
Neutrophils in Streptococcus suis Infection: From Host Defense to Pathology



Abstract
:1. Introduction
2. General Analysis of Published Literature
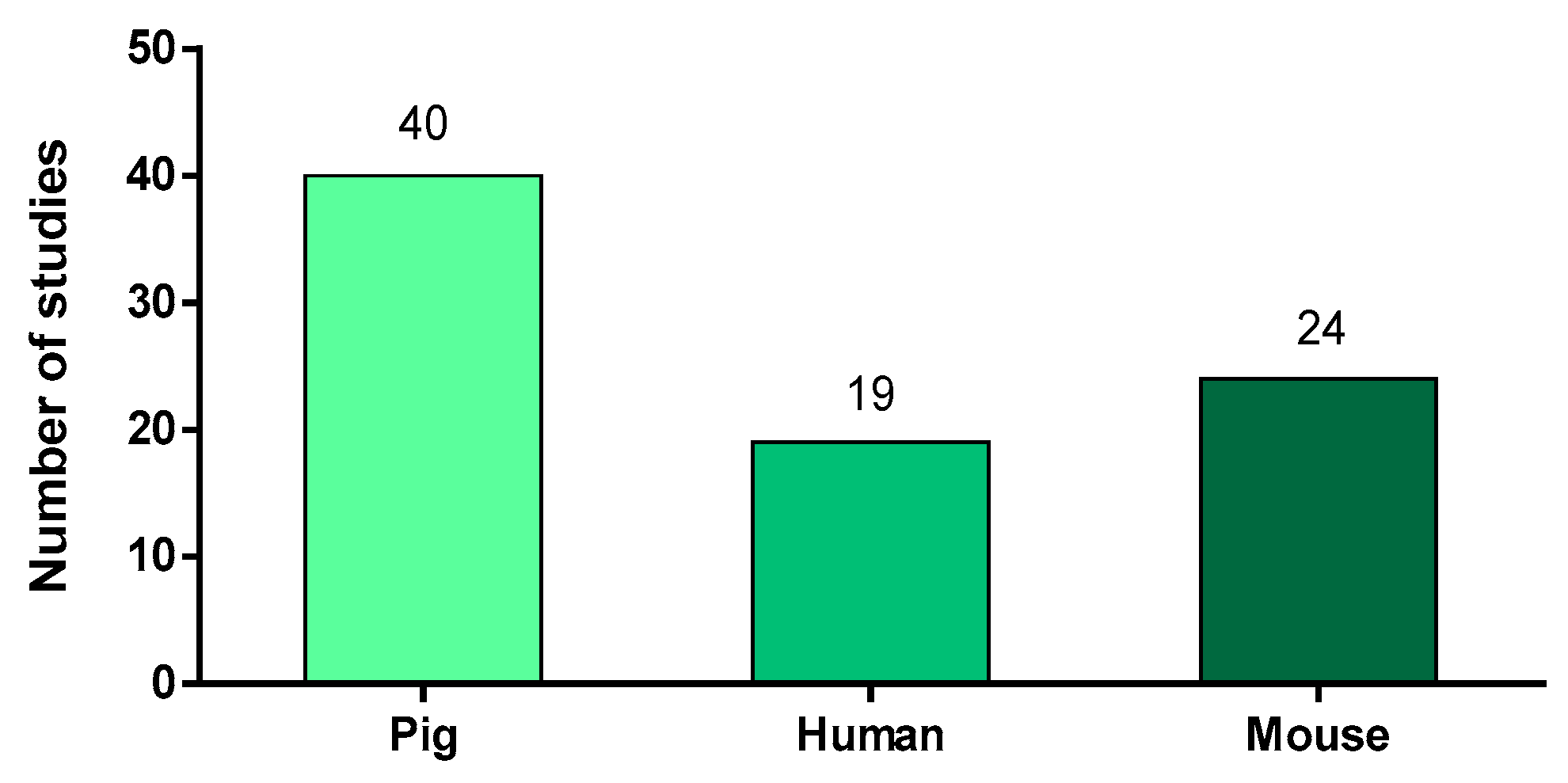
2.1. Animal Models or Species-Origin of the Neutrophils
2.2. S. suis Serotypes and Sequence Types
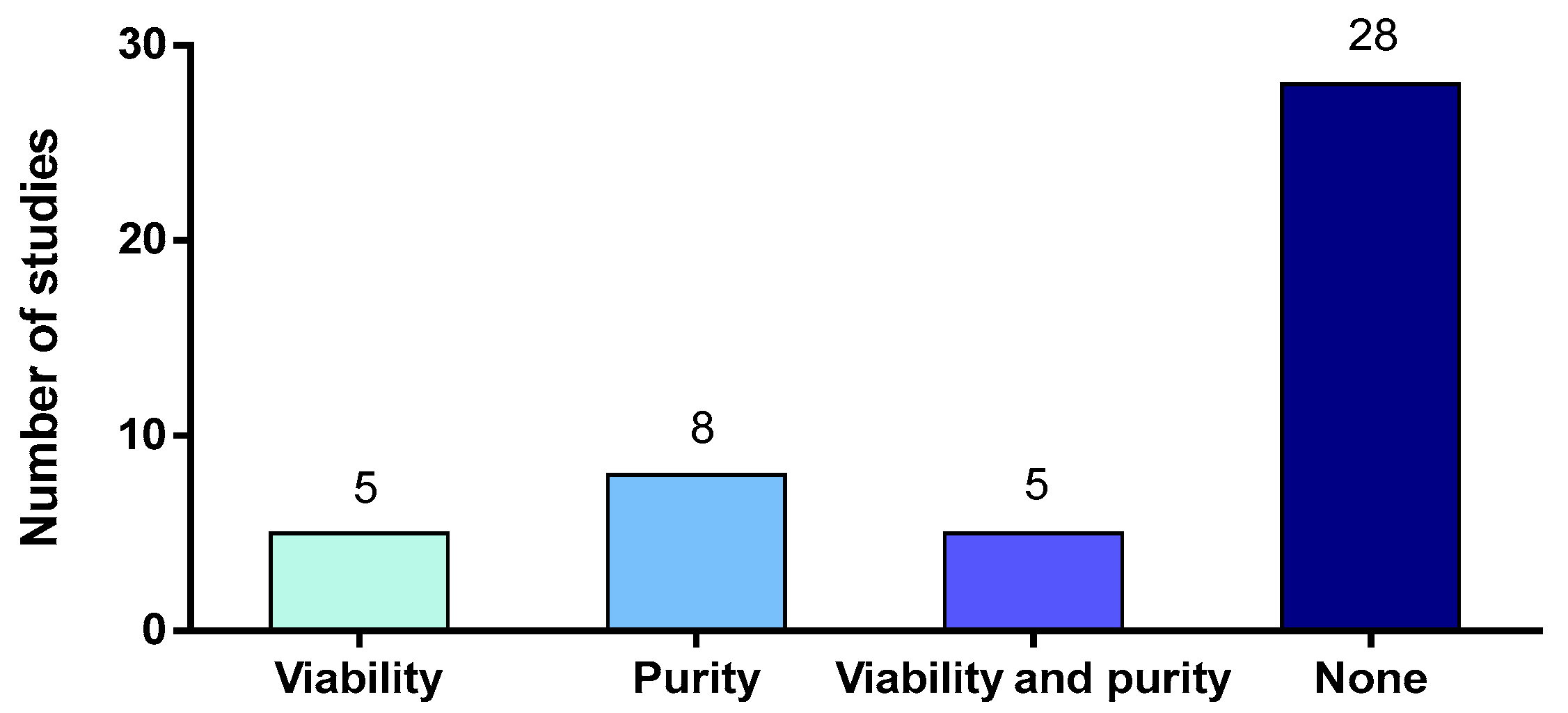
2.3. Purity and Viability of Neutrophils
2.3.1. Isolation Methods and Influence on the Purity of Neutrophils
2.3.2. Neutrophil Viability
3. In Vitro Functionality
3.1. Killing of S. suis by Neutrophils: Still a Controversy
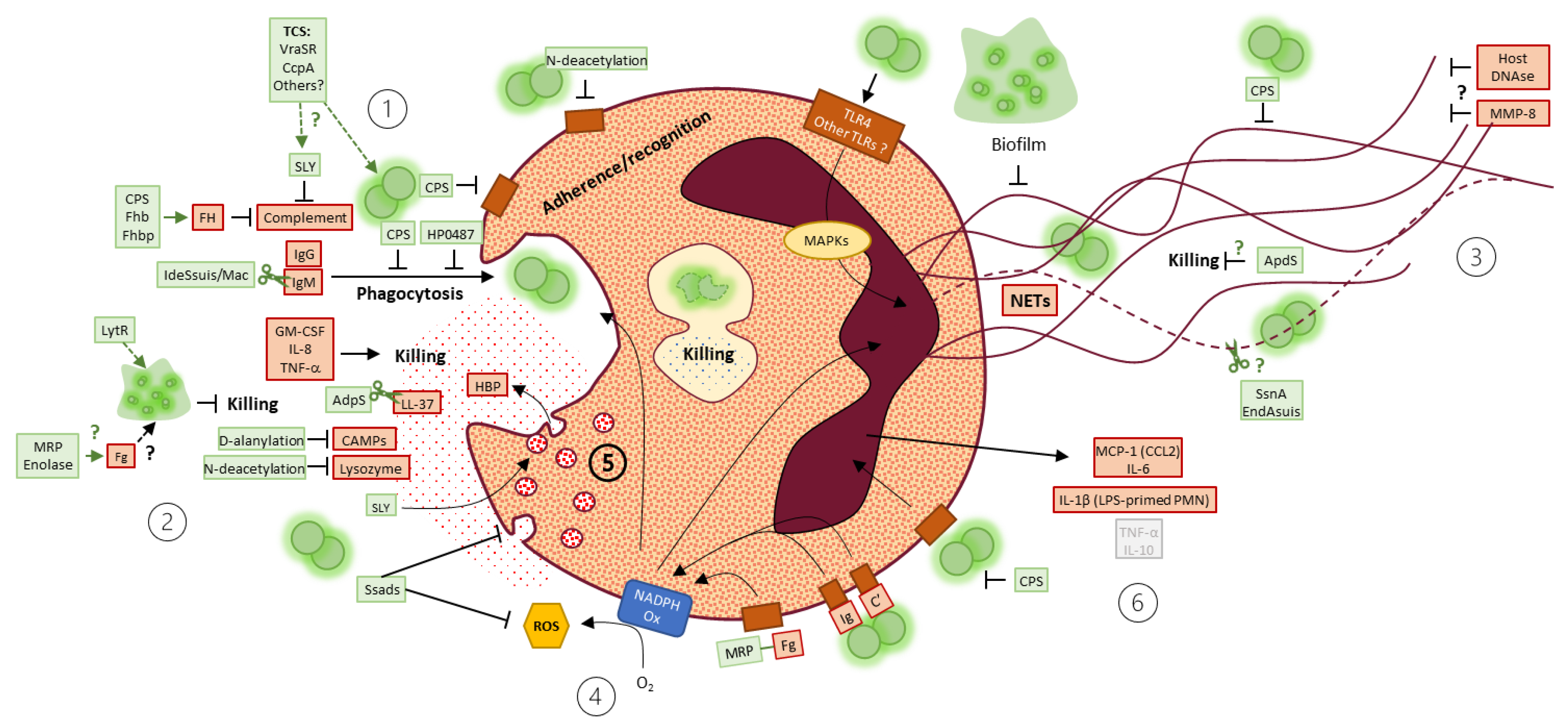
3.2. Phagocytosis: Neutrophils Poorly Phagocytize S. suis
3.3. Host Factors Facilitating S. suis Phagocytosis and/or Killing by Neutrophils
3.4. S. suis Strategies to Resist Neutrophil-Mediated Phagocytosis and/or Killing
3.5. NETs
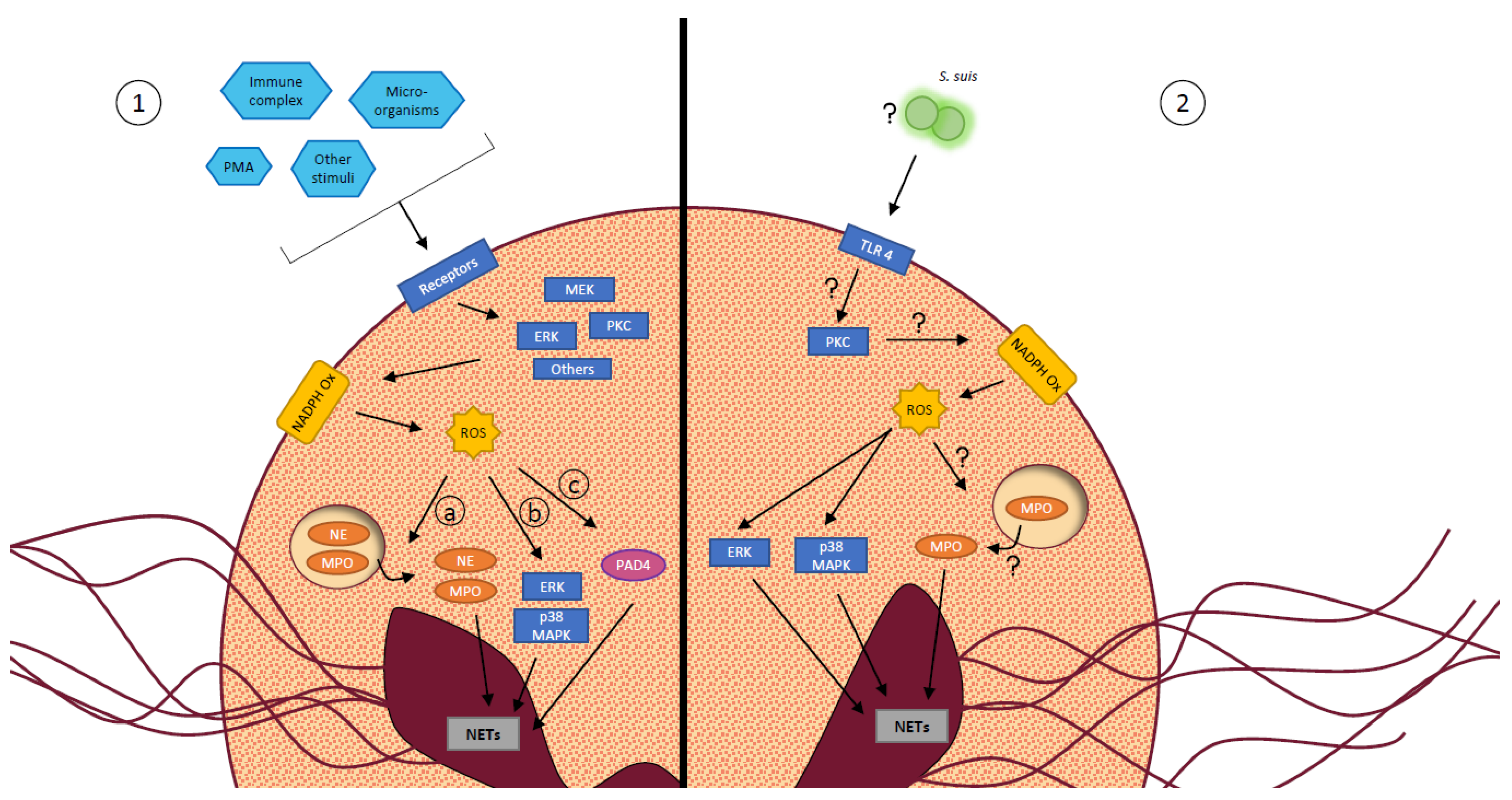
3.5.1. S. suis Induces NET Production by Neutrophils
3.5.2. S. suis Killing by NETs
3.5.3. S. suis Defense Mechanisms against NETs
3.6. ROS
3.7. Degranulation
3.8. Cytokines
3.9. Transmigration of Neutrophils
4. In Vivo Studies
4.1. S. suis-Induced Lesions Contain High Infiltration of Neutrophils
4.2. Recruitment and Role of Neutrophils during S. suis Systemic Infection
4.3. Recruitment and Role of Neutrophils in Central Nervous System Disease Caused by S. suis
4.4. Immune Regulation of Neutrophil in Response to S. suis Infection In Vivo
4.5. Bacterial Mechanisms against Neutrophils In Vivo
5. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Gottschalk, M.; Segura, M. Streptococcosis. In Diseases of Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2019; pp. 934–954. [Google Scholar]
- Gottschalk, M.; Xu, J.; Calzas, C.; Segura, M. Streptococcus suis: A new emerging or an old neglected zoonotic pathogen? Future Microbiol. 2010, 5, 371–391. [Google Scholar] [CrossRef] [PubMed]
- Wertheim, H.F.; Nghia, H.D.; Taylor, W.; Schultsz, C. Streptococcus suis: An emerging human pathogen. Clin. Infect. Dis. 2009, 48, 617–625. [Google Scholar] [CrossRef] [Green Version]
- Segura, M.; Fittipaldi, N.; Calzas, C.; Gottschalk, M. Critical Streptococcus suis virulence factors: Are they all really critical? Trends Microbiol. 2017, 25, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microbes Infect. 2014, 3, e45. [Google Scholar] [CrossRef] [PubMed]
- Ferrando, M.L.; Schultsz, C. A hypothetical model of host-pathogen interaction of Streptococcus suis in the gastro-intestinal tract. Gut Microbes 2016, 7, 154–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominguez-Punaro, M.C.; Segura, M.; Plante, M.M.; Lacouture, S.; Rivest, S.; Gottschalk, M. Streptococcus suis serotype 2, an important swine and human pathogen, induces strong systemic and cerebral inflammatory responses in a mouse model of infection. J. Immunol. 2007, 179, 1842–1854. [Google Scholar] [CrossRef] [Green Version]
- Reams, R.Y.; Glickman, L.T.; Harrington, D.D.; Thacker, H.L.; Bowersock, T.L. Streptococcus suis infection in swine: A retrospective study of 256 cases. Part II. Clinical signs, gross and microscopic lesions, and coexisting microorganisms. J. Vet. Diagn. Investig. 1994, 6, 326–334. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, D.; Middleton, D.M.; Chirino-Trejo, J.M. Lesions caused by natural infection with Streptococcus suis type 9 in weaned pigs. J. Vet. Diagn. Investig. 1994, 6, 335–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanford, S.E. Gross and histopathological findings in unusual lesions caused by Streptococcus suis in pigs. I. Cardiac lesions. Can. J. Vet. Res. 1987, 51, 481–485. [Google Scholar] [PubMed]
- Sanford, S.E. Gross and histopathological findings in unusual lesions caused by Streptococcus suis in pigs. II. Central nervous system lesions. Can. J. Vet. Res. 1987, 51, 486–489. [Google Scholar] [PubMed]
- Mayadas, T.N.; Cullere, X.; Lowell, C.A. The multifaceted functions of neutrophils. Annu. Rev. Pathol. 2014, 9, 181–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef]
- Migdal, C.; Serres, M. Reactive oxygen species and oxidative stress. Med. Sci. 2011, 27, 405–412. [Google Scholar] [CrossRef]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Tecchio, C.; Micheletti, A.; Cassatella, M.A. Neutrophil-derived cytokines: Facts beyond expression. Front. Immunol. 2014, 5, 508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordon, J.; Aliberti, S.; Fernandez-Botran, R.; Uriarte, S.M.; Rane, M.J.; Duvvuri, P.; Peyrani, P.; Morlacchi, L.C.; Blasi, F.; Ramirez, J.A. Understanding the roles of cytokines and neutrophil activity and neutrophil apoptosis in the protective versus deleterious inflammatory response in pneumonia. Int. J. Infect. Dis. 2013, 17, e76–e83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonego, F.; Castanheira, F.V.; Ferreira, R.G.; Kanashiro, A.; Leite, C.A.; Nascimento, D.C.; Colon, D.F.; Borges Vde, F.; Alves-Filho, J.C.; Cunha, F.Q. Paradoxical roles of the neutrophil in sepsis: Protective and deleterious. Front. Immunol. 2016, 7, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corsaut, L.; Misener, M.; Canning, P.; Beauchamp, G.; Gottschalk, M.; Segura, M. Field study on the immunological response and protective effect of a licensed autogenous vaccine to control Streptococcus suis infections in post-weaned piglets. Vaccines 2020, 8, 384. [Google Scholar] [CrossRef] [PubMed]
- Baums, C.G.; Bruggemann, C.; Kock, C.; Beineke, A.; Waldmann, K.H.; Valentin-Weigand, P. Immunogenicity of an autogenous Streptococcus suis bacterin in preparturient sows and their piglets in relation to protection after weaning. Clin. Vaccine Immunol. 2010, 17, 1589–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapointe, L.; D’Allaire, S.; Lebrun, A.; Lacouture, S.; Gottschalk, M. Antibody response to an autogenous vaccine and serologic profile for Streptococcus suis capsular type 1/2. Can. J. Vet. Res. 2002, 66, 8–14. [Google Scholar]
- Gottschalk, M.; Segura, M.; Xu, J. Streptococcus suis infections in humans: The Chinese experience and the situation in North America. Anim. Health Res. Rev. 2007, 8, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Chevaleyre, C.; Riou, M.; Brea, D.; Vandebrouck, C.; Barc, C.; Pezant, J.; Melo, S.; Olivier, M.; Delaunay, R.; Boulesteix, O.; et al. The pig: A relevant model for evaluating the neutrophil serine protease activities during acute Pseudomonas aeruginosa lung infection. PLoS ONE 2016, 11, e0168577. [Google Scholar] [CrossRef] [PubMed]
- Sipos, W.; Duvigneau, C.J.; Hartl, R.T.; Schwendenwein, I. Exploratory reference intervals on hematology and cellular immune system of multiparous Large White sows. Vet. Immunol. Immunopathol. 2011, 141, 307–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semiadi, G.; Nugraha, R.T.P. Some notes on biological aspects of captive Javan Warty Pig (Sus verrucosus). Biodivers. J. 2009, 10, 124–128. [Google Scholar] [CrossRef]
- Ježek, J.; Starič, J.; Nemec, M.; Plut, J.; Oven, I.G.; Klinkon, M.; Štukelj, M. The influence of age, farm, and physiological status on pig hematological profiles. J. Swine Health Prod. 2018, 26, 72–78. [Google Scholar]
- Mestas, J.; Hughes, C.C. Of mice and not men: Differences between mouse and human immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brea, D.; Meurens, F.; Dubois, A.V.; Gaillard, J.; Chevaleyre, C.; Jourdan, M.L.; Winter, N.; Arbeille, B.; Si-Tahar, M.; Gauthier, F.; et al. The pig as a model for investigating the role of neutrophil serine proteases in human inflammatory lung diseases. Biochem. J. 2012, 447, 363–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Buhr, N.; Neumann, A.; Jerjomiceva, N.; von Kockritz-Blickwede, M.; Baums, C.G. Streptococcus suis DNase SsnA contributes to degradation of neutrophil extracellular traps (NETs) and evasion of NET-mediated antimicrobial activity. Microbiology 2014, 160, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Roberts, R.L.; Hatori, N.; Drury, J.K.; Stiehm, E.R. Purification and properties of porcine polymorphonuclear cells. J. Immunol. Methods 1987, 103, 27–32. [Google Scholar] [CrossRef]
- Wang, F.I.; Yang, J.W.; Hung, S.Y.; Pan, I.J. In vitro migratory responses of swine neutrophils to Actinobacillus pleuropneumoniae. Exp. Anim. 2001, 50, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, M.P.; Stahl, G.L.; Longhurst, J.C. In vivo and in vitro assessment of porcine neutrophil activation responses to chemoattractants: Flow cytometric evidence for the selective absence of formyl peptide receptors. J. Leukoc. Biol. 1990, 47, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Eisenhauer, P.B.; Lehrer, R.I. Mouse neutrophils lack defensins. Infect. Immun. 1992, 60, 3446–3447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sang, Y.; Blecha, F. Porcine host defense peptides: Expanding repertoire and functions. Dev. Comp. Immunol. 2009, 33, 334–343. [Google Scholar] [CrossRef]
- Matute-Bello, G.; Downey, G.; Moore, B.B.; Groshong, S.D.; Matthay, M.A.; Slutsky, A.S.; Kuebler, W.M.; Acute Lung Injury in Animals Study Group. An official American Thoracic Society workshop report: Features and measurements of experimental acute lung injury in animals. Am. J. Respir. Cell Mol. Biol. 2011, 44, 725–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prufer, T.L.; Rohde, J.; Verspohl, J.; Rohde, M.; de Greeff, A.; Willenborg, J.; Valentin-Weigand, P. Molecular typing of Streptococcus suis strains isolated from diseased and healthy pigs between 1996–2016. PLoS ONE 2019, 14, e0210801. [Google Scholar] [CrossRef] [PubMed]
- Wisselink, H.J.; Smith, H.E.; Stockhofe-Zurwieden, N.; Peperkamp, K.; Vecht, U. Distribution of capsular types and production of muramidase-released protein (MRP) and extracellular factor (EF) of Streptococcus suis strains isolated from diseased pigs in seven European countries. Vet. Microbiol. 2000, 74, 237–248. [Google Scholar] [CrossRef]
- Baums, C.G.; Kock, C.; Beineke, A.; Bennecke, K.; Goethe, R.; Schroder, C.; Waldmann, K.H.; Valentin-Weigand, P. Streptococcus suis bacterin and subunit vaccine immunogenicities and protective efficacies against serotypes 2 and 9. Clin. Vaccine Immunol. 2009, 16, 200–208. [Google Scholar] [CrossRef] [Green Version]
- Kock, C.; Beineke, A.; Seitz, M.; Ganter, M.; Waldmann, K.H.; Valentin-Weigand, P.; Baums, C.G. Intranasal immunization with a live Streptococcus suis isogenic ofs mutant elicited suilysin-neutralization titers but failed to induce opsonizing antibodies and protection. Vet. Immunol. Immunopathol. 2009, 132, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Beineke, A.; Bennecke, K.; Neis, C.; Schroder, C.; Waldmann, K.H.; Baumgartner, W.; Valentin-Weigand, P.; Baums, C.G. Comparative evaluation of virulence and pathology of Streptococcus suis serotypes 2 and 9 in experimentally infected growers. Vet. Microbiol. 2008, 128, 423–430. [Google Scholar] [CrossRef]
- Rungelrath, V.; Ohlmann, S.; Alber, G.; Schrodl, W.; von Kockritz-Blickwede, M.; de Buhr, N.; Martens, A.; Baums, C.G.; Schutze, N. Survival of Streptococcus suis in porcine blood is limited by the antibody- and complement-dependent oxidative burst response of granulocytes. Infect. Immun. 2020, 88, 300598–300619. [Google Scholar] [CrossRef] [PubMed]
- Auger, J.P.; Payen, S.; Roy, D.; Dumesnil, A.; Segura, M.; Gottschalk, M. Interactions of Streptococcus suis serotype 9 with host cells and role of the capsular polysaccharide: Comparison with serotypes 2 and 14. PLoS ONE 2019, 14, e0223864. [Google Scholar] [CrossRef] [PubMed]
- Chabot-Roy, G.; Willson, P.; Segura, M.; Lacouture, S.; Gottschalk, M. Phagocytosis and killing of Streptococcus suis by porcine neutrophils. Microb. Pathog. 2006, 41, 21–32. [Google Scholar] [CrossRef]
- Quessy, S.; Dubreuil, J.D.; Jacques, M.; Malouin, F.; Higgins, R. Increase of capsular material thickness following in vivo growth of virulent Streptococcus suis serotype 2 strains. FEMS Microbiol. Lett. 1994, 115, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Henningham, A.; Dohrmann, S.; Nizet, V.; Cole, J.N. Mechanisms of group A streptococcus resistance to reactive oxygen species. FEMS Microbiol. Rev. 2015, 39, 488–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auger, J.P.; Dolbec, D.; Roy, D.; Segura, M.; Gottschalk, M. Role of the Streptococcus suis serotype 2 capsular polysaccharide in the interactions with dendritic cells is strain-dependent but remains critical for virulence. PLoS ONE 2018, 13, e0200453. [Google Scholar] [CrossRef]
- Roy, D.; Auger, J.P.; Segura, M.; Fittipaldi, N.; Takamatsu, D.; Okura, M.; Gottschalk, M. Role of the capsular polysaccharide as a virulence factor for Streptococcus suis serotype 14. Can. J. Vet. Res. 2015, 79, 141–146. [Google Scholar] [PubMed]
- Auger, J.P.; Christodoulides, M.; Segura, M.; Xu, J.; Gottschalk, M. Interactions of Streptococcus suis serotype 2 with human meningeal cells and astrocytes. BMC Res. Notes 2015, 8, 607. [Google Scholar] [CrossRef] [Green Version]
- Auger, J.P.; Santinon, A.; Roy, D.; Mossman, K.; Xu, J.; Segura, M.; Gottschalk, M. Type I interferon induced by Streptococcus suis serotype 2 is strain-dependent and may be beneficial for host survival. Front. Immunol. 2017, 8, 1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auger, J.P.; Fittipaldi, N.; Benoit-Biancamano, M.O.; Segura, M.; Gottschalk, M. Virulence studies of different sequence types and geographical origins of Streptococcus suis serotype 2 in a mouse model of infection. Pathogens 2016, 5, 48. [Google Scholar] [CrossRef] [PubMed]
- Kubes, P. The enigmatic neutrophil: What we do not know. Cell Tissue Res. 2018, 371, 399–406. [Google Scholar] [CrossRef]
- Kuhns, D.B.; Priel, D.A.L.; Chu, J.; Zarember, K.A. Isolation and functional analysis of human neutrophils. Curr. Protoc. Immunol. 2015, 111, 7.23.1–7.23.16. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, M.C.; Fingerhut, L.; Alfonso-Castro, A.; Mergani, A.; Schwennen, C.; von Kockritz-Blickwede, M.; de Buhr, N. How long does a neutrophil live?—The effect of 24 h whole blood storage on neutrophil functions in pigs. Biomedicines 2020, 8, 278. [Google Scholar] [CrossRef]
- Thomas, H.B.; Moots, R.J.; Edwards, S.W.; Wright, H.L. Whose gene is it anyway? The effect of preparation purity on neutrophil transcriptome studies. PLoS ONE 2015, 10, e0138982. [Google Scholar] [CrossRef] [Green Version]
- Calzetti, F.; Tamassia, N.; Arruda-Silva, F.; Gasperini, S.; Cassatella, M.A. The importance of being “pure” neutrophils. J. Allergy Clin. Immunol. 2017, 139, 352–355. [Google Scholar] [CrossRef] [Green Version]
- Silvestre-Roig, C.; Fridlender, Z.G.; Glogauer, M.; Scapini, P. Neutrophil diversity in health and disease. Trends Immunol. 2019, 40, 565–583. [Google Scholar] [CrossRef]
- Quach, A.; Ferrante, A. The application of dextran sedimentation as an initial step in neutrophil purification promotes their stimulation, due to the presence of monocytes. J. Immunol. Res. 2017, 2017, 1254792. [Google Scholar] [CrossRef] [PubMed]
- Baltimore, R.S.; Kasper, D.L.; Baker, C.J.; Goroff, D.K. Antigenic specificity of opsonophagocytic antibodies in rabbit anti-sera to group B streptococci. J. Immunol. 1977, 118, 673–678. [Google Scholar]
- Swamydas, M.; Luo, Y.; Dorf, M.E.; Lionakis, M.S. Isolation of Mouse Neutrophils. Curr. Protoc. Immunol. 2015, 110, 3–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasenberg, M.; Kohler, A.; Bonifatius, S.; Borucki, K.; Riek-Burchardt, M.; Achilles, J.; Mann, L.; Baumgart, K.; Schraven, B.; Gunzer, M. Rapid immunomagnetic negative enrichment of neutrophil granulocytes from murine bone marrow for functional studies in vitro and in vivo. PLoS ONE 2011, 6, e17314. [Google Scholar] [CrossRef] [PubMed]
- Hohnstein, F.S.; Meurer, M.; de Buhr, N.; von Kockritz-Blickwede, M.; Baums, C.G.; Alber, G.; Schutze, N. Analysis of porcine pro- and anti-inflammatory cytokine induction by S. suis in vivo and in vitro. Pathogens 2020, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Weisse, C.; Dittmar, D.; Jakobczak, B.; Florian, V.; Schutze, N.; Alber, G.; Klose, K.; Michalik, S.; Valentin-Weigand, P.; Volker, U.; et al. Immunogenicity and protective efficacy of a Streptococcus suis vaccine composed of six conserved immunogens. Vet. Res. 2021, 52, 112. [Google Scholar] [CrossRef]
- Tamassia, N.; Bianchetto-Aguilera, F.; Arruda-Silva, F.; Gardiman, E.; Gasperini, S.; Calzetti, F.; Cassatella, M.A. Cytokine production by human neutrophils: Revisiting the “dark side of the moon”. Eur J. Clin. Investig. 2018, 48 (Suppl. S2). [Google Scholar] [CrossRef] [PubMed]
- Doster, R.S.; Rogers, L.M.; Gaddy, J.A.; Aronoff, D.M. Macrophage extracellular traps: A scoping review. J. Innate Immun. 2018, 10, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Goldmann, O.; Medina, E. The expanding world of extracellular traps: Not only neutrophils but much more. Front. Immunol. 2012, 3, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupre-Crochet, S.; Erard, M.; Nubetae, O. ROS production in phagocytes: Why, when, and where? J. Leukoc. Biol. 2013, 94, 657–670. [Google Scholar] [CrossRef] [PubMed]
- SenGupta, S.; Rane, M.J.; Uriarte, S.M.; Woolley, C.; Mitchell, T.C. Human neutrophils depend on extrinsic factors produced by monocytes for their survival response to TLR4 stimulation. Innate Immun. 2019, 25, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Yi, L.; Yu, N.; Wang, G.; Ma, Z.; Lin, H.; Fan, H. Streptococcus suis serotype 2 biofilms inhibit the formation of neutrophil extracellular traps. Front. Cell. Infect. Microbiol. 2017, 7, 86. [Google Scholar] [CrossRef] [Green Version]
- Ma, F.; Chang, X.; Wang, G.; Zhou, H.; Ma, Z.; Lin, H.; Fan, H. Streptococcus suis serotype 2 stimulates neutrophil extracellular traps formation via activation of p38 MAPK and ERK1/2. Front. Immunol. 2018, 9, 2854. [Google Scholar] [CrossRef]
- Zhao, J.; Pan, S.; Lin, L.; Fu, L.; Yang, C.; Xu, Z.; Wei, Y.; Jin, M.; Zhang, A. Streptococcus suis serotype 2 strains can induce the formation of neutrophil extracellular traps and evade trapping. FEMS Microbiol. Lett. 2015, 362, fnv022. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Xu, L.; Lv, W.; Han, L.; Xiang, Y.; Fu, L.; Jin, M.; Zhou, R.; Chen, H.; Zhang, A. An NLRP3 inflammasome-triggered cytokine storm contributes to streptococcal toxic shock-like syndrome (STSLS). PLoS Pathog. 2019, 15, e1007795. [Google Scholar] [CrossRef]
- Scapinello, S.; Brooks, A.S.; MacInnes, J.I.; Hammermueller, J.; Clark, M.E.; Caswell, J.L. Bactericidal activity of porcine neutrophil secretions. Vet. Immunol. Immunopathol. 2011, 139, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xie, W.; Wu, K.; Li, P.; Ren, Z.; Li, L.; Yuan, Y.; Zhang, C.; Zheng, Y.; Lv, Q.; et al. Suilysin stimulates the release of heparin binding protein from neutrophils and increases vascular permeability in mice. Front. Microbiol. 2016, 7, 1338. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhao, J.; Cai, C.; Tang, X.; Fu, L.; Zhang, A.; Han, L. A label-free quantitative proteomic analysis of mouse neutrophil extracellular trap formation induced by Streptococcus suis or phorbol myristate acetate (PMA). Front. Immunol. 2018, 9, 2615. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Li, N.; Jia, L.; Guo, W.; Jiang, H.; Liu, B.; Bao, C.; Liu, M.; Huang, J.; Lei, L. Ribosomal protein SA-positive neutrophil elicits stronger phagocytosis and neutrophil extracellular trap formation and subdues pro-inflammatory cytokine secretion against Streptococcus suis serotype 2 infection. Front. Immunol. 2020, 11, 585399. [Google Scholar] [CrossRef] [PubMed]
- Ni, C.; Gao, S.; Zheng, Y.; Liu, P.; Zhai, Y.; Huang, W.; Jiang, H.; Lv, Q.; Kong, D.; Jiang, Y. Annexin A1 attenuates neutrophil migration and IL-6 expression through Fpr2 in a mouse model of Streptococcus suis-induced meningitis. Infect. Immun. 2021, 89, e00680-20. [Google Scholar] [CrossRef] [PubMed]
- Leitch, A.E.; Lucas, C.D.; Marwick, J.A.; Duffin, R.; Haslett, C.; Rossi, A.G. Cyclin-dependent kinases 7 and 9 specifically regulate neutrophil transcription and their inhibition drives apoptosis to promote resolution of inflammation. Cell Death Differ. 2012, 19, 1950–1961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boxio, R.; Bossenmeyer-Pourie, C.; Steinckwich, N.; Dournon, C.; Nusse, O. Mouse bone marrow contains large numbers of functionally competent neutrophils. J. Leukoc. Biol. 2004, 75, 604–611. [Google Scholar] [CrossRef]
- Li, M.; Wang, C.; Feng, Y.; Pan, X.; Cheng, G.; Wang, J.; Ge, J.; Zheng, F.; Cao, M.; Dong, Y.; et al. SalK/SalR, a two-component signal transduction system, is essential for full virulence of highly invasive Streptococcus suis serotype 2. PLoS ONE 2008, 3, e2080. [Google Scholar] [CrossRef] [Green Version]
- Wibawan, I.W.; Lammler, C. Relation between encapsulation and various properties of Streptococcus suis. J. Vet. Med. B 1994, 41, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Benga, L.; Fulde, M.; Neis, C.; Goethe, R.; Valentin-Weigand, P. Polysaccharide capsule and suilysin contribute to extracellular survival of Streptococcus suis co-cultivated with primary porcine phagocytes. Vet. Microbiol. 2008, 132, 211–219. [Google Scholar] [CrossRef]
- Li, W.; Hu, X.; Liu, L.; Chen, H.; Zhou, R. Induction of protective immune response against Streptococcus suis serotype 2 infection by the surface antigen HP0245. FEMS Microbiol. Lett. 2011, 316, 115–122. [Google Scholar] [CrossRef] [PubMed]
- de Buhr, N.; Reuner, F.; Neumann, A.; Stump-Guthier, C.; Tenenbaum, T.; Schroten, H.; Ishikawa, H.; Muller, K.; Beineke, A.; Hennig-Pauka, I.; et al. Neutrophil extracellular trap formation in the Streptococcus suis-infected cerebrospinal fluid compartment. Cell. Microbiol. 2017, 19, e12649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, P.; Li, W.; Shi, G.; Li, H.; Yang, X.; Xia, Z.; Ren, Y.; Li, Z.; Chen, H.; Bei, W. The VraSR regulatory system contributes to virulence in Streptococcus suis via resistance to innate immune defenses. Virulence 2018, 9, 771–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fittipaldi, N.; Sekizaki, T.; Takamatsu, D.; de la Cruz Dominguez-Punaro, M.; Harel, J.; Bui, N.K.; Vollmer, W.; Gottschalk, M. Significant contribution of the pgdA gene to the virulence of Streptococcus suis. Mol. Microbiol. 2008, 70, 1120–1135. [Google Scholar] [CrossRef]
- Fittipaldi, N.; Sekizaki, T.; Takamatsu, D.; Harel, J.; de la Cruz Dominguez-Punaro, M.; Von Aulock, S.; Draing, C.; Marois, C.; Kobisch, M.; Gottschalk, M. D-alanylation of lipoteichoic acid contributes to the virulence of Streptococcus suis. Infect. Immun. 2008, 76, 3587–3594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Gascon, L.; Cardoso-Toset, F.; Tarradas, C.; Gomez-Laguna, J.; Maldonado, A.; Nielsen, J.; Olaya-Abril, A.; Rodriguez-Ortega, M.J.; Luque, I. Characterization of the immune response and evaluation of the protective capacity of rSsnA against Streptococcus suis infection in pigs. Comp. Immunol. Microbiol. Infect. Dis. 2016, 47, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Seele, J.; Beineke, A.; Hillermann, L.M.; Jaschok-Kentner, B.; von Pawel-Rammingen, U.; Valentin-Weigand, P.; Baums, C.G. The immunoglobulin M-degrading enzyme of Streptococcus suis, IdeSsuis, is involved in complement evasion. Vet. Res. 2015, 46, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, F.; Zan, Y.; Zhang, Y.; Zheng, N.; Yan, Q.; Zhang, W.; Zhang, H.; Jin, M.; Chen, F.; Zhang, X.; et al. The cysteine protease ApdS from Streptococcus suis promotes evasion of innate immune defenses by cleaving the antimicrobial peptide cathelicidin LL-37. J. Biol. Chem. 2019, 294, 17962–17977. [Google Scholar] [CrossRef]
- de Buhr, N.; Stehr, M.; Neumann, A.; Naim, H.Y.; Valentin-Weigand, P.; von Kockritz-Blickwede, M.; Baums, C.G. Identification of a novel DNase of Streptococcus suis (EndAsuis) important for neutrophil extracellular trap degradation during exponential growth. Microbiology 2015, 161, 838–850. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Chen, Y.; Li, Q.; Jiang, H.; Lv, Q.; Zheng, Y.; Han, X.; Kong, D.; Liu, P.; Jiang, Y. LytR plays a role in normal septum formation and contributes to full virulence in Streptococcus suis. Vet. Microbiol. 2021, 254, 109003. [Google Scholar] [CrossRef] [PubMed]
- Pian, Y.; Li, X.; Zheng, Y.; Wu, X.; Yuan, Y.; Jiang, Y. Binding of human fibrinogen to MRP enhances Streptococcus suis survival in host blood in a αXβ2 integrin-dependent manner. Sci. Rep. 2016, 6, 26966. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, P.; Gan, S.; Zhang, C.; Zheng, Y.; Jiang, Y.; Yuan, Y. Mechanisms of host-pathogen protein complex formation and bacterial immune evasion of Streptococcus suis protein Fhb. J. Biol. Chem. 2016, 291, 17122–17132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, X.; Ge, J.; Li, M.; Wu, B.; Wang, C.; Wang, J.; Feng, Y.; Yin, Z.; Zheng, F.; Cheng, G.; et al. The orphan response regulator CovR: A globally negative modulator of virulence in Streptococcus suis serotype 2. J. Bacteriol. 2009, 191, 2601–2612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pian, Y.; Wang, P.; Liu, P.; Zheng, Y.; Zhu, L.; Wang, H.; Xu, B.; Yuan, Y.; Jiang, Y. Proteomics identification of novel fibrinogen-binding proteins of Streptococcus suis contributing to antiphagocytosis. Front. Cell. Infect. Microbiol 2015, 5, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Pian, Y.; Li, X.; Liu, R.; Xie, W.; Zhang, C.; Zheng, Y.; Jiang, Y.; Yuan, Y. Streptococcus suis adenosine synthase functions as an effector in evasion of PMN-mediated innate immunity. J. Infect. Dis. 2014, 210, 35–45. [Google Scholar] [CrossRef]
- Pian, Y.; Gan, S.; Wang, S.; Guo, J.; Wang, P.; Zheng, Y.; Cai, X.; Jiang, Y.; Yuan, Y. Fhb, a novel factor H-binding surface protein, contributes to the antiphagocytic ability and virulence of Streptococcus suis. Infect. Immun. 2012, 80, 2402–2413. [Google Scholar] [CrossRef] [Green Version]
- Rosales, C.; Uribe-Querol, E. Phagocytosis: A fundamental process in immunity. BioMed Res. Int. 2017, 2017, 9042851. [Google Scholar] [CrossRef] [Green Version]
- Lun, S.; Willson, P.J. Expression of green fluorescent protein and its application in pathogenesis studies of serotype 2 Streptococcus suis. J. Microbiol. Methods 2004, 56, 401–412. [Google Scholar] [CrossRef]
- He, Z.; Pian, Y.; Ren, Z.; Bi, L.; Yuan, Y.; Zheng, Y.; Jiang, Y.; Wang, F. Increased production of suilysin contributes to invasive infection of the Streptococcus suis strain 05ZYH33. Mol. Med. Rep. 2014, 10, 2819–2826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fittipaldi, N.; Segura, M.; Grenier, D.; Gottschalk, M. Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol. 2012, 7, 259–279. [Google Scholar] [CrossRef] [PubMed]
- Hui, X.; Xu, Z.; Cao, L.; Liu, L.; Lin, X.; Yang, Y.; Sun, X.; Zhang, Q.; Jin, M. HP0487 contributes to the virulence of Streptococcus suis serotype 2 by mediating bacterial adhesion and anti-phagocytosis to neutrophils. Vet. Microbiol. 2021, 260, 109164. [Google Scholar] [CrossRef] [PubMed]
- Jog, N.R.; Rane, M.J.; Lominadze, G.; Luerman, G.C.; Ward, R.A.; McLeish, K.R. The actin cytoskeleton regulates exocytosis of all neutrophil granule subsets. Am. J. Physiol. Cell Physiol. 2007, 292, C1690–C1700. [Google Scholar] [CrossRef] [Green Version]
- Busque, P.; Higgins, R.; Senechal, S.; Marchand, R.; Quessy, S. Simultaneous flow cytometric measurement of Streptococcus suis phagocytosis by polymorphonuclear and mononuclear blood leukocytes. Vet. Microbiol. 1998, 63, 229–238. [Google Scholar] [CrossRef]
- Lecours, M.P.; Segura, M.; Lachance, C.; Mussa, T.; Surprenant, C.; Montoya, M.; Gottschalk, M. Characterization of porcine dendritic cell response to Streptococcus suis. Vet. Res. 2011, 42, 72. [Google Scholar] [CrossRef] [Green Version]
- Meijerink, M.; Ferrando, M.L.; Lammers, G.; Taverne, N.; Smith, H.E.; Wells, J.M. Immunomodulatory effects of Streptococcus suis capsule type on human dendritic cell responses, phagocytosis and intracellular survival. PLoS ONE 2012, 7, e35849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meurer, M.; Ohlmann, S.; Bonilla, M.C.; Valentin-Weigand, P.; Beineke, A.; Hennig-Pauka, I.; Schwerk, C.; Schroten, H.; Baums, C.G.; Kockritz-Blickwede, M.V.; et al. Role of bacterial and host DNases on host-pathogen interaction during Streptococcus suis meningitis. Int. J. Mol. Sci. 2020, 21, 5289. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Gottschalk, M.; Esgleas, M.; Lacouture, S.; Dubreuil, J.D.; Willson, P.; Harel, J. Immunization with recombinant Sao protein confers protection against Streptococcus suis infection. Clin. Vaccine Immunol. 2007, 14, 937–943. [Google Scholar] [CrossRef] [Green Version]
- Miralda, I.; Uriarte, S.M.; McLeish, K.R. Multiple phenotypic changes define neutrophil priming. Front. Cell. Infect. Microbiol. 2017, 7, 217. [Google Scholar] [CrossRef] [PubMed]
- Willenborg, J.; Fulde, M.; de Greeff, A.; Rohde, M.; Smith, H.E.; Valentin-Weigand, P.; Goethe, R. Role of glucose and CcpA in capsule expression and virulence of Streptococcus suis. Microbiology 2011, 157, 1823–1833. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Li, L.; Ge, H.; Meng, H.; Li, Y.; Bei, W.; Zhou, X. Role of two-component regulatory systems in the virulence of Streptococcus suis. Microbiol. Res. 2018, 214, 123–128. [Google Scholar] [CrossRef]
- Xu, J.; Fu, S.; Liu, M.; Xu, Q.; Bei, W.; Chen, H.; Tan, C. The two-component system NisK/NisR contributes to the virulence of Streptococcus suis serotype 2. Microbiol. Res. 2014, 169, 541–546. [Google Scholar] [CrossRef]
- Gottschalk, M.; Segura, M. The pathogenesis of the meningitis caused by Streptococcus suis: The unresolved questions. Vet. Microbiol. 2000, 76, 259–272. [Google Scholar] [CrossRef]
- Ragland, S.A.; Criss, A.K. From bacterial killing to immune modulation: Recent insights into the functions of lysozyme. PLoS Pathog. 2017, 13, e1006512. [Google Scholar] [CrossRef]
- Boneca, I.G.; Dussurget, O.; Cabanes, D.; Nahori, M.A.; Sousa, S.; Lecuit, M.; Psylinakis, E.; Bouriotis, V.; Hugot, J.P.; Giovannini, M.; et al. A critical role for peptidoglycan N-deacetylation in Listeria evasion from the host innate immune system. Proc. Natl. Acad. Sci. USA 2007, 104, 997–1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, D.; Grenier, D.; Segura, M.; Mathieu-Denoncourt, A.; Gottschalk, M. Recruitment of factor H to the Streptococcus suis cell surface is multifactorial. Pathogens 2016, 5, 47. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Wu, Z.; Zhang, S.; Tang, H.; Wang, F.; Lu, C. Mac protein is not an essential virulence factor for the virulent reference strain Streptococcus suis P1/7. Curr. Microbiol. 2017, 74, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Zawrotniak, M.; Rapala-Kozik, M. Neutrophil extracellular traps (NETs)—Formation and implications. Acta Biochim. Pol. 2013, 60, 277–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennig-Pauka, I.; Imker, R.; Mayer, L.; Brugmann, M.; Werckenthin, C.; Weber, H.; Menrath, A.; de Buhr, N. From stable to lab-investigating key factors for sudden deaths caused by Streptococcus suis. Pathogens 2019, 8, 249. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Lin, L.; Fu, L.; Han, L.; Zhang, A. Neutrophil extracellular traps play an important role in clearance of Streptococcus suis in vivo. Microbiol. Immunol. 2016, 60, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH oxidase activation and bacterial resistance. Front. Cell. Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dai, M.; Yuan, Z. Methods for the detection of reactive oxygen species. Anal. Methods 2018, 10, 4625–4638. [Google Scholar] [CrossRef]
- Segal, A.W. How neutrophils kill microbes. Annu. Rev. Immunol. 2005, 23, 197–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, S.M.; Corriden, R.; Nizet, V. The ontogeny of a neutrophil: Mechanisms of granulopoiesis and homeostasis. Microbiol. Mol. Biol. Rev. 2018, 82, e00057-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Cellular and Molecular Immunology, 8th ed.; Elsevier Saunders: Philadelphia, PA, USA, 2015; p. 535. [Google Scholar]
- Dutkiewicz, J.; Zajac, V.; Sroka, J.; Wasinski, B.; Cisak, E.; Sawczyn, A.; Kloc, A.; Wojcik-Fatla, A. Streptococcus suis: A re-emerging pathogen associated with occupational exposure to pigs or pork products. Part II—Pathogenesis. Ann. Agric. Environ. Med. 2018, 25, 186–203. [Google Scholar] [CrossRef] [PubMed]
- Wewer, C.; Seibt, A.; Wolburg, H.; Greune, L.; Schmidt, M.A.; Berger, J.; Galla, H.J.; Quitsch, U.; Schwerk, C.; Schroten, H.; et al. Transcellular migration of neutrophil granulocytes through the blood-cerebrospinal fluid barrier after infection with Streptococcus suis. J. Neuroinflamm. 2011, 8, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Besung, I.N.K.; Suarjana, I.G.K.; Agustina, K.K.; Winaya, I.B.O.; Soeharsono, H.; Suwiti, N.K.; Mahardika, G.N. Isolation and identification of Streptococcus suis from sick pigs in Bali, Indonesia. BMC Res. Notes 2019, 12, 795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, P.; Zhao, Y.X.; Zhang, A.D.; Kang, C.; Chen, H.C.; Jin, M.L. Pathologic analysis of the brain from Streptococcus suis type 2 experimentally infected pigs. Vet. Pathol. 2009, 46, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Niu, X.; Mei, W.; Li, W.; Liu, Y.; Willias, S.P.; Yuan, C.; Bei, W.; Wang, X.; Li, J. Immunogenicity and protective capacity of EF-Tu and FtsZ of Streptococcus suis serotype 2 against lethal infection. Vaccine 2018, 36, 2581–2588. [Google Scholar] [CrossRef] [PubMed]
- Seitz, M.; Beineke, A.; Singpiel, A.; Willenborg, J.; Dutow, P.; Goethe, R.; Valentin-Weigand, P.; Klos, A.; Baums, C.G. Role of capsule and suilysin in mucosal infection of complement-deficient mice with Streptococcus suis. Infect. Immun. 2014, 82, 2460–2471. [Google Scholar] [CrossRef] [Green Version]
- OuYang, X.; Guo, J.; Lv, Q.; Jiang, H.; Zheng, Y.; Liu, P.; Zhao, T.; Kong, D.; Hao, H.; Jiang, Y. TRIM32 drives pathogenesis in streptococcal toxic shock-like syndrome and Streptococcus suis meningitis by regulating innate immune responses. Infect. Immun. 2020, 88, e00957-19. [Google Scholar] [CrossRef]
- Liu, M.; Li, N.; Guo, W.; Jia, L.; Jiang, H.; Li, Z.; Wang, J.; Zhang, X.; Zhu, R.; Bao, C.; et al. RPSA distribution and expression in tissues and immune cells of pathogen-infected mice. Microb. Pathog. 2021, 152, 104609. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.; Du, Y.; Mao, C.; Li, J.; Jin, M.; Sun, L.; Wang, Y. Immunogenicity and protective ability of RpoE against Streptococcus suis serotype 2. J. Appl. Microbiol. 2021, 130, 1075–1083. [Google Scholar] [CrossRef]
- Yang, C.; Zhao, J.; Lin, L.; Pan, S.; Fu, L.; Han, L.; Jin, M.; Zhou, R.; Zhang, A. Targeting TREM-1 signaling in the presence of antibiotics is effective against streptococcal toxic-shock-like syndrome (STSLS) caused by Streptococcus suis. Front. Cell. Infect. Microbiol. 2015, 5, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Lv, Y.; Li, Y.A.; Du, Y.; Guo, W.; Chu, D.; Wang, X.; Wang, S.; Shi, H. Live attenuated Salmonella enterica serovar Choleraesuis vector delivering a conserved surface protein enolase induces high and broad protection against Streptococcus suis serotypes 2, 7, and 9 in mice. Vaccine 2020, 38, 6904–6913. [Google Scholar] [CrossRef]
- Li, P.; Li, M.; Lindberg, M.R.; Kennett, M.J.; Xiong, N.; Wang, Y. PAD4 is essential for antibacterial innate immunity mediated by neutrophil extracellular traps. J. Exp. Med. 2010, 207, 1853–1862. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, J.; Liu, Y.; Yang, J.; Guo, L.; Ren, S.; Chen, Z.; Liu, Z.; Zhang, Y.; Qiu, W.; et al. Porcine reproductive and respiratory syndrome virus NADC30-like strain accelerates Streptococcus suis serotype 2 infection in vivo and in vitro. Transbound. Emerg. Dis. 2019, 66, 729–742. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, H.; Du, R.; Li, S.; Qu, G.; Zhu, R.; Zhao, S.; Gu, J.; Sun, C.; Feng, X.; et al. Characteristic comparison of meningitis and non-meningitis of Streptococcus suis in an experimentally infected porcine model. Inflammation 2018, 41, 368–377. [Google Scholar] [CrossRef] [PubMed]
- Brockmeier, S.L.; Loving, C.L.; Eberle, K.C.; Hau, S.J.; Mou, K.T.; Kehrli, M.E., Jr. Administration of granulocyte-colony stimulating factor (G-CSF) to pigs results in a longer mean survival time after exposure to Streptococcus suis. Vet. Microbiol. 2019, 231, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Mu, Y.; Zhang, Y.; Dong, W.; Zhu, Y.; Ma, J.; Song, W.; Pan, Z.; Lu, C.; Yao, H. Antibacterial effect of porcine PTX3 against Streptococcus suis type 2 infection. Microb. Pathog. 2015, 89, 128–139. [Google Scholar] [CrossRef]
- Yang, C.; Chen, B.; Zhao, J.; Lin, L.; Han, L.; Pan, S.; Fu, L.; Jin, M.; Chen, H.; Zhang, A. TREM-1 signaling promotes host defense during the early stage of infection with highly pathogenic Streptococcus suis. Infect. Immun. 2015, 83, 3293–3301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Lu, X.; Xiao, P.; Liu, R.; Xia, K.L.; Wu, M.Z.; Jin, M.L.; Zhang, A.D. Interleukin-17A contributes to bacterial clearance in a mouse model of streptococcal toxic shock-like syndrome. Pathogens 2021, 10, 766. [Google Scholar] [CrossRef] [PubMed]
- Auger, J.P.; Rivest, S.; Benoit-Biancamano, M.O.; Segura, M.; Gottschalk, M. Inflammatory monocytes and neutrophils regulate Streptococcus suis-induced systemic inflammation and disease but are not critical for the development of central nervous system disease in a mouse model of infection. Infect. Immun. 2020, 88, e00787-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camicia, G.; Pozner, R.; de Larranaga, G. Neutrophil extracellular traps in sepsis. Shock 2014, 42, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, T.; Fisher, J.; Bakochi, A.; Neumann, A.; Cardoso, J.F.P.; Karlsson, C.A.Q.; Pavan, C.; Lundgaard, I.; Nilson, B.; Reinstrup, P.; et al. Neutrophil extracellular traps in the central nervous system hinder bacterial clearance during pneumococcal meningitis. Nat. Commun. 2019, 10, 1667. [Google Scholar] [CrossRef]
- Han, L.; Fu, L.; Peng, Y.; Zhang, A. Triggering receptor expressed on myeloid cells-1 signaling: Protective and pathogenic roles on streptococcal toxic-shock-like syndrome caused by Streptococcus suis. Front. Immunol. 2018, 9, 577. [Google Scholar] [CrossRef] [PubMed]
- Bleuze, M.; Auger, J.P.; Lavagna, A.; Gisch, N.; Gottschalk, M.; Segura, M. In vitro characterization of granulocyte-colony stimulating factor (G-CSF) production by dendritic cells and macrophages during Streptococcus suis infection. Immunobiology 2020, 225, 151979. [Google Scholar] [CrossRef]
- Dai, J.; Lai, L.; Tang, H.; Wang, W.; Wang, S.; Lu, C.; Yao, H.; Fan, H.; Wu, Z. Streptococcus suis synthesizes deoxyadenosine and adenosine by 5’-nucleotidase to dampen host immune responses. Virulence 2018, 9, 1509–1520. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Granule Name | Protein Content |
|---|---|
| Azurophilic or primary granules | myeloperoxidase (MPO) elastases (NE) heparin-binding protein (HBP) proteinases defensins |
| Specific or secondary granules | lactoferrin cathelicidin lysozyme membrane proteins including subunits of the NAPDH oxidase |
| Gelatinase or tertiary granules | gelatinases lysozyme membrane receptors |
| Secretory vesicles | HBP membrane proteins including complement receptors |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bleuzé, M.; Gottschalk, M.; Segura, M. Neutrophils in Streptococcus suis Infection: From Host Defense to Pathology. Microorganisms 2021, 9, 2392. https://doi.org/10.3390/microorganisms9112392
Bleuzé M, Gottschalk M, Segura M. Neutrophils in Streptococcus suis Infection: From Host Defense to Pathology. Microorganisms. 2021; 9(11):2392. https://doi.org/10.3390/microorganisms9112392
Chicago/Turabian StyleBleuzé, Marêva, Marcelo Gottschalk, and Mariela Segura. 2021. "Neutrophils in Streptococcus suis Infection: From Host Defense to Pathology" Microorganisms 9, no. 11: 2392. https://doi.org/10.3390/microorganisms9112392
APA StyleBleuzé, M., Gottschalk, M., & Segura, M. (2021). Neutrophils in Streptococcus suis Infection: From Host Defense to Pathology. Microorganisms, 9(11), 2392. https://doi.org/10.3390/microorganisms9112392