
Combined Therapy with microRNA-Expressing Salmonella and Irradiation in Melanoma


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of Attenuated Salmonella Strains Expressing miRNA
2.2. Transfection of Salmonella into Cells and Radiation
2.3. Salmonella Invasion Assay
2.4. LDH Assay
2.5. Protein Extraction and Immunoblot Analysis
2.6. Real-Time Quantitative PCR Analysis
2.7. Reactive Oxygen Species (ROS) Assay
2.8. Combined Treatment with S. Typhimurium and Irradiation
2.9. Melanoma Cancer Challenge in Mice
2.10. Bacterial Distribution and Cytokine Analysis in Mice
2.11. Statistical Analysis
3. Results
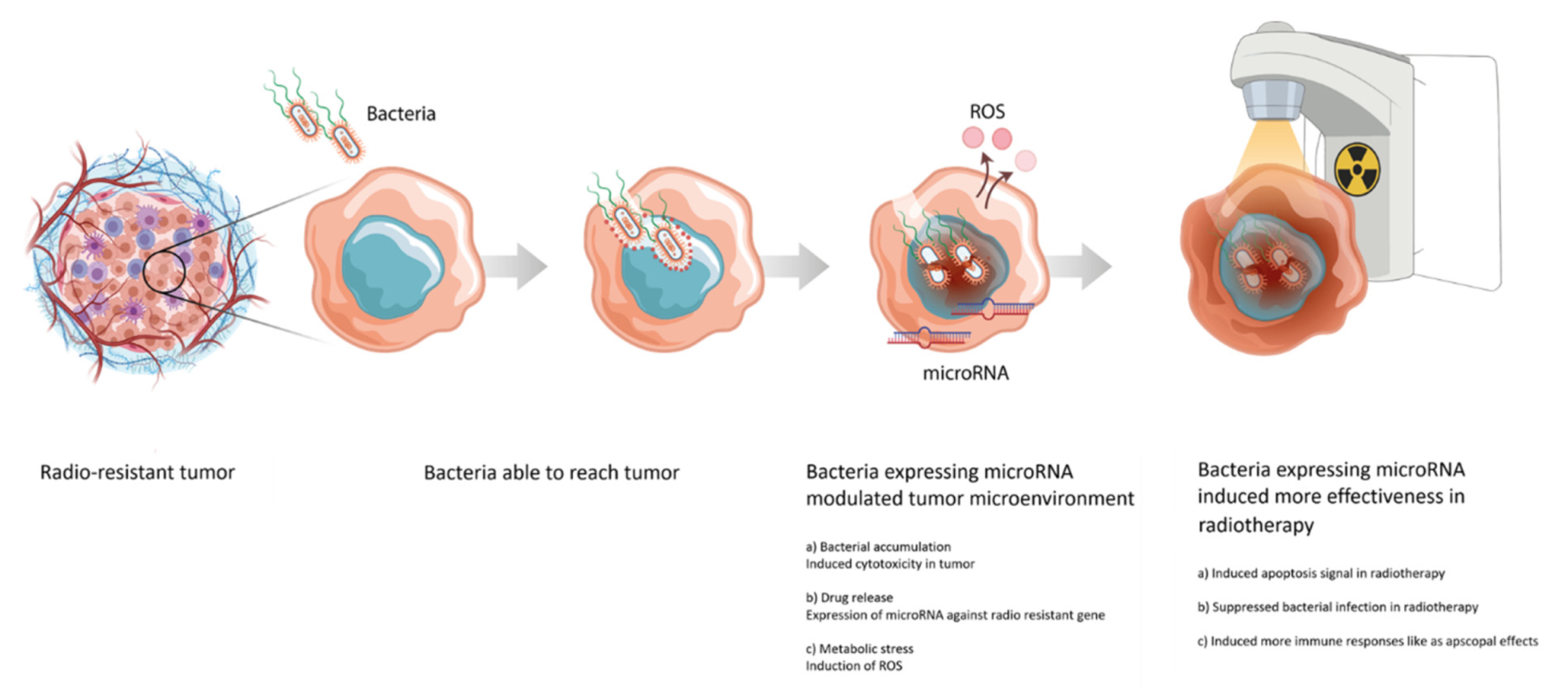
3.1. Application of Bacterial Treatments to Radiotherapy
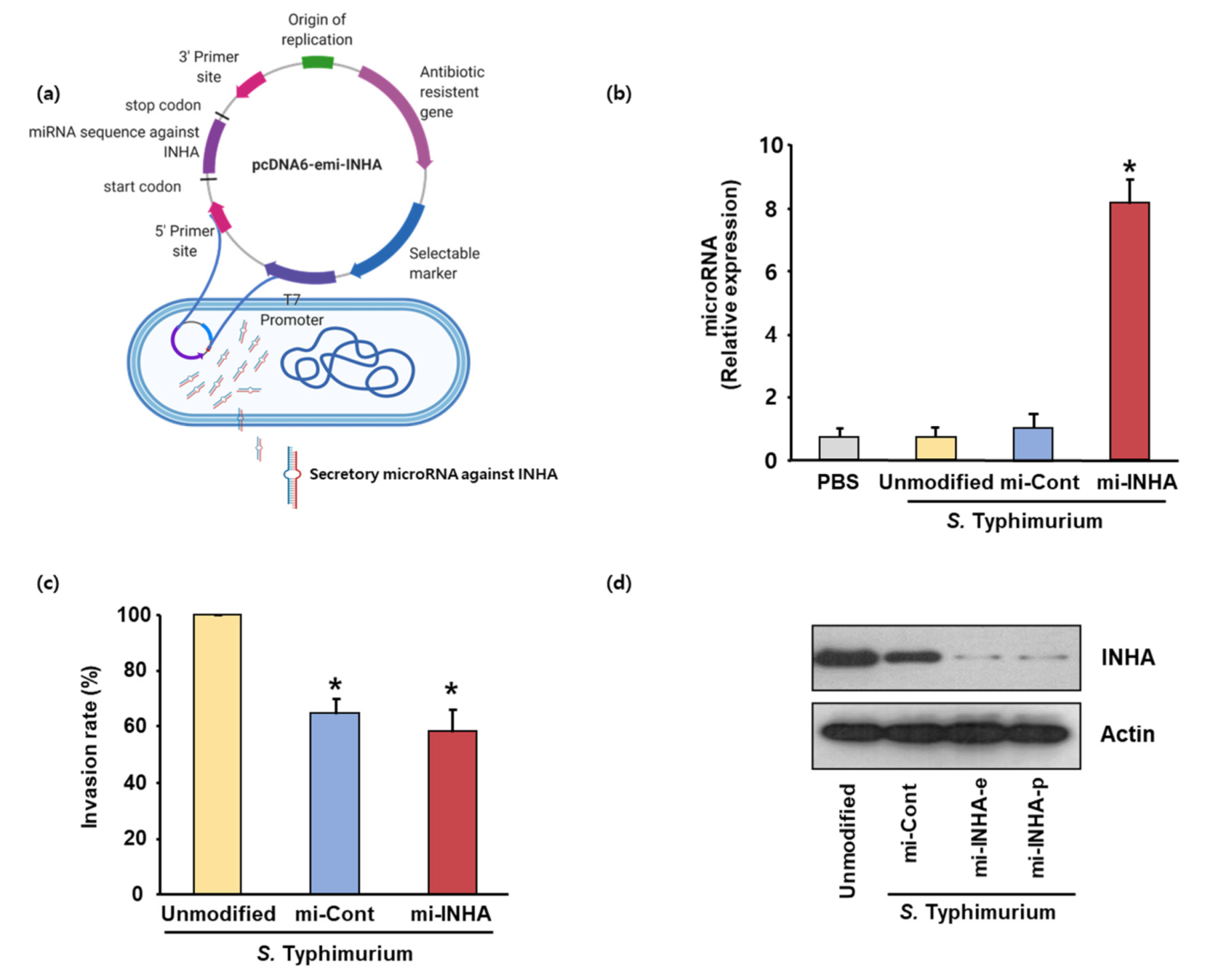
3.2. Construction of Recombinant Salmonella Expressing miRNA for Radioresistant Gene
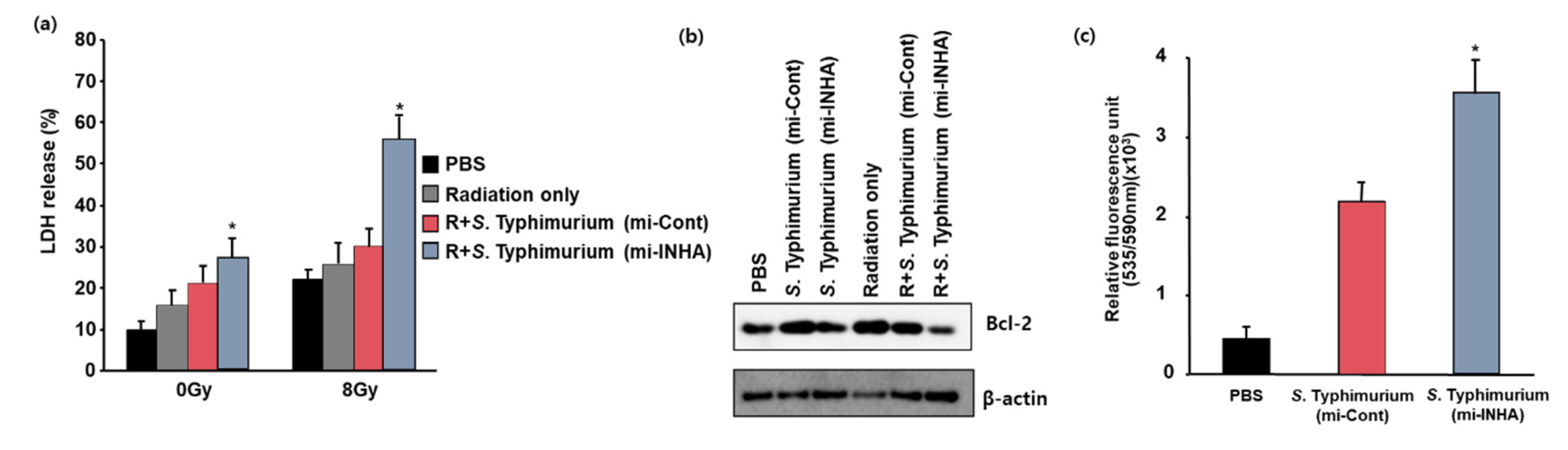
3.3. Reduction in Tumor Cell Viability by S. Typhimurium Expressing miRNA-INHA with Irradiation
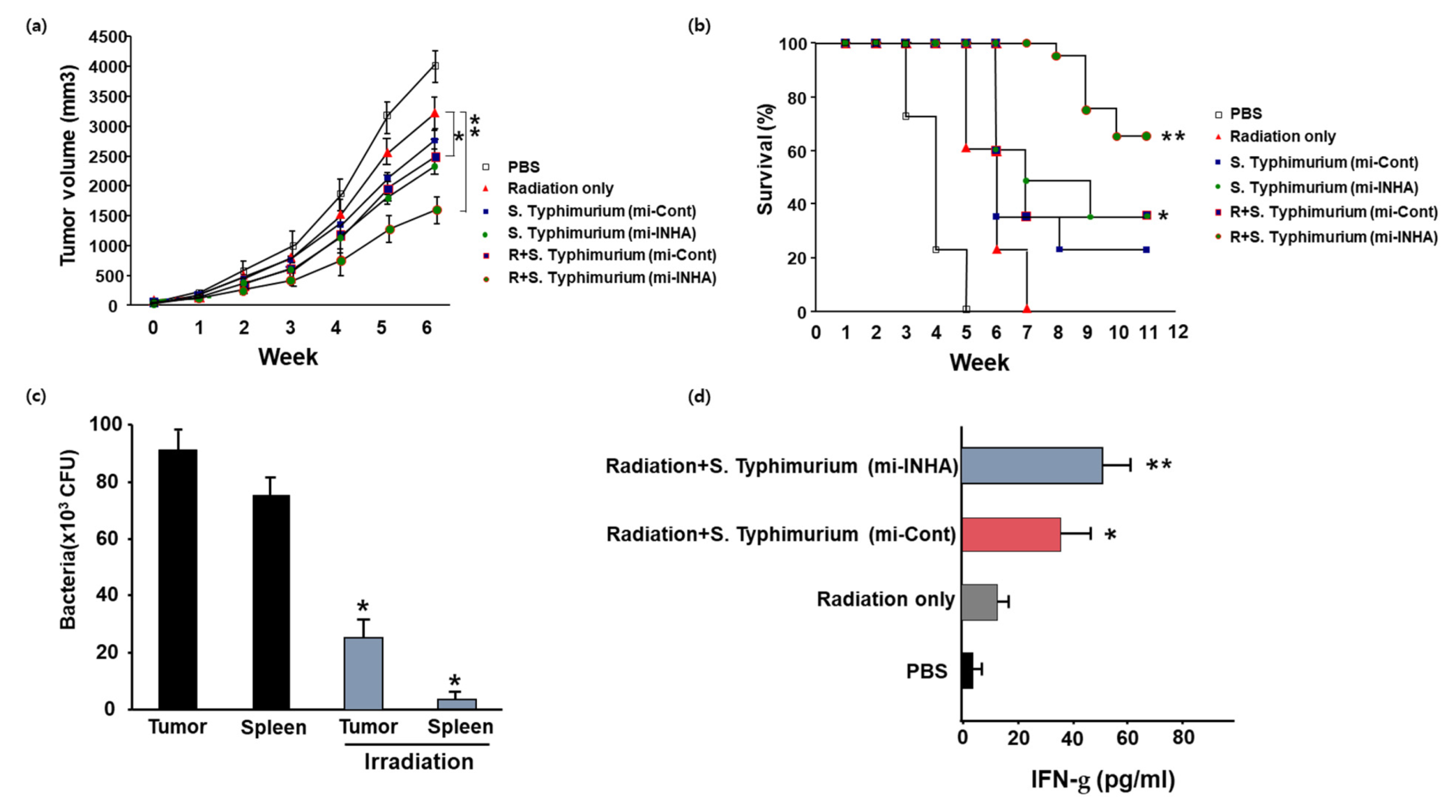
3.4. Effects of Combined Treatments with S. Typhimurium Expressing miRNA and Irradiation in Animals Bearing Melanoma
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Brown, J.M.; Wilson, W.R. Exploiting tumour hypoxia in cancer treatment. Nat. Rev. Cancer 2004, 4, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Cripps, A.; Wei, M.Q. New strategies for cancer gene therapy: Progress and opportunities. Clin. Exp. Pharmacol. Physiol. 2010, 37, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Byrne, N.M.; Al Jamal, W.; Coulter, J.A. Exploiting Current Understanding of Hypoxia Mediated Tumour Progression for Nanotherapeutic Development. Cancers 2019, 11, 1989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, K.; Liu, Q.; Li, P.; Luo, H.; Wang, H.; Kong, Q. Genetically engineered Salmonella Typhimurium: Recent advances in cancer therapy. Cancer Lett. 2019, 448, 168–181. [Google Scholar] [CrossRef]
- Phan, T.X.; Nguyen, V.H.; Duong, M.T.; Hong, Y.; Choy, H.E.; Min, J.J. Activation of inflammasome by attenuated Salmonella typhimurium in bacteria-mediated cancer therapy. Microbiol. Immunol. 2015, 59, 664–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, W.; Yoo, Y.; Chae, Y.S.; Kee, S.H.; Kim, B.M. Therapeutic advantage of genetically engineered Salmonella typhimurium carrying short hairpin RNA against inhibin alpha subunit in cancer treatment. Ann. Oncol. 2018, 29, 2010–2017. [Google Scholar] [CrossRef]
- Blache, C.A.; Manuel, E.R.; Kaltcheva, T.I.; Wong, A.N.; Ellenhorn, J.D.; Blazar, B.R.; Diamond, D.J. Systemic delivery of Salmonella typhimurium transformed with IDO shRNA enhances intratumoral vector colonization and suppresses tumor growth. Cancer Res. 2012, 72, 6447–6456. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.; Zhang, X.; Zhou, Y.; Zhang, C.; Hua, Z.C. A Salmonella Typhimurium mutant strain capable of RNAi delivery: Higher tumor-targeting and lower toxicity. Cancer Biol. Ther. 2014, 15, 1068–1076. [Google Scholar] [CrossRef] [Green Version]
- Deng, J.; Guo, Y.; Jiang, Z.; Yang, M.; Li, H.; Wang, J. Enhancement of ovarian cancer chemotherapy by delivery of multidrug-resistance gene small interfering RNA using tumor targeting Salmonella. J. Obstet. Gynaecol. Res. 2015, 41, 615–622. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Wang, B.; Ji, K.; Liang, Z.; Guo, B.; Hu, J.; Yin, D.; Du, Y.; Kopecko, D.J.; et al. Delivery of the co-expression plasmid pEndo-Si-Stat3 by attenuated Salmonella serovar typhimurium for prostate cancer treatment. J. Cancer Res. Clin. Oncol. 2013, 139, 971–980. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Yu, B.; Cai, C.H.; Huang, W.; Zheng, B.J.; Smith, D.K.; Huang, J.D. Combined prokaryotic-eukaryotic delivery and expression of therapeutic factors through a primed autocatalytic positive-feedback loop. J. Control. Release 2016, 222, 130–140. [Google Scholar] [CrossRef] [Green Version]
- Balanathan, P.; Williams, E.D.; Wang, H.; Pedersen, J.S.; Horvath, L.G.; Achen, M.G.; Stacker, S.A.; Risbridger, G.P. Elevated level of inhibin-alpha subunit is pro-tumourigenic and pro-metastatic and associated with extracapsular spread in advanced prostate cancer. Br. J. Cancer 2009, 100, 1784–1793. [Google Scholar] [CrossRef] [Green Version]
- Chrusciel, M.; Doroszko, M.; Stelmaszewska, J.; Li, X.; Ziecik, A.J.; Coelingh-Bennink, H.J.; Huhtaniemi, I.; Rahman, N.A. Transgenic mice expressing inhibin alpha-subunit promoter (inhalpha)/Simian Virus 40 T-antigen (Tag) transgene as a model for the therapy of granulosa cell-derived ovarian cancer. Reprod. Biol. 2014, 14, 25–31. [Google Scholar] [CrossRef] [PubMed]
- De Jong, F.H. Inhibin. Physiol. Rev. 1988, 68, 555–607. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Jenkins, L.M.; Horst, B.; Alers, V.; Pradhan, S.; Kaur, P.; Srivastava, T.; Hempel, N.; Gyorffy, B.; Broude, E.V.; et al. Inhibin Is a Novel Paracrine Factor for Tumor Angiogenesis and Metastasis. Cancer Res. 2018, 78, 2978–2989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, Z.; Shen, X.; Zhang, X.; Li, F.; Amevor, F.K.; Zhu, Q.; Wang, Y.; Li, D.; Shu, G.; Tian, Y.; et al. A functional polymorphism of inhibin alpha subunit at miR-181b-1-3p-binding site regulates proliferation and apoptosis of chicken ovarian granular cells. Cell Tissue Res. 2021, 384, 545–560. [Google Scholar] [CrossRef]
- Dayal, R.; Singh, A.; Pandey, A.; Mishra, K.P. Reactive oxygen species as mediator of tumor radiosensitivity. J. Cancer Res. Ther. 2014, 10, 811–818. [Google Scholar] [CrossRef]
- Loo, G. Redox-sensitive mechanisms of phytochemical-mediated inhibition of cancer cell proliferation (review). J. Nutr. Biochem. 2003, 14, 64–73. [Google Scholar] [CrossRef]
- Abshire, D.; Lang, M.K. The Evolution of Radiation Therapy in Treating Cancer. Semin. Oncol. Nurs. 2018, 34, 151–157. [Google Scholar] [CrossRef]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.W. Cancer and radiation therapy: Current advances and future directions. Int. J. Med. Sci. 2012, 9, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, T.S. Radiation sensitizers and targeted therapies. Oncology (Williston Park) 2003, 17, 23–28. [Google Scholar] [PubMed]
- Thanekar, A.M.; Sankaranarayanan, S.A.; Rengan, A.K. Role of nano-sensitizers in radiation therapy of metastatic tumors. Cancer Treat. Res. Commun. 2021, 26, 100303. [Google Scholar] [CrossRef] [PubMed]
- Al-Qadami, G.; Van Sebille, Y.; Le, H.; Bowen, J. Gut microbiota: Implications for radiotherapy response and radiotherapy-induced mucositis. Expert Rev. Gastroenterol. Hepatol. 2019, 13, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, Q.; Zhou, C.; Chen, K.; Chang, H.; Xiao, W.; Gao, Y. Colorectal cancer, radiotherapy and gut microbiota. Chin. J. Cancer Res. 2019, 31, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Clerc, P.L.; Berthon, B.; Claret, M.; Sansonetti, P.J. Internalization of Shigella flexneri into HeLa cells occurs without an increase in cytosolic Ca2+ concentration. Infect. Immun. 1989, 57, 2919–2922. [Google Scholar] [CrossRef] [Green Version]
- Kohler, H.; Rodrigues, S.P.; Maurelli, A.T.; McCormick, B.A. Inhibition of Salmonella typhimurium enteropathogenicity by piperidine, a metabolite of the polyamine cadaverine. J. Infect. Dis. 2002, 186, 1122–1130. [Google Scholar] [CrossRef]
- Kopecka, J.; Godel, M.; Riganti, C. Cholesterol metabolism: At the cross road between cancer cells and immune environment. Int. J. Biochem. Cell Biol. 2020, 129, 105876. [Google Scholar] [CrossRef]
- Leone, R.D.; Powell, J.D. Metabolism of immune cells in cancer. Nat. Rev. Cancer 2020, 20, 516–531. [Google Scholar] [CrossRef]
- Nencioni, A.; Caffa, I.; Cortellino, S.; Longo, V.D. Fasting and cancer: Molecular mechanisms and clinical application. Nat. Rev. Cancer 2018, 18, 707–719. [Google Scholar] [CrossRef]
- Baskar, R.; Dai, J.; Wenlong, N.; Yeo, R.; Yeoh, K.W. Biological response of cancer cells to radiation treatment. Front. Mol. Biosci. 2014, 1, 24. [Google Scholar] [CrossRef] [Green Version]
- Ngwa, W.; Irabor, O.C.; Schoenfeld, J.D.; Hesser, J.; Demaria, S.; Formenti, S.C. Using immunotherapy to boost the abscopal effect. Nat. Rev. Cancer 2018, 18, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Uribe-Herranz, M.; Rafail, S.; Beghi, S.; Gil-de-Gomez, L.; Verginadis, I.; Bittinger, K.; Pustylnikov, S.; Pierini, S.; Perales-Linares, R.; Blair, I.A.; et al. Gut microbiota modulate dendritic cell antigen presentation and radiotherapy-induced antitumor immune response. J. Clin. Investig. 2020, 130, 466–479. [Google Scholar] [CrossRef]
- Mortezaee, K. Immune escape: A critical hallmark in solid tumors. Life Sci. 2020, 258, 118110. [Google Scholar] [CrossRef] [PubMed]
- Spranger, S.; Gajewski, T.F. Impact of oncogenic pathways on evasion of antitumour immune responses. Nat. Rev. Cancer 2018, 18, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Mi, Z.; Feng, Z.C.; Li, C.; Yang, X.; Ma, M.T.; Rong, P.F. Salmonella-Mediated Cancer Therapy: An Innovative Therapeutic Strategy. J. Cancer 2019, 10, 4765–4776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, V.H.; Kim, H.S.; Ha, J.M.; Hong, Y.; Choy, H.E.; Min, J.J. Genetically engineered Salmonella typhimurium as an imageable therapeutic probe for cancer. Cancer Res. 2010, 70, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Wei, M.Q.; Mengesha, A.; Good, D.; Anne, J. Bacterial targeted tumour therapy-dawn of a new era. Cancer Lett. 2008, 259, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.W.L.; Tsolis, R.M.; Baumler, A.J. Salmonella versus the Microbiome. Microbiol. Mol. Biol. Rev. 2021, 85, e00027-19. [Google Scholar] [CrossRef]
- Bohnhoff, M.; Miller, C.P.; Martin, W.R. Resistance of the Mouse’s Intestinal Tract to Experimental Salmonella Infection. I. Factors Which Interfere with the Initiation of Infection by Oral Inoculation. J. Exp. Med. 1964, 120, 805–816. [Google Scholar] [CrossRef] [Green Version]
- Shukla, A.; Parmar, P.; Saraf, M. Radiation, radionuclides and bacteria: An in-perspective review. J. Environ. Radioact. 2017, 180, 27–35. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yoon, W.; Park, Y.; Kim, S.; Park, Y.; Kim, C.Y. Combined Therapy with microRNA-Expressing Salmonella and Irradiation in Melanoma. Microorganisms 2021, 9, 2408. https://doi.org/10.3390/microorganisms9112408
Yoon W, Park Y, Kim S, Park Y, Kim CY. Combined Therapy with microRNA-Expressing Salmonella and Irradiation in Melanoma. Microorganisms. 2021; 9(11):2408. https://doi.org/10.3390/microorganisms9112408
Chicago/Turabian StyleYoon, Wonsuck, Yongsung Park, Seunghyun Kim, Yongkeun Park, and Chul Yong Kim. 2021. "Combined Therapy with microRNA-Expressing Salmonella and Irradiation in Melanoma" Microorganisms 9, no. 11: 2408. https://doi.org/10.3390/microorganisms9112408
APA StyleYoon, W., Park, Y., Kim, S., Park, Y., & Kim, C. Y. (2021). Combined Therapy with microRNA-Expressing Salmonella and Irradiation in Melanoma. Microorganisms, 9(11), 2408. https://doi.org/10.3390/microorganisms9112408