholar] [CrossRef] [Green Version]
- Deardorff, E.R.; Nofchissey, R.A.; Cook, J.A.; Hope, A.G.; Tsvetkova, A.; Talbot, S.L.; Ebel, G.D. Powassan Virus in mammals, Alaska and New Mexico, USA, and Russia, 2004–2007. Emerg. Infect. Dis. 2013, 19, 1–5. [Google Scholar] [CrossRef]
- Sprague, A. Down the Woodchuck Hole: Investigating the Potential for Peromyscus Leucopus to Act as a Mixing Vessel for the Powassan Virus and Deer Tick Virus Disease Systems (Issue Usgs); University of Massachusetts: Amherst, MA, USA, 2010. [Google Scholar]
- Frey, S.; Essbauer, S.; Zöller, G.; Klempa, B.; Dobler, G.; Pfeffer, M. Full genome sequences and preliminary molecular characterization of three tick-borne encephalitis virus strains isolated from ticks and a bank vole in Slovak Republic. Virus Genes 2014, 48, 184–188. [Google Scholar] [CrossRef]
- Weidmann, M.; Schmidt, P.; Hufert, F.T.; Krivanec, K.; Meyer, H. Tick-borne encephalitis virus in Clethrionomys glareolus in the Czech Republic. Vector-Borne Zoonotic Dis. 2006, 6, 379–381. [Google Scholar] [CrossRef] [PubMed]
- Tonteri, E.; Kipar, A.; Voutilainen, L.; Vene, S.; Vaheri, A.; Vapalahti, O.; Lundkvist, Å. The three subtypes of tick-borne encephalitis virus induce encephalitis in a natural host, the bank vole (Myodes glareolus). PLoS ONE 2013, 8, e81214. [Google Scholar]
- Burri, C.; Korva, M.; Bastic, V.; Knap, N.; Avšič-Županc, T.; Gern, L. Serological Evidence of Tick-Borne Encephalitis Virus Infection in Rodents Captured at Four Sites in Switzerland. J. Med. Entomol. 2012, 49, 436–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zöldi, V.; Papp, T.; Reiczigel, J.; Egyed, L. Bank voles show high seropositivity rates in a natural TBEV focus in Hungary. Infect. Dis. 2015, 47, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Mlera, L.; Bloom, M.E. The role of mammalian reservoir hosts in tick-borne flavivirus biology. Front. Cell. Infect. Microbiol. 2018, 8. [Google Scholar] [CrossRef]
- Ebel, G.D.; Kramer, L.D. Short report: Duration of tick attachment required for transmission of powassan virus by deer ticks. Am. J. Trop. Med. Hyg. 2004, 71, 268–271. [Google Scholar] [CrossRef] [Green Version]
- Hermance, M.E.; Thangamani, S. Tick−Virus−Host Interactions at the Cutaneous Interface: The Nidus of Flavivirus Transmission. Viruses 2018, 10, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feder, H.M.; Telford, S.; Goethert, H.K.; Wormser, G.P. Powassan virus encephalitis following brief attachment of Connecticut deer ticks. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Spielman, A.; Ribeiro, J.M.C.; Mather, T.N.; Piesman, J. Dissemination and salivary delivery of Lyme disease spirochetes in vector ticks (Acari: Ixodidae). J. Med. Entomol. 1987, 24, 201–205. [Google Scholar] [CrossRef]
- Hermance, M.E.; Thangamani, S. Tick saliva enhances Powassan virus transmission to the host, influencing its dissemination and the course of disease. J. Virol. 2015, 89, 7852–7860. [Google Scholar] [CrossRef] [Green Version]
- Costero, A.; Grayson, M.A. Experimental transmission of Powassan virus (Flaviviridae) by Ixodes scapularis ticks (Acari:Ixodidae). Am. J. Trop. Med. Hyg. 1996, 55, 536–546. [Google Scholar] [CrossRef]
- Woodall, J.P.; Roz, A. Experimental milk-borne transmission of Powassan virus in the goat. Am. J. Trop. Med. Hyg. 1977, 26, 190–192. [Google Scholar] [CrossRef] [PubMed]
- Dorko, E.; Hockicko, J.; Rimárová, K.; Bušová, A.; Popaďák, P.; Popaďáková, J.; Schréter, I. Milk outbreaks of tick-borne encephalitis in Slovakia, 2012-2016. Cent. Eur. J. Public Health 2018, 26, S47–S50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cisak, E.; Wójcik-Fatla, A.; Zając, V.; Sroka, J.; Buczek, A.; Dutkiewicz, J. Prevalence of tick-borne encephalitis virus (TBEV) in samples of raw milk taken randomly from cows, goats and sheep in eastern Poland. Ann. Agric. Environ. Med. 2010, 17, 283–286. [Google Scholar]
- Jones, L.D.; Davies, C.R.; Steele, G.M.; Nuttall, P.A. A novel mode of arbovirus transmission involving a nonviremic host. Science 1987, 237, 775–777. [Google Scholar] [CrossRef] [PubMed]
- Havlíková, S.; Licková, M.; Klempa, B. Non-viraemic transmission of tick-borne viruses. Acta Virol. 2013, 57, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Labuda, M.; Danielova, V.; Jones, L.D.; Nuttall, P.A. Amplification of tick-borne encephalitis virus infection during co-feeding of ticks. Med. Vet. Entomol. 1993, 7, 339–342. [Google Scholar] [CrossRef]
- Labuda, M.; Jones, L.D.; Williams, T.; Danielova, V.; Nuttall, P.A. Efficient transmission of tick-borne encephalitis virus between cofeeding ticks. J. Med. Entomol. 1993, 30, 295–299. [Google Scholar] [CrossRef]
- Labuda, M.; Kozuch, O.; Zuffová, E.; Elecková, E.; Hails, R.S.; Nuttall, P.A. Tick-borne encephalitis virus transmission between ticks cofeeding on specific immune natural rodent hosts. Virology 1997, 235, 138–143. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Balzano, C.; Hess, M.; Malhotra, A.; Lenox, R. Severe babesiosis and Borrelia burgdorferi co-infection. QJM An. Int. J. Med. 2015, 108, 141–143. [Google Scholar] [CrossRef] [Green Version]
- Vinasco, J.; Braga, W.; Zegarra-Moro, O.; Moro, M.H. Cellular immune responses in a murine model of Borrelia burgdorferi and Babesia microti coinfection. J. Immunol. 2007, 178 (Suppl. 1), S47 LP-S47. Available online: http://www.jimmunol.org/content/178/1_Supplement/S47.3.abstract (accessed on 13 October 2021).
- Dunn, J.M.; Krause, P.J.; Davis, S.; Vannier, E.G.; Fitzpatrick, M.C.; Rollend, L.; Belperron, A.A.; States, S.L.; Stacey, A.; Bockenstedt, L.K.; et al. Borrelia burgdorferi promotes the establishment of Babesia microti in the Northeastern United States. PLoS ONE 2015, 9, e115494. [Google Scholar] [CrossRef]
- Charles, A.S.; Christofferson, R.C. Utility of a dengue-derived monoclonal antibody to enhance Zika infection in vitro. PLoS Curr. 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Durbin, A.P. Dengue antibody and Zika: Friend or foe? Trends Immunol. 2016, 37, 635–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, L.M.; Carlin, E.R.; Jenkins, M.M.; Tan, A.L.; Barcellona, C.M.; Nicholson, C.O.; Michael, S.F.; Isern, S. Dengue virus antibodies enhance Zika virus infection. Clin. Transl. Immunol. 2016, 5, e117. [Google Scholar] [CrossRef] [PubMed]
- Lehane, A.; Maes, S.E.; Graham, C.B.; Jones, E.; Delorey, M.; Eisen, R.J. Prevalence of single and coinfections of human pathogens in Ixodes ticks from five geographical regions in the United States, 2013–2019. Ticks Tick-Borne Dis. 2021, 12, 101637. [Google Scholar] [CrossRef]
- Ko, R.C. Biology of Ixodes cookei Packard (Ixodidae) of groundhogs (Marmota monax Erxleben). Can. J. Zool. 1972, 50, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Rand, P.W.; Lacombe, E.H.; Dearborn, R.; Cahill, B.; Elias, S.; Lubelczyk, C.B.; Beckett, G.A.; Smith, R.P. Passive surveillance in Maine, an area emergent for tick-borne diseases. J. Med. Entomol. 2007, 44, 1118–1129. [Google Scholar] [CrossRef] [PubMed]
- Pak, D.; Jacobs, S.B.; Sakamoto, J.M. A 117-year retrospective analysis of Pennsylvania tick community dynamics. Parasites Vectors 2019, 12, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisen, R.J.; Eisen, L.; Beard, C.B. County-scale distribution of Ixodes scapularis and Ixodes pacificus (Acari: Ixodidae) in the continental United States. J. Med. Entomol. 2016, 53, 349–386. [Google Scholar] [CrossRef] [Green Version]
- Alkishe, A.; Raghavan, R.K.; Peterson, A.T. Likely geographic distributional shifts among medically important tick species and tick-associated diseases under climate change in North America: A review. Insects 2021, 12, 225. [Google Scholar] [CrossRef] [PubMed]
- Simmons, T.W.; Shea, J.; Myers-Claypole, M.A.; Kruise, R.; Hutchinson, M.L. Seasonal activity, density, and collection efficiency of the blacklegged tick (Ixodes scapularis) (Acari: Ixodidae) in Mid-Western Pennsylvania. J. Med. Entomol. 2015, 52, 1260–1269. [Google Scholar] [CrossRef]
- CDC. How Ticks Spread Disease. 2020. Available online: https://www.cdc.gov/ticks/life_cycle_and_hosts.html (accessed on 10 October 2021).
- Guerra, M.; Walker, E.; Jones, C.; Paskewitz, S.; Roberto Cortinas, M.; Ashley Stancil, L.B.; Bobo, M.; Kitron, U. Predicting the risk of Lyme disease: Habitat suitability for Ixodes scapularis in the north central United States. Emerg. Infect. Dis. 2002, 8, 289–297. [Google Scholar] [CrossRef]
- Guglielmone, A.A.; Robbins, R.G.; Apanaskevich, D.A.; Petney, T.N.; Estrada-Peña, A.; Horak, I.G. The Hard Ticks of the World; Springer: Dordrecht, The Netherlands, 2014. [Google Scholar] [CrossRef]
- Clotfelter, E.D.; Pedersen, A.B.; Cranford, J.A.; Ram, N.; Snajdr, E.A.; Nolan, V.; Ketterson, E.D. Acorn mast drives long-term dynamics of rodent and songbird populations. Oecologia 2007, 154, 493–503. [Google Scholar] [CrossRef]
- Schulze, T.L.; Jordan, R.A.; Hung, R.W. Effects of selected meteorological factors on diurnal questing of Ixodes scapularis and Amblyomma americanum (Acari: Ixodidae). J. Med. Entomol. 2001, 38, 318–324. [Google Scholar] [CrossRef]
- Burtis, J.C.; Sullivan, P.; Levi, T.; Oggenfuss, K.; Fahey, T.J.; Ostfeld, R.S. The impact of temperature and precipitation on blacklegged tick activity and Lyme disease incidence in endemic and emerging regions. Parasites Vectors 2016, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elias, S.P.; Lubelczyk, C.B.; Rand, P.W.; Lacombe, E.H.; Holman, M.S.; Smith Jr, R.P. Deer browse resistant exotic-invasive understory: An indicator of elevated human risk of exposure to Ixodes scapularis (Acari: Ixodidae) in southern coastal Maine woodlands. J. Med. Entomol. 2006, 43, 1142–1152. [Google Scholar] [CrossRef]
- Pesko, K.N.; Torres-Perez, F.; Hjelle, B.L.; Ebel, G.D. Molecular epidemiology of Powassan virus in North America. J. Gen. Virol. 2010, 91, 2698–2705. [Google Scholar] [CrossRef]
- Anderson, J.F.; Armstrong, P.M. Prevalence and genetic characterization of Powassan virus strains infecting Ixodes scapularis in Connecticut. Am. J. Trop. Med. Hyg. 2012, 87, 754–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borde, J.P.; Kaier, K.; Hehn, P.; Matzarakis, A.; Frey, S.; Bestehorn, M.; Dobler, G.; Chitimia-Dobler, L. The complex interplay of climate, TBEV vector dynamics and TBEV infection rates in ticks—Monitoring a natural TBEV focus in Germany, 2009–2018. PLoS ONE 2021, 16, e0244668. [Google Scholar] [CrossRef] [PubMed]
- Jorgenson, A.K. Global warming and the neglected greenhouse gas: A cross-national study of the social causes of methane emissions intensity, 1995. Soc. Forces 2006, 84, 1779–1798. [Google Scholar] [CrossRef]
- Ogden, N.H.; Ben Beard, C.; Ginsberg, H.S.; Tsao, J.I. Possible effects of climate change on Ixodid ticks and the pathogens they transmit: Predictions and observations. J. Med. Entomol. 2021, 58, 1536–1545. [Google Scholar] [CrossRef] [PubMed]
- Ebi, K.L.; Vanos, J.; Baldwin, J.W.; Bell, J.E.; Hondula, D.M.; Errett, N.A.; Hayes, K.; Reid, C.E.; Saha, S.; Spector, J.; et al. Extreme weather and climate change: Population health and health system implications. Annu. Rev. Public Health 2020, 42, 293–315. [Google Scholar] [CrossRef]
- Roy-Dufresne, E.; Logan, T.; Simon, J.A.; Chmura, G.L.; Millien, V. Poleward expansion of the white-footed mouse (Peromyscus leucopus) under climate change: Implications for the spread of lyme disease. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [Green Version]
- Caignard, T.; Kremer, A.; Firmat, C.; Nicolas, M.; Venner, S.; Delzon, S. Increasing spring temperatures favor oak seed production in temperate areas. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Zervanos, S.M.; Maher, C.R.; Waldvogel, J.A.; Florant, G.L. Latitudinal differences in the hibernation characteristics of woodchucks (Marmota monax). Physiol. Biochem. Zool. 2010, 83, 135–141. [Google Scholar] [CrossRef] [Green Version]
- EDDMapS. Japanese Barberry. 2021. Available online: https://www.eddmaps.org/distribution/uscounty.cfm?sub=3010 (accessed on 10 October 2021).
- Williams, S.C.; Ward, J.S. Effects of Japanese barberry (Ranunculales: Berberidaceae) removal and resulting microclimatic changes on Ixodes scapularis (Acari: Ixodidae) abundances in Connecticut, USA. Environ. Entomol. 2010, 39, 1911–1921. [Google Scholar] [CrossRef]
- Vessey, S.; Vessey, K.B. Linking behavior, life history and food supply with the population dynamics of white-footed mice (Peromyscus leucopus). Integr. Zool. 2007, 2, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Wolff, J.O.; Schauder, E.M.; Daniel Edge, W. Effects of habitat loss and fragmentation on the behavior and demography of gray-tailed voles. Conserv. Biol. 1997, 11, 945–956. [Google Scholar] [CrossRef] [Green Version]
- Ostfeld, R.S.; Keesing, F. Is biodiversity bad for your health? Ecosphere 2015, 8, 17–19. [Google Scholar] [CrossRef]
- Allan, B.F.; Keesing, F.; Ostfeld, R.S. Effect of Forest Fragmentation on Lyme Disease Risk. Conserv. Biol. 2003, 17, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Stafford, K.C. Tick Management Handbook. The Connecticut Agricultural Experiment Station, 71. 2004. Available online: http://www.ct.gov/caes/lib/caes/documents/special_features/tickhandbook.pdf (accessed on 10 July 2021).





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassett, E.M.; Thangamani, S. Ecology of Powassan Virus in the United States. Microorganisms 2021, 9, 2317. https://doi.org/10.3390/microorganisms9112317
Hassett EM, Thangamani S. Ecology of Powassan Virus in the United States. Microorganisms. 2021; 9(11):2317. https://doi.org/10.3390/microorganisms9112317
Chicago/Turabian StyleHassett, Erin M., and Saravanan Thangamani. 2021. "Ecology of Powassan Virus in the United States" Microorganisms 9, no. 11: 2317. https://doi.org/10.3390/microorganisms9112317
APA StyleHassett, E. M., & Thangamani, S. (2021). Ecology of Powassan Virus in the United States. Microorganisms, 9(11), 2317. https://doi.org/10.3390/microorganisms9112317