
Proteomic Analysis Explores Interactions between Lactiplantibacillus plantarum and Saccharomyces cerevisiae during Sourdough Fermentation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fermentation and Growth Determination
2.2. Determination of pH and TTA
2.3. Microbial Diversity
2.4. Protein Extraction
2.5. Protein Identification
2.6. Statistical Analysis
3. Results and Discussion
3.1. Viable Cell Counts, pH and TTA
3.2. Microbial Diversity of Different Sourdough
3.3. Proteomic
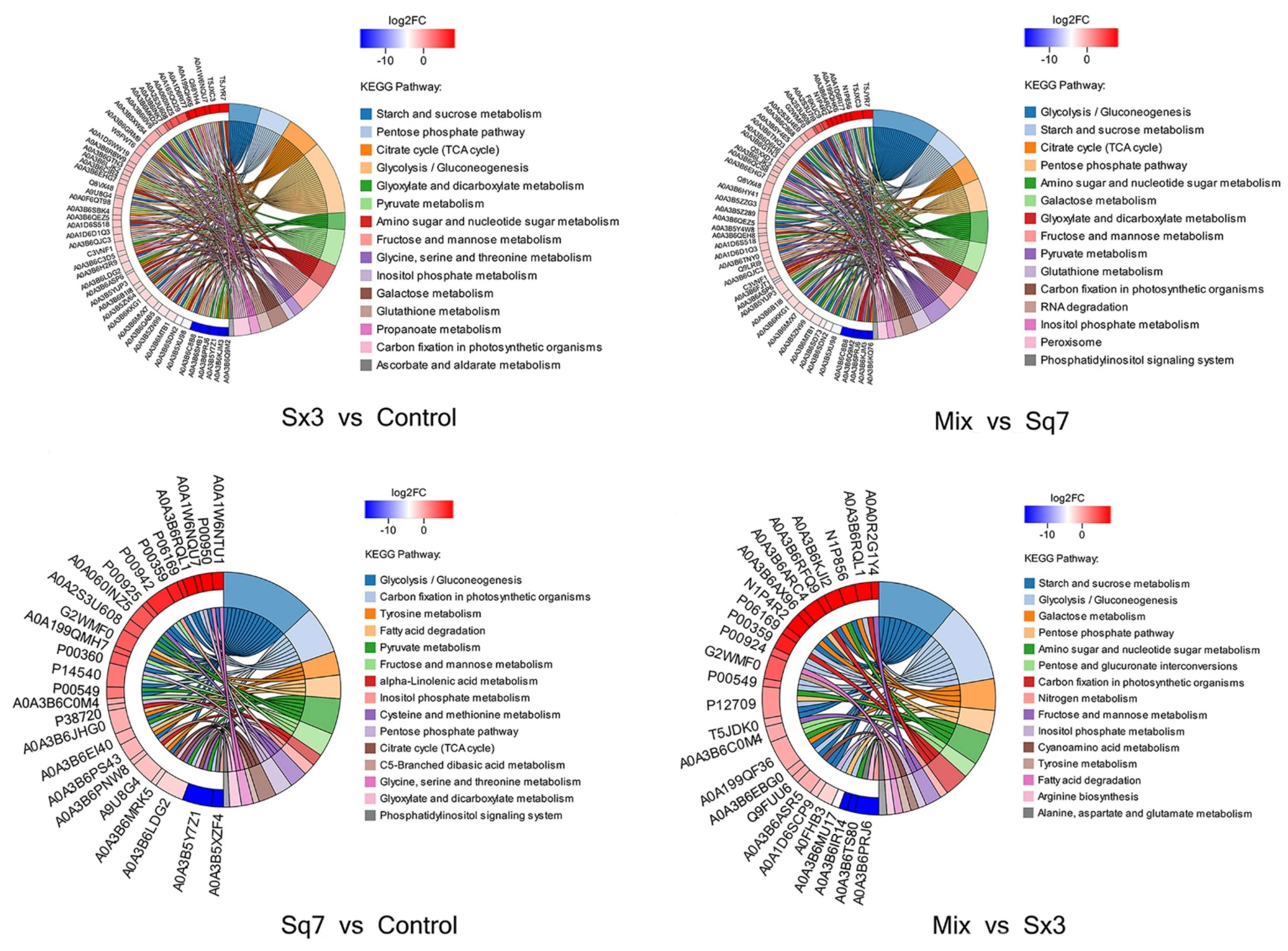
3.3.1. Carbohydrate Metabolism
Starch and Sucrose Metabolism
Glycolysis
TCA-Cycle of Mix Sourdough
3.3.2. Amino Acid Metabolism
3.3.3. Protein Translation
3.3.4. Protein Folding, Sorting and Degradation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oshiro, M.; Zendo, T.; Nakayama, J. Diversity and dynamics of sourdough lactic acid bacteriota created by a slow food fermentation system. J. Biosci. Bioeng. 2021, 131, 333–340. [Google Scholar] [CrossRef]
- Lau, S.W.; Chong, A.Q.; Chin, N.L.; Talib, R.A.; Basha, R.K. Sourdough microbiome comparison and benefits. Microorganisms 2021, 9, 1355. [Google Scholar] [CrossRef]
- Menezes, L.A.A.; SavoSardaro, M.L.; Duarte, R.T.D.; Mazzon, R.R.; Neviani, E.; Gatti, M.; De Dea Lindner, J. Sourdough bacterial dynamics revealed by metagenomic analysis in Brazil. Microorganisms 2020, 85, 103302. [Google Scholar] [CrossRef]
- De Vuyst, L.; Neysens, P. The sourdough microflora: Biodiversity and metabolic interactions. Trends Food Sci. Tech. 2005, 16, 43–56. [Google Scholar] [CrossRef]
- Zhu, J.; Huang, R.; Hu, D.X.; Dou, Y.; Ren, H.Y.; Yang, Z.X.; Deng, C.; Xiong, W.; Wang, D.; Mao, Y. Individualized prediction of metastatic involvement of lymph nodes posterior to the right recurrent laryngeal nerve in papillary thyroid carcinoma. OncoTargets Ther. 2019, 12, 9077–9084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.H.; Tu, J.; Sadiq, F.A.; Zhang, W.Z.; Wang, W. Prevalence, genetic diversity, and technological functions of the Lactobacillus sanfranciscensis in sourdough: A Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1209–1226. [Google Scholar] [CrossRef] [Green Version]
- Chavan, R.S.; Chavan, S.R. Sourdough technology-a traditional way for wholesome foods: A review. Compr. Rev. Food Sci. Food Saf. 2011, 10, 170–183. [Google Scholar] [CrossRef]
- De Vuyst, L.; Harth, H.; Van Kerrebroeck, S.; Leroy, F. Yeast diversity of sourdoughs and associated metabolic properties and functionalities. Int. J. Food Microbiol. 2016, 239, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Pontonio, E.; Di Cagno, R.; Mahony, J.; Lanera, A.; De Angelis, M.; van Sinderen, D.; Gobbetti, M. Sourdough authentication: Quantitative PCR to detect the lactic acid bacterial microbiota in breads. Sci. Rep. 2017, 7, 624. [Google Scholar] [CrossRef]
- Pulvirenti, A.; Solieri, L.; Gullo, M.; De Vero, L.; Giudici, P. Occurrence and dominance of yeast species in sourdough. Lett. Appl. Microbiol. 2004, 38, 113–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vuyst, L.; Van Kerrebroeck, S.; Harth, H.; Huys, G.; Daniel, H.M.; Weckx, S. Microbial ecology of sourdough fermentations: Diverse or uniform? Food Microbiol. 2014, 37, 11–29. [Google Scholar] [CrossRef] [PubMed]
- Van Rossum, H.M.; Kozak, B.U.; Niemeijer, M.S.; Duine, H.J.; Luttik, M.A.H.; Boer, V.M.; Kotter, P.; Daran, J.M.G.; van Maris, A.J.A.; Pronk, J.T. Alternative reactions at the interface of glycolysis and citric acid cycle in Saccharomyces cerevisiae. Fems. Yeast Res. 2016, 16, fow017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimoto, A.; Ito, K.; Narushima, N.; Miyamoto, T. Identification of lactic acid bacteria and yeasts, and characterization of food components of sourdoughs used in Japanese bakeries. J. Biosci. Bioeng. 2019, 127, 575–581. [Google Scholar] [CrossRef]
- Boyaci Gunduz, C.P.; Gaglio, R.; Franciosi, E.; Settanni, L.; Erten, H. Molecular analysis of the dominant lactic acid bacteria of chickpea liquid starters and doughs and propagation of chickpea sourdoughs with selected Weissella confusa. Food Microbiol. 2020, 91, 103490. [Google Scholar] [CrossRef] [PubMed]
- Abedfar, A.; Hosseininezhad, M.; Corsetti, A. Effect of wheat bran sourdough with exopolysaccharide producing Lactobacillus plantarum (NR_104573.1) on quality of pan bread during shelf life. LWT 2019, 111, 158–166. [Google Scholar] [CrossRef]
- Mihhalevski, A.; Sarand, I.; Viiard, E.; Salumets, A.; Paalme, T. Growth characterization of individual rye sourdough bacteria by isothermal microcalorimetry. J. Appl. Microbiol. 2011, 110, 529–540. [Google Scholar] [CrossRef]
- Teleky, B.E.; M artău, A.G.; Ranga, F.; Chețan, F.; Vodnar, D.C. Exploitation of lactic acid bacteria and baker’s yeast as single or multiple starter cultures of wheat flour dough enriched with soy flour. Biomolecules 2020, 10, 778. [Google Scholar] [CrossRef]
- Thiele, C.; Gänzle, M.G.; Vogel, R.F. Contribution of sourdough Lactobacilli, yeast, and cereal enzymes to the generation of amino acids in dough relevant for bread flavor. Cereal. Chem. 2002, 79, 45–51. [Google Scholar] [CrossRef]
- Sieuwerts, S.; Bron, P.A.; Smid, E.J. Mutually stimulating interactions between lactic acid bacteria and Saccharomyces cerevisiae in sourdough fermentation. LWT 2018, 90, 201–206. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, Y.; Tang, K.X.; Hu, Y.; Xu, X.M.; Ganzle, M.G. Effect of mixed cultures of yeast and Lactobacilli on the quality of wheat sourdough bread. Front. Microbiol. 2019, 10, 2113. [Google Scholar] [CrossRef]
- Dymond, J.S. Saccharomyces cerevisiae growth media. Methods Enzymol. 2013, 533, 191–204. [Google Scholar] [CrossRef]
- Hayek, S.A.; Gyawali, R.; Aljaloud, S.O.; Krastanov, A.; Ibrahim, S.A. Cultivation media for lactic acid bacteria used in dairy products. J. Dairy Res. 2019, 86, 490–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantzourani, I.; Plessas, S.; Odatzidou, M.; Alexopoulos, A.; Galanis, A.; Bezirtzoglou, E.; Bekatorou, A. Effect of a novel Lactobacillus paracasei starter on sourdough bread quality. Food Chem. 2019, 271, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Ravi, R.K.; Walton, K.; Khosroheidari, M. MiSeq: A next generation sequencing platform for genomic analysis. Methods Mol. Biol. 2018, 1706, 223–232. [Google Scholar] [CrossRef]
- Wang, X.; Li, X.; Deng, X.; Han, H.; Shi, W.; Li, Y. A protein extraction method compatible with proteomic analysis for the euhalophyte Salicornia europaea. Electrophoresis 2007, 28, 3976–3987. [Google Scholar] [CrossRef]
- Cortés-Ríos, J.; Zárate, A.M.; Figueroa, J.D.; Medina, J.; Fuentes-Lemus, E.; Rodríguez-Fernández, M.; Aliaga, M.; López-Alarcón, C. Protein quantification by bicinchoninic acid (BCA) assay follows complex kinetics and can be performed at short incubation times. Anal. Biochem. 2020, 608, 113904. [Google Scholar] [CrossRef]
- Vrancken, G.; De Vuyst, L.; Rimaux, T.; Allemeersch, J.; Weckx, S. Adaptation of Lactobacillus plantarum IMDO 130201, a wheat sourdough isolate, to growth in wheat sourdough simulation medium at different pH values through differential gene expression. Appl. Environ. Microb. 2011, 77, 3406–3412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teleky, B.E.; Martău, G.A.; Vodnar, D.C. Physicochemical effects of Lactobacillus plantarum and Lactobacillus casei cocultures on soy–wheat flour dough fermentation. Foods 2020, 9, 1894. [Google Scholar] [CrossRef]
- Penido, F.C.L.; de Oliveira Goulart, C.; Galvão, Y.C.F.; Teixeira, C.V.; de Oliveira, R.B.P.; Borelli, B.M.; Guimarães, G.M.; Neumann, E.; Sande, D.; de Araújo, R.L.B.; et al. Antagonistic lactic acid bacteria in association with Saccharomyces cerevisiae as starter cultures for standardization of sour cassava starch production. J. Food Sci. Technol. 2019, 56, 3969–3979. [Google Scholar] [CrossRef]
- García Vilanova, M.; Díez, C.; Quirino, B.; Iñaki Álava, J. Microbiota distribution in sourdough: Influence of High sucrose resistant strains. Int. J. Gastron. Food Sci. 2015, 2, 98–102. [Google Scholar] [CrossRef] [Green Version]
- Richardson, G.; Langton, M.; Bark, A.; Hermansson, A.M. Wheat starch gelatinization—the effects of sucrose, emulsifier and the physical state of the emulsifier. Starke 2003, 55, 150–161. [Google Scholar] [CrossRef]
- Kang, M.J.; Bae, I.Y.; Lee, H.G. Rice noodle enriched with okara: Cooking property, texture, and in vitro starch digestibility. Food Biosci. 2018, 22, 178–183. [Google Scholar] [CrossRef]
- Meng, J.; Zhao, W. Properties and applications of trehalose in new food development. Food Sci. 2005, 26, 281–283. [Google Scholar] [CrossRef]
- Zhang, X.R.; Zhang, Y.X.; Li, H. Regulation of trehalose, a typical stress protectant, on central metabolisms, cell growth and division of Saccharomyces cerevisiae CEN.PK113-7D. Food Microbiol. 2020, 89, 103459. [Google Scholar] [CrossRef]
- Nam, Y.W.; Nishimoto, M.; Arakawa, T.; Kitaoka, M.; Pushinobu, S. Structural basis for broad substrate specificity of UDP-glucose 4-epimerase in the human milk oligosaccharide catabolic pathway of Bifidobacterium longum. Sci. Rep. 2019, 9, 11081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Cai, W.T.; Liu, S.L.; Zhou, C.H.; Cao, M.Y.; Yin, H.W.; Sun, D.X.; Zhang, S.L.; Loor, J.J. Association of UDP-galactose-4-epimerase with milk protein concentration in the Chinese Holstein population. Asian-Australas. J. Anim. Sci. 2020, 33, 1725–1731. [Google Scholar] [CrossRef]
- Zhang, S.S.; Xu, Z.S.; Qin, L.H.; Kong, J. Low-sugar yogurt making by the co-cultivation of Lactobacillus plantarum WCFS1 with yogurt starter cultures. J. Dairy Sci. 2020, 103, 3045–3054. [Google Scholar] [CrossRef]
- Heinisch, J.J.; Boles, E.; Timpel, C. A yeast phosphofructokinase insensitive to the allosteric activator fructose 2,6-bisphosphate -Glycolysis metabolic regulation allosteric control. J. Biol. Chem. 1996, 271, 15928–15933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, S.; Ali, S.; Mumtaz, S.; Shakir, H.A.; Ahmad, F.; Tahir, H.M.; Ulhaq, M.; Khan, M.A.; Zahid, M.T. Dose and duration-dependent toxicological evaluation of lead acetate in chicks. Environ. Sci. Pollut. R. 2020, 27, 15149–15164. [Google Scholar] [CrossRef] [PubMed]
- Heitmann, M.; Zannini, E.; Arendt, E. Impact of Saccharomyces cerevisiae metabolites produced during fermentation on bread quality parameters: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1152–1164. [Google Scholar] [CrossRef]
- Corsetti, A.; Settanni, L. Lactobacilli in sourdough fermentation. Food Res. Int. 2007, 40, 539–558. [Google Scholar] [CrossRef]
- Salim-ur-Rehman; Paterson, A.; Piggott, J.R. Flavour in sourdough breads: A review. Trends Food Sci. Tech. 2006, 17, 557–566. [Google Scholar] [CrossRef]
- Su, X.Q.; Wu, F.F.; Zhang, Y.Q.; Yang, N.; Chen, F.; Jina, Z.Y.; Xu, X.M. Effect of organic acids on bread quality improvement. Food Chem. 2019, 278, 267–275. [Google Scholar] [CrossRef]
- Kieliszek, M.; Pobiega, K.; Piwowarek, K.; Kot, A.M. Characteristics of the proteolytic enzymes produced by lactic acid bacteria. Molecules 2021, 26, 1858. [Google Scholar] [CrossRef]
- Waśko, A.; Kieliszek, M.; Targoński, Z. Purification and characterization of a proteinase from the probiotic Lactobacillus rhamnosus OXY. Prep. Biochem. Biotechnol. 2012, 42, 476–488. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, N.; Ganzle, M.G.; Vogel, R.F. Influence of peptide supply and cosubstrates on phenylalanine metabolism of Lactobacillus sanfranciscensis DSM20451(T) and Lactobacillus plantarum TMW1.468. J. Agric. Food Chem. 2006, 54, 3832–3839. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.; Zuniga, M. Amino acid catabolic pathways of lactic acid bacteria. Crit. Rev. Microbiol. 2006, 32, 155–183. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.; Rodriguez, A.; Fulco, M.; Soteras, T.; Mozgovoj, M.; Cap, M. Effects of lactic, malic and fumaric acids on Salmonella spp. counts and on chicken meat quality and sensory characteristics. J. Food Sci. Technol. 2020, 58, 3817–3824. [Google Scholar] [CrossRef] [PubMed]
- Mocibob, M.; Weygand-Durasevic, I. The proximal region of a noncatalytic eukaryotic seryl-tRNA synthetase extension is required for protein stability in vitro and in vivo. Arch. Biochem. Biophys. 2008, 470, 129–138. [Google Scholar] [CrossRef]
- Majid, M.; Khan, M.S.; Mustafa, G.; Salam, A. Structural and functional analysis of RPL16 a large ribosomal subunit protein of plastid translational machinery in plants. Int. J. Agric. Biol. 2019, 21, 223–228. [Google Scholar] [CrossRef]
- Ting, Y.H.; Lu, T.J.; Johnson, A.W.; Shie, J.T.; Chen, B.R.; Suresh, S.S.; Lo, K.Y. Bcp1 is the nuclear chaperone of Rpl23 in Saccharomyces cerevisiae. J. Biol. Chem. 2017, 292, 585–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciplys, E.; Zitkus, E.; Gold, L.I.; Daubriac, J.; Pavlides, S.C.; Hojrup, P.; Houen, G.; Wang, W.A.; Michalak, M.; Slibinskas, R. High-level secretion of native recombinant human calreticulin in yeast. Microb. Cell Factories 2015, 14, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession | Protein | Sx3 vs. Control | Sq7 vs. Control | Mix vs. Sx3 | Mix vs. Sq7 |
|---|---|---|---|---|---|
| Q88XY4 | Ribosomal Protein LW | up | - | - | - |
| Q88XY3 | Ribosomal Protein L2 | up | - | - | - |
| Q88XY2 | Ribosomal Protein S19 | up | up | - | - |
| A0A5R1Q4J0 | Ribosomal Protein L22 | up | - | - | - |
| Q88XY9 | Ribosomal Protein L16 | - | - | up | - |
| P38701 | Ribosomal Protein S20 | up | up | - | - |
| Q01855 | Ribosomal Protein S15 | - | up | - | - |
| Q88XX6 | Ribosomal Protein L14 | up | up | - | - |
| Q88XY4 | Ribosomal Protein L5 | - | - | up | - |
| A0A151G4G6 | Ribosomal Protein S8 | up | - | - | - |
| A0A199QKN2 | Ribosomal Protein L6 | up | - | - | - |
| Q88XW8 | Ribosomal Protein L30 | up | - | - | - |
| W5BFB7 | Ribosomal Protein S11 | up | - | - | - |
| W5E6W1 | Ribosomal Protein L23 | - | - | up | up |
| P41057 | Ribosomal Protein S29 | - | up | - | - |
| W5FEV1 | Ribosomal Protein L7 | up | - | - | - |
| T5JQX3 | Ribosomal Protein S13 | up | - | - | - |
| Q88XW1 | Ribosomal Protein SK | up | - | - | - |
| A0A199QHM8 | Ribosomal Protein S4 | up | - | - | - |
| A0A5R9DK27 | Ribosomal Protein L17 | up | - | - | - |
| A0A1W6NPG2 | Ribosomal Protein L13 | up | - | up | - |
| Q88XU7 | Ribosomal Protein S9 | up | up | - | - |
| P39516 | Ribosomal Protein S14 | - | up | - | - |
| B3LM31 | Ribosomal Protein S16 | - | up | - | - |
| Q88XY9 | Ribosomal Protein S7 | up | - | - | - |
| Q88YW7 | Ribosomal Protein L7 | up | up | - | - |
| Q88YW8 | Ribosomal Protein L10 | up | - | - | - |
| T5JY27 | Ribosomal Protein L11 | up | up | up | - |
| P0CX53 | Ribosomal Protein L12 | - | up | - | - |
| A0A484HZQ2 | Ribosomal Protein S2 | up | - | - | - |
| A0A165WB37 | Ribosomal Protein L31 | up | - | - | - |
| A0A2S3U801 | Ribosomal Protein L33 | up | - | - | - |
| Q88WN3 | Ribosomal Protein L27 | up | - | - | - |
| A0A165VYQ7 | Ribosomal Protein S1 | up | - | - | |
| A0A3B6EL93 | Ribosomal Protein L28 | up | - | up | - |
| A0A3B6AUJ9 | Ribosomal Protein S3 | down | up | - | - |
| E2F3W4 | Ribosomal Protein S15A | down | up | - | - |
| A0A3B6TBF6 | Ribosomal Protein L9 | down | up | down | down |
| A0A2X0S8T6 | Ribosomal Protein L14 | down | - | - | - |
| A0A3B5ZQF2 | Ribosomal Protein L18 | down | - | - | - |
| A0A3B6GLN8 | Ribosomal Protein S5 | down | up | - | - |
| A0A3B6SID4 | Ribosomal Protein L7A | down | down | - | - |
| Q5I7K5 | Ribosomal Protein LP1 | down | up | down | - |
| Q5I7L3 | Ribosomal Protein L10A | down | - | down | - |
| A0A1W5RML9 | Ribosomal Protein SA | down | - | - | - |
| W5G990 | Ribosomal Protein L31 | down | up | - | - |
| P07280 | Ribosomal Protein S19 | down | up | down | up |
| W5EP45 | Ribosomal Protein S25 | down | - | - | - |
| W5E8X2 | Ribosomal Protein S28 | down | - | down | - |
| W5FEZ3 | Ribosomal Protein S12 | - | - | down | - |
| Q5I7K2 | Ribosomal Protein S7 | down | up | - | - |
| A0A3B6ISR9 | Ribosomal Protein S21 | down | down | - | down |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, G.; Qi, Q.; Sadiq, F.A.; Wang, W.; He, X.; Wang, W. Proteomic Analysis Explores Interactions between Lactiplantibacillus plantarum and Saccharomyces cerevisiae during Sourdough Fermentation. Microorganisms 2021, 9, 2353. https://doi.org/10.3390/microorganisms9112353
Zhang G, Qi Q, Sadiq FA, Wang W, He X, Wang W. Proteomic Analysis Explores Interactions between Lactiplantibacillus plantarum and Saccharomyces cerevisiae during Sourdough Fermentation. Microorganisms. 2021; 9(11):2353. https://doi.org/10.3390/microorganisms9112353
Chicago/Turabian StyleZhang, Guohua, Qianhui Qi, Faizan Ahmed Sadiq, Wei Wang, Xiaxia He, and Wei Wang. 2021. "Proteomic Analysis Explores Interactions between Lactiplantibacillus plantarum and Saccharomyces cerevisiae during Sourdough Fermentation" Microorganisms 9, no. 11: 2353. https://doi.org/10.3390/microorganisms9112353
APA StyleZhang, G., Qi, Q., Sadiq, F. A., Wang, W., He, X., & Wang, W. (2021). Proteomic Analysis Explores Interactions between Lactiplantibacillus plantarum and Saccharomyces cerevisiae during Sourdough Fermentation. Microorganisms, 9(11), 2353. https://doi.org/10.3390/microorganisms9112353