
Exploration of Microalgal Species for Nutrient Removal from Anaerobically Digested Swine Wastewater and Potential Lipids Production



Abstract
:1. Introduction
2. Materials and Methods
2.1. Wastewater Collection, Pretreatment, and Analysis
2.2. Strains, Culture Conditions and Microalgae Selection
2.3. Measurement of Cell Growth
2.4. Nutrient Removal Analysis
2.5. Determination of Biochemical Composition of Algal Cells
2.6. Lipid and Fatty Acid Analysis
2.7. Statistical Analysis
3. Results and Discussion
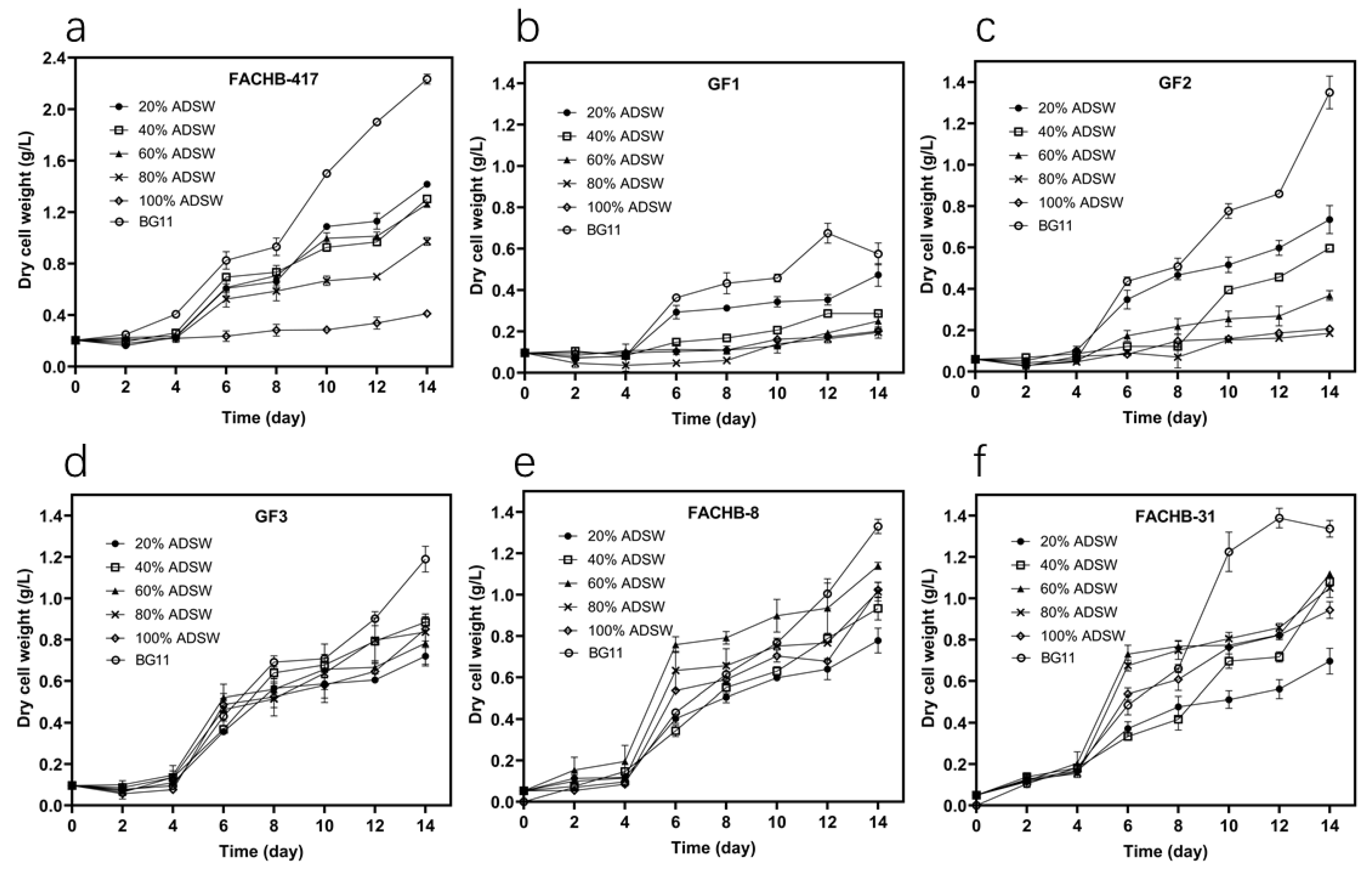
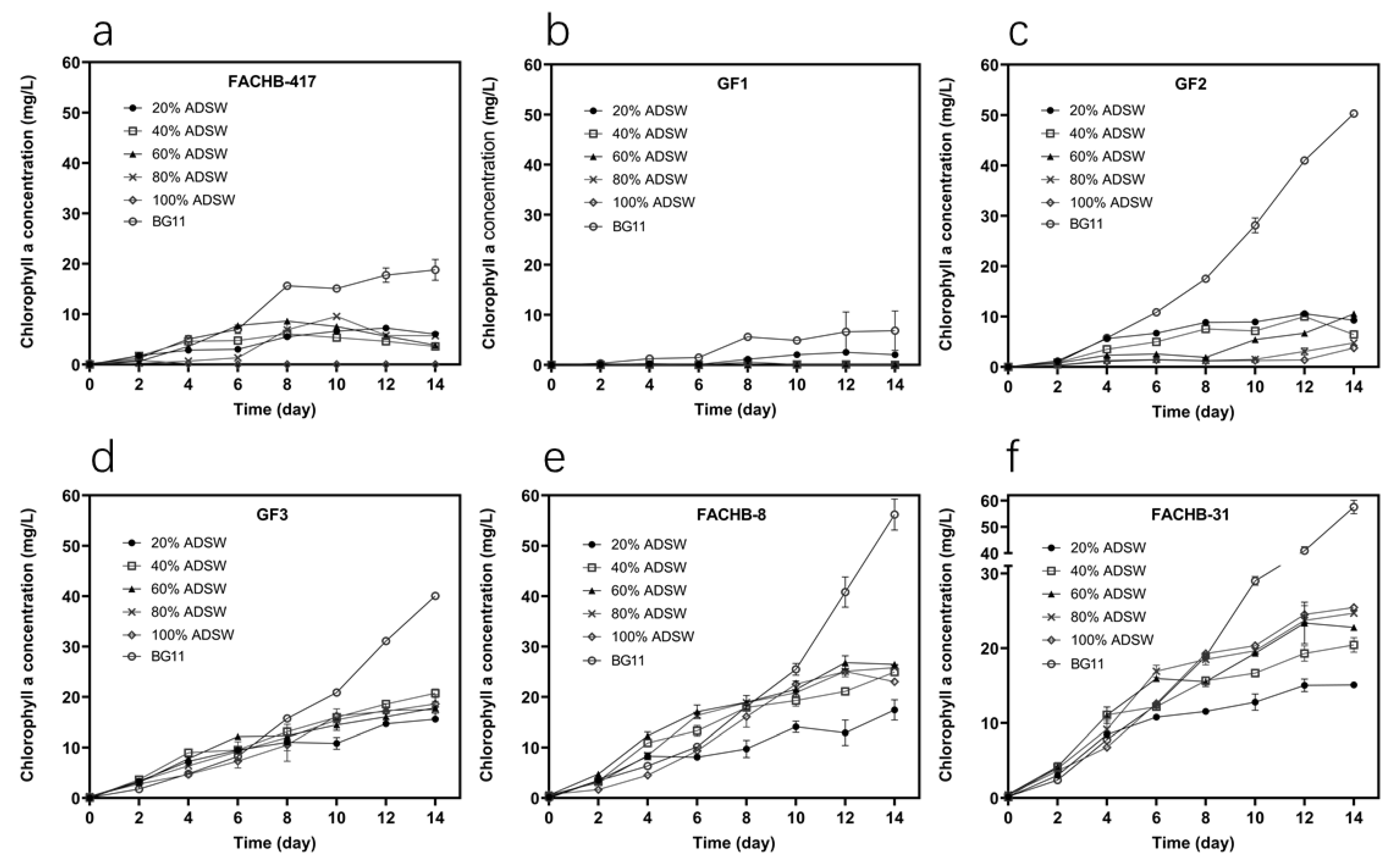
3.1. Selection of Microalgae Based on Growth Performance
3.2. Selection of Microalgae Based on Nutrient Removal Ability
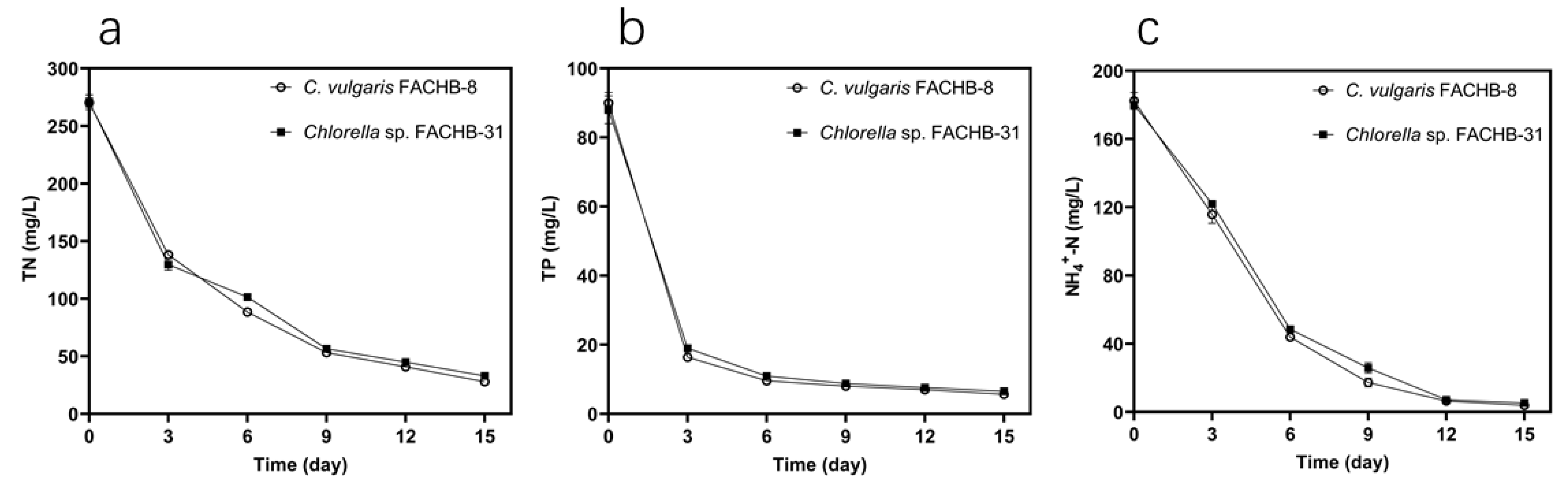
3.3. Nutrient Removal Ability of Two Selected Strains in Undiluted ADSW
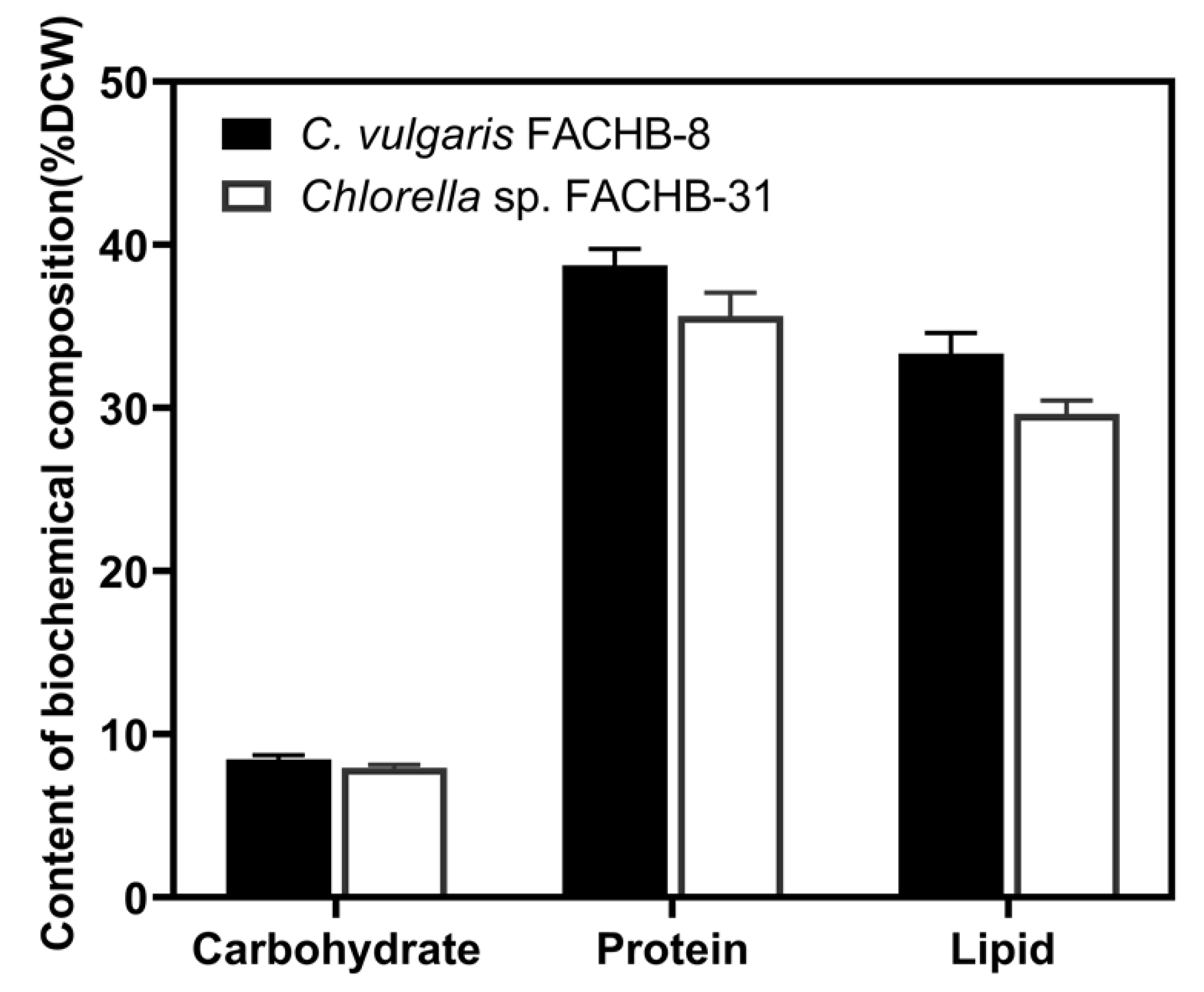
3.4. Lipid Production, Biomass Composition, and FAME Composition
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, J.; Qian, W.; Du, J.; Liu, Y. Effective allocation of resources in water pollution treatment alternatives: A multi-stage gray group decision-making method based on hesitant fuzzy linguistic term sets. Environ. Sci. Pollut. Res. Int. 2020, 27, 3173–3186. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Zhang, K.; Zhi, S.; Li, J.; Tian, X.; Gu, Y.; Zhou, J. High prevalence and dissemination of β-lactamase genes in swine farms in northern China. Sci. Total Environ. 2019, 651, 2507–2513. [Google Scholar] [CrossRef]
- Dan, N.H.; Rene, E.R.; Le Luu, T. Removal of Nutrients From Anaerobically Digested Swine Wastewater Using an Intermittent Cycle Extended Aeration System. Front. Microbiol. 2020, 11, 576438. [Google Scholar] [CrossRef]
- Griffith, A.W.; Gobler, C.J. Harmful algal blooms: A climate change co-stressor in marine and freshwater ecosystems. Harmful Algae 2020, 91, 101590. [Google Scholar] [CrossRef] [PubMed]
- Al-Jabri, H.; Das, P.; Khan, S.; Thaher, M.; AbdulQuadir, M. Treatment of Wastewaters by Microalgae and the Potential Applications of the Produced Biomass—A Review. Water 2021, 13, 27. [Google Scholar] [CrossRef]
- Nagarajan, D.; Kusmayadi, A.; Yen, H.W.; Dong, C.D.; Lee, D.J.; Chang, J.S. Current advances in biological swine wastewater treatment using microalgae-based processes. Bioresour. Technol. 2019, 289, 121718. [Google Scholar] [CrossRef]
- Hom-Diaz, A.; Jaén-Gil, A.; Bello-Laserna, I.; Rodríguez-Mozaz, S.; Vicent, T.; Barceló, D.; Blánquez, P. Performance of a microalgal photobioreactor treating toilet wastewater: Pharmaceutically active compound removal and biomass harvesting. Sci. Total Environ. 2017, 592, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tale, M.; Ghosh, S.; Kapadnis, B.; Kale, S. Isolation and characterization of microalgae for biodiesel production from Nisargruna biogas plant effluent. Bioresour. Technol. 2014, 169, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yang, Y.; Chen, Z.; Chen, Y.; Wen, Y.; Chen, B. Removal of nutrients from undiluted anaerobically treated piggery wastewater by improved microalgae. Bioresour. Technol. 2016, 222, 130–138. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, J.; Chen, P.; Ji, C.; Kang, Q.; Lu, B.; Li, K.; Liu, J.; Ruan, R. Bio-mitigation of carbon dioxide using microalgal systems: Advances and perspectives. Renew. Sustain. Energy Rev. 2017, 76, 1163–1175. [Google Scholar] [CrossRef]
- Ye, S.; Gao, L.; Zhao, J.; An, M.; Wu, H.; Li, M. Simultaneous wastewater treatment and lipid production by Scenedesmus sp. HXY2. Bioresour. Technol. 2020, 302, 122903. [Google Scholar] [CrossRef] [PubMed]
- Aravantinou, A.F.; Theodorakopoulos, M.A.; Manariotis, I.D. Selection of microalgae for wastewater treatment and potential lipids production. Bioresour. Technol. 2013, 147, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Delgado, R.T.; Guarieiro, M.D.; Antunes, P.W.; Cassini, S.T.; Terreros, H.M.; Fernandes, V.D. Effect of nitrogen limitation on growth, biochemical composition, and cell ultrastructure of the microalga Picocystis salinarum. J. Appl. Phycol. 2021, 33, 2083–2092. [Google Scholar] [CrossRef]
- Nagi, M.; He, M.; Li, D.; Gebreluel, T.; Cheng, B.; Wang, C. Utilization of tannery wastewater for biofuel production: New insights on microalgae growth and biomass production. Sci. Rep. 2020, 10, 1530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Sun, Y.; Rathour, R.; Pandey, A.; Thakur, I.S.; Tsang, D.C.W. Algae as potential feedstock for the production of biofuels and value-added products: Opportunities and challenges. Sci. Total Environ. 2020, 716, 137116. [Google Scholar] [CrossRef]
- Yadav, G.; Dash, S.K.; Sen, R. A biorefinery for valorization of industrial waste-water and flue gas by microalgae for waste mitigation, carbon-dioxide sequestration and algal biomass production. Sci. Total Environ. 2019, 688, 129–135. [Google Scholar] [CrossRef]
- Ziganshina, E.E.; Bulynina, S.S.; Ziganshin, A.M. Assessment of Chlorella sorokiniana Growth in Anaerobic Digester Effluent. Plants 2021, 10, 478. [Google Scholar] [CrossRef]
- Li, K.; Liu, Q.; Fang, F.; Luo, R.; Lu, Q.; Zhou, W.; Huo, S.; Cheng, P.; Liu, J.; Addy, M.; et al. Microalgae-based wastewater treatment for nutrients recovery: A review. Bioresour. Technol. 2019, 291, 121934. [Google Scholar] [CrossRef]
- Moondra, N.; Jariwala, N.D.; Christian, R.A. Sustainable treatment of domestic wastewater through microalgae. Int. J. Phytoremediation 2020, 22, 1480–1486. [Google Scholar] [CrossRef]
- Luo, L.; He, H.; Yang, C.; Wen, S.; Zeng, G.; Wu, M.; Zhou, Z.; Lou, W. Nutrient removal and lipid production by Coelastrella sp. in anaerobically and aerobically treated swine wastewater. Bioresour. Technol. 2016, 216, 135–141. [Google Scholar] [CrossRef]
- Arbib, Z.; Ruiz, J.; Álvarez-Díaz, P.; Garrido-Pérez, C.; Perales, J.A. Capability of different microalgae species for phytoremediation processes: Wastewater tertiary treatment, CO2 bio-fixation and low cost biofuels production. Water Res. 2014, 49, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Fan, Y.; Xie, B.; Tang, X.; Guo, T.; Luo, L.; Liang, H. Immobilizing Microcystis aeruginosa and powdered activated carbon for the anaerobic digestate effluent treatment. Chemosphere 2020, 244, 125420. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Diaz, P.D.; Ruiz, J.; Arbib, Z.; Barragan, J.; Garrido-Perez, M.C.; Perales, J.A. Freshwater microalgae selection for simultaneous wastewater nutrient removal and lipid production. Algal Res. Biomass Biofuels Bioprod. 2017, 24, 477–485. [Google Scholar] [CrossRef]
- Franchino, M.; Comino, E.; Bona, F.; Riggio, V.A. Growth of three microalgae strains and nutrient removal from an agro-zootechnical digestate. Chemosphere 2013, 92, 738–744. [Google Scholar] [CrossRef]
- Park, J.; Jin, H.F.; Lim, B.R.; Park, K.Y.; Lee, K. Ammonia removal from anaerobic digestion effluent of livestock waste using green alga Scenedesmus sp. Bioresour. Technol. 2010, 101, 8649–8657. [Google Scholar] [CrossRef] [PubMed]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef]
- Ma, X.; Yao, L.; Yang, B.; Lee, Y.K.; Chen, F.; Liu, J. RNAi-mediated silencing of a pyruvate dehydrogenase kinase enhances triacylglycerol biosynthesis in the oleaginous marine alga Nannochloropsis Salina. Sci. Rep. 2017, 7, 11485. [Google Scholar] [CrossRef] [PubMed]
- Masuko, T.; Minami, A.; Iwasaki, N.; Majima, T.; Nishimura, S.; Lee, Y.C. Carbohydrate analysis by a phenol-sulfuric acid method in microplate format. Anal. Biochem. 2005, 339, 69–72. [Google Scholar] [CrossRef]
- Wen, Y.; He, Y.; Ji, X.; Li, S.; Chen, L.; Zhou, Y.; Wang, M.; Chen, B. Isolation of an indigenous Chlorella vulgaris from swine wastewater and characterization of its nutrient removal ability in undiluted sewage. Bioresour. Technol. 2017, 243, 247–253. [Google Scholar] [CrossRef]
- Amit; Chandra, R.; Ghosh, U.K.; Nayak, J.K. Phycoremediation potential of marine microalga Tetraselmis indica on secondary treated domestic sewage for nutrient removal and biodiesel production. Environ. Sci. Pollut. Res. Int. 2017, 24, 20868–20875. [Google Scholar] [CrossRef]
- Xinjie, W.; Xin, N.; Qilu, C.; Ligen, X.; Yuhua, Z.; Qifa, Z. Vetiver and Dictyosphaerium sp. co-culture for the removal of nutrients and ecological inactivation of pathogens in swine wastewater. J. Adv. Res. 2019, 20, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Ho, S.H.; Cheng, C.L.; Guo, W.Q.; Nagarajan, D.; Ren, N.Q.; Lee, D.J.; Chang, J.S. Perspectives on the feasibility of using microalgae for industrial wastewater treatment. Bioresour. Technol. 2016, 222, 485–497. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, R.; Dikshit, A.K.; Tong, Y.W. Carbon-dioxide biofixation and phycoremediation of municipal wastewater using Chlorella vulgaris and Scenedesmus obliquus. Environ. Sci. Pollut. Res. Int. 2018, 25, 20399–20406. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.K.; Kim, H.C.; Sapireddy, V.R.; Yun, H.S.; Abou-Shanab, R.A.; Choi, J.; Lee, W.; Timmes, T.C.; Inamuddin; Jeon, B.H. Simultaneous nutrient removal and lipid production from pretreated piggery wastewater by Chlorella vulgaris YSW-04. Appl. Microbiol. Biotechnol. 2013, 97, 2701–2710. [Google Scholar] [CrossRef]
- Zhou, G.J.; Ying, G.G.; Liu, S.; Zhou, L.J.; Chen, Z.F.; Peng, F.Q. Simultaneous removal of inorganic and organic compounds in wastewater by freshwater green microalgae. Environ. Sci. Process. Impacts 2014, 16, 2018–2027. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.X.; Zeng, Y.; Zhu, K.; Tan, X. Vehicle mix evaluation in Beijing’s passenger-car sector: From air pollution control perspective. Sci. Total Environ. 2021, 785, 147264. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, A.L.; Pires, J.C.M.; Simoes, M. A review on the use of microalgal consortia for wastewater treatment. Algal Res. Biomass Biofuels Bioprod. 2017, 24, 403–415. [Google Scholar] [CrossRef]
- Sun, Z.L.; Sun, L.Q.; Chen, G.Z. Microalgal Cultivation and Nutrient Removal from Digested Piggery Wastewater in a Thin-film Flat Plate Photobioreactor. Appl. Biochem. Biotechnol. 2019, 187, 1488–1501. [Google Scholar] [CrossRef]
- Han, L.; Pei, H.; Hu, W.; Han, F.; Song, M.; Zhang, S. Nutrient removal and lipid accumulation properties of newly isolated microalgal strains. Bioresour. Technol. 2014, 165, 38–41. [Google Scholar] [CrossRef]
- Aketo, T.; Hoshikawa, Y.; Nojima, D.; Yabu, Y.; Maeda, Y.; Yoshino, T.; Takano, H.; Tanaka, T. Selection and characterization of microalgae with potential for nutrient removal from municipal wastewater and simultaneous lipid production. J. Biosci. Bioeng. 2020, 129, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.F.; Teng, J.C.; Lin, Y.H.; Hwang, S.C. Increasing algal biofuel production using Nannocholropsis oculata cultivated with anaerobically and aerobically treated swine wastewater. Bioresour. Technol. 2013, 133, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.Y.; Yun, Y.M.; Shin, H.S.; Han, J.I. Cultivation of four microalgae species in the effluent of anaerobic digester for biodiesel production. Bioresour. Technol. 2017, 224, 738–742. [Google Scholar] [CrossRef]
- Sydney, E.B.; da Silva, T.E.; Tokarski, A.; Novak, A.C.; de Carvalho, J.C.; Woiciecohwski, A.L.; Larroche, C.; Soccol, C.R. Screening of microalgae with potential for biodiesel production and nutrient removal from treated domestic sewage. Appl. Energy 2011, 88, 3291–3294. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Non-Autoclaved ADSW | Autoclaved ADSW |
|---|---|---|
| TN (mg·L−1) | 341.50 ± 9.71 | 258.50 ± 6.36 |
| TP (mg·L−1) | 80.75 ± 0.49 | 65.75 ± 0.64 |
| NH4+-N (mg·L−1) | 238.50 ± 4.95 | 183.50 ± 2.12 |
| COD (mg·L−1) | 1124.01 ± 23.46 | 900.01 ± 13.53 |
| TOC (mg·L−1) | 393.80 ± 10.87 | 194.80 ± 6.41 |
| IC (mg·L−1) | 324.12 ± 12.96 | 191.92 ± 7.05 |
| As (mg·L−1) | 0.00 | 0.00 |
| Cd (mg·L−1) | 0.00 | 0.00 |
| Cr (mg·L−1) | 0.86 | 0.65 |
| Cu (mg·L−1) | 0.21 | 0.03 |
| Fe (mg·L−1) | 3.66 | 2.19 |
| Mn (mg·L−1) | 0.45 | 0.18 |
| Ni (mg·L−1) | 0.16 | 0.08 |
| Pb (mg·L−1) | 3.90 | 3.90 |
| Zn (mg·L−1) | 0.12 | 0.05 |
| Sb (mg·L−1) | 0.005 | 0.004 |
| pH | 7.35 ± 0.21 | 8.36 ± 0.34 |
| Algal Species | Culture Medium | Biomass Production (g L−1) | Biomass Productivity (mg L−1 d−1) | Maximum Biomass Productivity (mg L−1 d−1) | Specific Growth Rate (d−1) | Maximum Specific Growth Rate (d−1) |
|---|---|---|---|---|---|---|
| S. obliquus FACHB-417 | 20% ADSW | 1.417 ± 0.016 b | 86.548 ± 0.546 b | 213.333 ± 17.559 b | 0.182 ± 0.009 b | 0.250 ± 0.031 c |
| 40% ADSW | 1.303 ± 0.010 c | 78.453 ± 1.148 c | 216.667 ± 17.017 b | 0.161 ± 0.012 bc | 0.490 ± 0.053 a | |
| 60% ADSW | 1.262 ± 0.025 c | 75.476 ± 2.431 c | 212.500 ± 18.284 b | 0.169 ± 0.021 bc | 0.485 ± 0.037 a | |
| 80% ADSW | 0.973 ± 0.031 d | 54.881 ± 2.508 d | 166.250 ± 8.839 c | 0.147 ± 0.007 c | 0.425 ± 0.044 ab | |
| 100% ADSW | 0.412 ± 0.010 e | 14.762 ± 1.443 e | 26.250 ± 5.303 d | 0.063 ± 0.011 d | 0.068 ± 0.010 d | |
| BG11 medium | 2.233 ± 0.039 a | 144.881 ± 3.037 a | 284.167 ± 33.572 a | 0.362 ± 0.009 a | 0.353 ± 0.037 bc | |
| C. zofingiensis GF1 | 20% ADSW | 0.473 ± 0.055 b | 26.905 ± 4.306 b | 106.667 ± 14.216 b | 0.178 ± 0.012 ab | 0.650 ± 0.054 b |
| 40% ADSW | 0.287 ± 0.015 c | 13.571 ± 1.429 c | 41.667 ± 10.104 c | 0.125 ± 0.025 bc | 0.163 ± 0.042 d | |
| 60% ADSW | 0.250 ± 0.035 cd | 10.953 ± 2.909 cd | 42.500 ± 3.536 c | 0.088 ± 0.030 c | 0.286 ± 0.018 c | |
| 80% ADSW | 0.195 ± 0.028 d | 7.024 ± 1.967 d | 40.000 ± 11.456 c | 0.185 ± 0.075 ab | 0.348 ± 0.037 c | |
| 100% ADSW | 0.202 ± 0.015 d | 7.500 ± 1.237 d | 16.250 ± 5.303 d | 0.074 ± 0.013 c | 0.156 ± 0.010 d | |
| BG11 medium | 0.575 ± 0.053 a | 33.929 ± 3.763 a | 140.833 ± 10.104 a | 0.195 ± 0.003 a | 0.748 ± 0.064 a | |
| C. protothecoides GF2 | 20% ADSW | 0.735 ± 0.068 b | 48.214 ± 4.831 b | 123.333 ± 28.759 b | 0.199 ± 0.020 b | 0.6863 ± 0.294 a |
| 40% ADSW | 0.597 ± 0.018 c | 38.333 ± 1.254 c | 136.667 ± 3.819 b | 0.194 ± 0.015 b | 0.589 ± 0.025 b | |
| 60% ADSW | 0.368 ± 0.024 d | 22.024 ± 1.688 d | 60.833 ± 11.273 c | 0.196 ± 0.011 b | 0.602 ± 0.049 b | |
| 80% ADSW | 0.185 ± 0.013 e | 8.929 ± 0.945 e | 25.000 ± 5.000 d | 0.138 ± 0.018 c | 0.358 ±0.036 c | |
| 100% ADSW | 0.207 ± 0.003 e | 10.476 ± 0.206 e | 25.833 ± 1.443 d | 0.105 ± 0.019 d | 0.310 ± 0.048 c | |
| BG11 medium | 1.350 ± 0.079 a | 92.143 ± 5.669 a | 188.333 ± 12.583 a | 0.311 ± 0.009 a | 0.225 ± 0.023 d | |
| C. pyrenoidosa GF3 | 20% ADSW | 0.720 ± 0.048 c | 44.524 ± 3.079 c | 123.333 ± 10.408 d | 0.188 ± 0.013 b | 0.591 ± 0.073 c |
| 40% ADSW | 0.885 ± 0.040 b | 56.309 ± 3.221 b | 116.667 ± 9.465 d | 0.188 ± 0.004 b | 0.502 ± 0.047 c | |
| 60% ADSW | 0.783 ± 0.102 bc | 49.048 ± 7.092 bc | 211.333 ± 15.543 a | 0.170 ± 0.037 b | 0.747 ± 0.049 b | |
| 80% ADSW | 0.837 ± 0.075 bc | 52.619 ± 5.872 bc | 189.167 ± 7.638 b | 0.183 ± 0.030 b | 0.614 ± 0.172 bc | |
| 100% ADSW | 0.852 ± 0.058 b | 53.929 ± 4.345 b | 205.833 ± 11.547 ab | 0.241 ± 0.001 a | 0.926 ± 0.009 a | |
| BG11 medium | 1.190 ± 0.062 a | 78.095 ± 4.185 a | 168.333 ± 11.547 c | 0.253 ± 0.004 a | 0.757 ± 0.034 b | |
| C. vulgaris FACHB-8 | 20% ADSW | 0.778 ± 0.060 e | 51.786 ± 4.684 e | 141.667 ± 11.815 d | 0.189 ± 0.011 c | 0.614 ± 0.072 c |
| 40% ADSW | 0.933 ± 0.056 d | 62.857 ± 4.33 d | 106.667 ± 7.638 e | 0.185 ± 0.008 c | 0.424 ± 0.044 d | |
| 60% ADSW | 1.140 ± 0.017 b | 77.619 ± 1.649 b | 295.000 ± 25.372 a | 0.198 ± 0.009 bc | 0.770 ± 0.075 b | |
| 80% ADSW | 1.015 ± 0.045 c | 68.691 ± 2.865 cd | 239.833 ± 16.127 b | 0.218 ± 0.015 b | 0.852 ± 0.103 ab | |
| 100% ADSW | 1.023 ± 0.038 c | 69.285 ± 3.093 c | 225.833 ± 2.887 b | 0.249 ± 0.007 a | 0.923 ± 0.039 a | |
| BG11 medium | 1.330 ± 0.035 a | 91.190 ± 2.909 a | 174.1667 ± 7.638 c | 0.264 ± 0.020 a | 0.755 ± 0.122 bc | |
| Chlorella. sp. FACHB-31 | 20% ADSW | 0.697 ± 0.062 d | 46.190 ± 4.437 d | 95.000 ± 15.613 d | 0.142 ± 0.020 b | 0.364 ± 0.051 c |
| 40% ADSW | 1.080 ± 0.022 b | 73.572 ± 1.557 b | 161.667 ± 3.819 c | 0.178 ± 0.006 a | 0.292 ± 0.025 c | |
| 60% ADSW | 1.118 ± 0.012 b | 76.310 ± 0.825 b | 273.333 ± 16.646 ab | 0.173 ± 0.031 a | 0.586 ± 0.049 b | |
| 80% ADSW | 1.050 ± 0.046 b | 71.428 ± 3.273 b | 260.000 ± 8.660 b | 0.192 ± 0.009 a | 0.738 ± 0.052 a | |
| 100% ADSW | 0.943 ± 0.040 c | 63.810 ± 2.509 c | 187.500 ± 9.014 c | 0.176 ± 0.007 a | 0.598 ± 0.035 b | |
| BG11 medium | 1.337 ± 0.040 a | 87.500 ± 6.814 a | 292.500 ± 27.839 a | 0.199 ± 0.007 a | 0.319 ± 0.019 c |
| Algal Species | Culture Medium | TN | TP | NH4+-N | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Initial (mg L−1) | Final (mg L−1) | Removal Efficiency (%) | Initial (mg L−1) | Final (mg L−1) | Removal Efficiency (%) | Initial (mg L−1) | Final (mg L−1) | Removal Efficiency (%) | ||
| S. obliquus FACHB-417 | 20% ADSW | 52.43 ± 1.23 | 13.51 ± 0.70 | 74.24 ± 1.34 b | 14.21 ± 0.21 | 0.40 ± 0.05 | 97.19 ± 0.37 a | 37.46 ± 1.45 | 0.84 ± 0.03 | 97.75 ± 0.07 a |
| 40% ADSW | 105.71 ± 2.46 | 18.08 ± 1.25 | 82.89 ± 1.18 a | 26.87 ± 0.19 | 0.85 ± 0.12 | 96.85 ± 0.45 a | 73.60 ± 1.78 | 3.23 ± 0.15 | 95.61 ± 0.21 b | |
| 60% ADSW | 153.80 ± 4.21 | 46.92 ± 1.12 | 69.48 ± 0.73 d | 39.60 ± 0.43 | 4.80 ± 0.14 | 87.87 ± 0.34 c | 111.34 ± 3.98 | 12.00 ± 0.84 | 89.22 ± 0.76 c | |
| 80% ADSW | 205.80 ± 5.46 | 57.84 ± 0.71 | 71.91 ± 0.35 c | 53.46 ± 0.34 | 4.02 ± 0.03 | 92.47 ± 0.06 b | 147.87 ± 2.45 | 17.77 ± 1.89 | 87.98 ± 1.28 c | |
| 100% ADSW | 258.50 ± 6.36 | 71.33 ± 0.91 | 72.41 ± 0.35 c | 65.75 ± 0.64 | 9.79 ± 0.18 | 85.12 ± 0.27 d | 183.50 ± 2.12 | 59.75 ± 1.17 | 67.44 ± 0.64 d | |
| C. zofingiensis GF1 | 20% ADSW | 52.43 ± 1.23 | 13.15 ± 0.74 | 74.87 ± 1.40 a | 14.21 ± 0.21 | 5.39 ± 0.21 | 62.05 ± 1.44 b | 37.46 ± 1.45 | 13.22 ± 1.91 | 64.71 ± 5.10 a |
| 40% ADSW | 105.71 ± 2.46 | 29.28 ± 0.68 | 72.30 ± 0.64 b | 26.87 ± 0.19 | 15.57 ± 1.16 | 42.07 ± 4.31 d | 73.60 ± 1.78 | 26.23 ± 0.73 | 64.36 ± 1.00 a | |
| 60% ADSW | 153.80 ± 4.21 | 59.06 ± 1.26 | 61.60 ± 0.82 d | 39.60 ± 0.43 | 16.82 ± 0.31 | 57.52 ± 0.78 c | 111.34 ± 3.98 | 55.42 ± 3.37 | 50.22 ± 3.03 b | |
| 80% ADSW | 205.80 ± 5.46 | 69.61 ± 0.95 | 66.18 ± 0.46 c | 53.46 ± 0.34 | 18.76 ± 0.15 | 64.91 ± 0.28 b | 147.87 ± 2.45 | 71.97 ± 1.52 | 51.33 ± 1.03 b | |
| 100% ADSW | 258.50 ± 6.36 | 84.03 ± 1.80 | 67.49 ± 0.70 c | 65.75 ± 0.64 | 19.76 ± 0.45 | 69.94 ± 0.68 a | 183.50 ± 2.12 | 91.17 ± 3.15 | 50.32 ± 1.72 b | |
| C. protothecoides GF2 | 20% ADSW | 52.43 ± 1.23 | 10.59 ± 0.14 | 79.80 ± 0.26 b | 14.21 ± 0.21 | 2.38 ± 0.26 | 83.23 ± 1.85 d | 37.46 ± 1.45 | 2.00 ± 0.04 | 94.66 ± 0.12 b |
| 40% ADSW | 105.71 ± 2.46 | 17.43 ± 0.80 | 83.51 ± 0.75 a | 26.87 ± 0.19 | 0.65 ± 0.08 | 97.57 ± 0.28 a | 73.60 ± 1.78 | 0.74 ± 0.02 | 99.00 ± 0.03 a | |
| 60% ADSW | 153.80 ± 4.21 | 41.31 ± 0.94 | 73.14 ± 0.61 d | 39.60 ± 0.43 | 4.42 ± 0.40 | 88.83 ± 1.00 b | 111.34 ± 3.98 | 24.83 ± 1.00 | 77.70 ± 0.90 c | |
| 80% ADSW | 205.80 ± 5.46 | 53.41 ± 1.97 | 74.05 ± 0.95 d | 53.46 ± 0.34 | 6.29 ± 0.19 | 88.24 ± 0.36 bc | 147.87 ± 2.45 | 34.64 ± 0.86 | 76.57 ± 0.58 d | |
| 100% ADSW | 258.50 ± 6.36 | 60.83 ± 1.09 | 76.47 ± 0.42 c | 65.75 ± 0.64 | 8.74 ± 0.16 | 86.71 ± 0.24 c | 183.50 ± 2.12 | 47.80 ± 0.98 | 73.95 ± 0.54 e | |
| C. pyrenoidosa GF3 | 20% ADSW | 52.43 ± 1.23 | 25.90 ± 1.55 | 50.60 ± 2.96 d | 14.21 ± 0.21 | 1.09 ± 0.14 | 92.31 ± 1.02 b | 37.46 ± 1.45 | 1.45 ± 0.13 | 96.13 ± 0.35 b |
| 40% ADSW | 105.71 ± 2.46 | 32.51 ± 2.16 | 69.25 ± 2.04 c | 26.87 ± 0.19 | 1.36 ± 0.14 | 94.94 ± 0.54 a | 73.60 ± 1.78 | 1.81 ± 0.16 | 97.54 ± 0.22 a | |
| 60% ADSW | 153.80 ± 4.21 | 38.00 ± 0.90 | 75.29 ± 0.58 b | 39.60 ± 0.43 | 2.69 ± 0.28 | 93.22 ± 0.71 b | 111.34 ± 3.98 | 17.88 ± 0.89 | 83.94 ± 0.80 c | |
| 80% ADSW | 205.80 ± 5.46 | 48.14 ± 0.34 | 76.61 ± 0.16 b | 53.46 ± 0.34 | 5.04 ± 0.02 | 90.57 ± 0.04 c | 147.87 ± 2.45 | 21.87 ± 1.34 | 85.21 ± 0.91 c | |
| 100% ADSW | 258.50 ± 6.36 | 52.62 ± 1.02 | 79.64 ± 0.39 a | 65.75 ± 0.64 | 7.91 ± 0.14 | 87.96 ± 0.21 d | 183.50 ± 2.12 | 36.07 ± 1.18 | 80.35 ± 0.65 d | |
| C. vulgaris FACHB-8 | 20% ADSW | 52.43 ± 1.23 | 23.09 ± 0.08 | 55.95 ± 0.15 d | 14.21 ± 0.21 | 1.05 ± 0.14 | 92.63 ± 1.00 bc | 37.46 ± 1.45 | 1.67 ± 0.03 | 95.55 ± 0.09 b |
| 40% ADSW | 105.71 ± 2.46 | 27.95 ± 0.51 | 73.56 ± 0.48 c | 26.87 ± 0.19 | 1.24 ± 0.22 | 95.39 ± 0.81 a | 73.60 ± 1.78 | 1.62 ± 0.18 | 97.80 ± 0.25 a | |
| 60% ADSW | 153.80 ± 4.21 | 32.85 ± 0.90 | 78.64 ± 0.59 b | 39.60 ± 0.43 | 1.87 ± 0.10 | 95.29 ± 0.26 a | 111.34 ± 3.98 | 8.38 ± 0.17 | 92.48 ± 0.15 c | |
| 80% ADSW | 205.80 ± 5.46 | 41.90 ± 0.44 | 79.64 ± 0.21 b | 53.46 ± 0.34 | 3.57 ± 0.19 | 93.32 ± 0.36 b | 147.87 ± 2.45 | 12.73 ± 0.35 | 91.39 ± 0.24 d | |
| 100% ADSW | 258.50 ± 6.36 | 40.85 ± 1.28 | 84.21 ± 0.49 a | 65.75 ± 0.64 | 5.13 ± 0.13 | 92.19 ± 0.20 c | 183.50 ± 2.12 | 16.22 ± 0.47 | 91.16 ± 0.25 d | |
| Chlorella sp. FACHB-31 | 20% ADSW | 52.43 ± 1.23 | 18.48 ± 1.97 | 64.76 ± 1.76 d | 14.21 ± 0.21 | 0.77 ± 0.03 | 94.60 ± 0.21 b | 37.46 ± 1.45 | 2.26 ± 0.15 | 93.98 ± 0.40 b |
| 40% ADSW | 105.71 ± 2.46 | 25.94 ± 0.32 | 75.46 ± 0.32 c | 26.87 ± 0.19 | 0.98 ± 0.02 | 96.35 ± 0.07 a | 73.60 ± 1.78 | 1.54 ± 0.32 | 97.91 ± 0.43 a | |
| 60% ADSW | 153.80 ± 4.21 | 35.33 ± 0.94 | 77.03 ± 0.61 bc | 39.60 ± 0.43 | 2.76 ± 0.15 | 93.03 ± 0.39 c | 111.34 ± 3.98 | 9.89 ± 0.42 | 91.12 ± 0.38 c | |
| 80% ADSW | 205.80 ± 5.46 | 41.87 ± 2.36 | 79.66 ± 1.14 b | 53.46 ± 0.34 | 3.69 ± 0.31 | 93.10 ± 0.58 c | 147.87 ± 2.45 | 12.44 ± 0.47 | 91.59 ± 0.32 c | |
| 100% ADSW | 258.50 ± 6.36 | 40.75 ± 0.59 | 84.24 ± 0.23 a | 65.75 ± 0.64 | 6.01 ± 0.14 | 90.89 ± 0.21 d | 183.50 ± 2.12 | 17.47 ± 0.52 | 90.48 ± 0.28 d | |
| Fatty Acid | C. vulgaris FACHB-8 | Chlorella sp. FACHB-31 |
|---|---|---|
| C14:0 | 8.52 ± 0.69 | 8.23 ± 1.09 |
| C14:1 | 2.21 ± 0.03 | 2.37 ± 0.20 |
| C16:0 | 27.46 ± 0.32 | 27.95 ± 0.37 |
| C16:1 | 2.81 ± 0.13 | 2.40 ± 0.70 |
| C18:0 | 17.59 ± 1.26 | 17.29 ± 0.82 |
| C18:1 n-9 | 20.57 ± 0.07 | 20.25 ± 0.44 |
| C18:2 n-6 | 16.61 ± 1.09 | 17.27 ± 0.11 |
| C20:0 | 3.55 ± 0.86 | 3.58 ± 0.91 |
| C20:5 n-3 | 0.67 ± 0.12 | 0.65 ± 0.10 |
| SFA a | 56.74 ± 0.56 | 57.41 ± 0.50 |
| UFA b | 43.26 ± 0.56 | 42.59 ± 0.50 |
| MUFA c | 25.62 ± 0.04 | 24.65 ± 0.47 |
| PUFA d | 17.65 ± 0.52 | 17.94 ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Xiao, Y.; Liu, T.; Yuan, M.; Liu, G.; Fang, J.; Yang, B. Exploration of Microalgal Species for Nutrient Removal from Anaerobically Digested Swine Wastewater and Potential Lipids Production. Microorganisms 2021, 9, 2469. https://doi.org/10.3390/microorganisms9122469
Chen Z, Xiao Y, Liu T, Yuan M, Liu G, Fang J, Yang B. Exploration of Microalgal Species for Nutrient Removal from Anaerobically Digested Swine Wastewater and Potential Lipids Production. Microorganisms. 2021; 9(12):2469. https://doi.org/10.3390/microorganisms9122469
Chicago/Turabian StyleChen, Zhihui, Yunhua Xiao, Tan Liu, Mingmin Yuan, Gang Liu, Jun Fang, and Bo Yang. 2021. "Exploration of Microalgal Species for Nutrient Removal from Anaerobically Digested Swine Wastewater and Potential Lipids Production" Microorganisms 9, no. 12: 2469. https://doi.org/10.3390/microorganisms9122469
APA StyleChen, Z., Xiao, Y., Liu, T., Yuan, M., Liu, G., Fang, J., & Yang, B. (2021). Exploration of Microalgal Species for Nutrient Removal from Anaerobically Digested Swine Wastewater and Potential Lipids Production. Microorganisms, 9(12), 2469. https://doi.org/10.3390/microorganisms9122469