
Modelling the Radial Growth of Geotrichum candidum: Effects of Temperature and Water Activity



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Experimental Design
2.3. Model Description
2.3.1. Primary Model
2.3.2. Secondary Modelling
2.3.3. Time Required to Achieve Visible Colonies
2.4. Statistical Analysis and Model Evaluation
3. Results and Discussion
3.1. Primary Surface Growth Modelling
3.2. Secondary Modelling
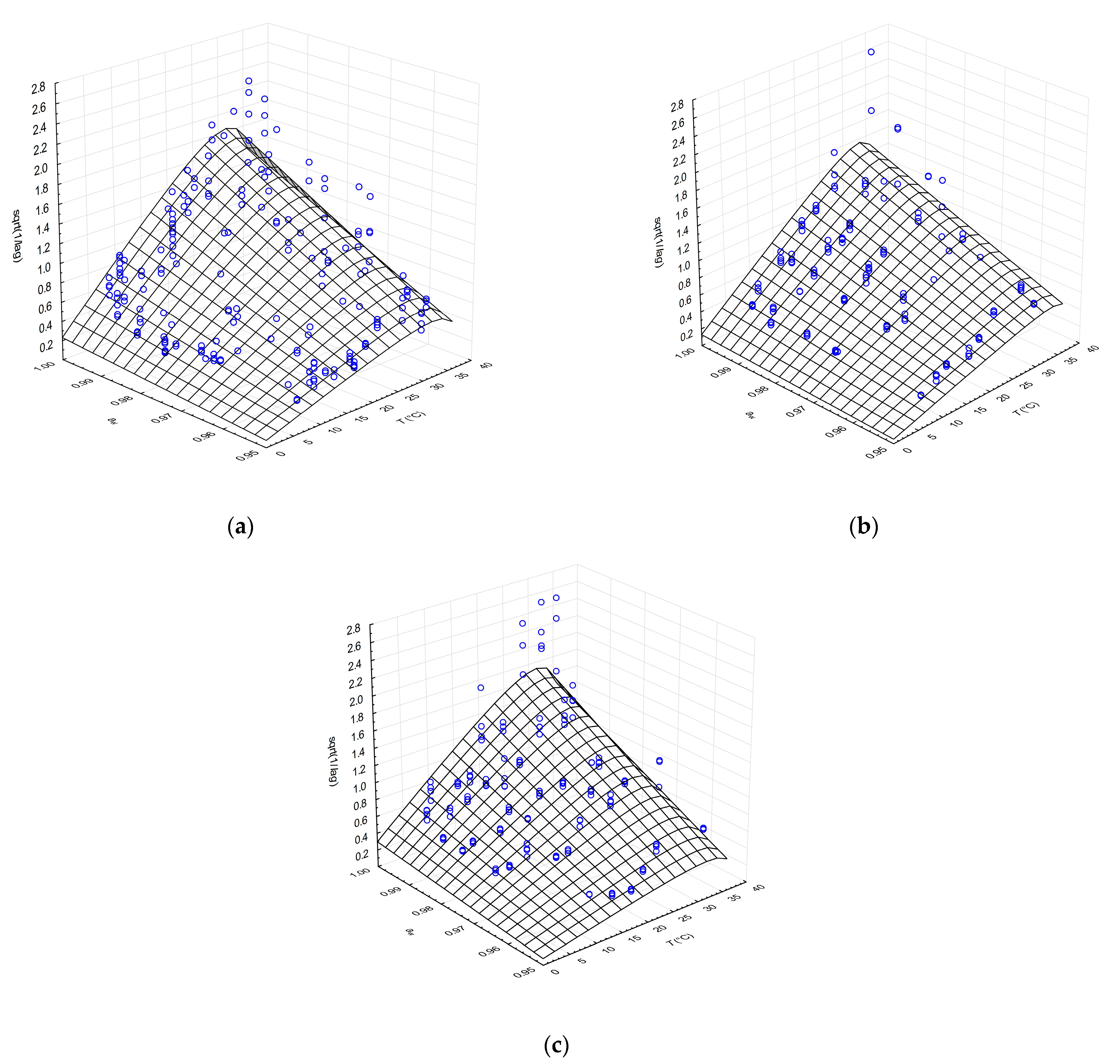
3.2.1. Combined Effects of aw Adjusted with NaCl and Temperature on Lag Time
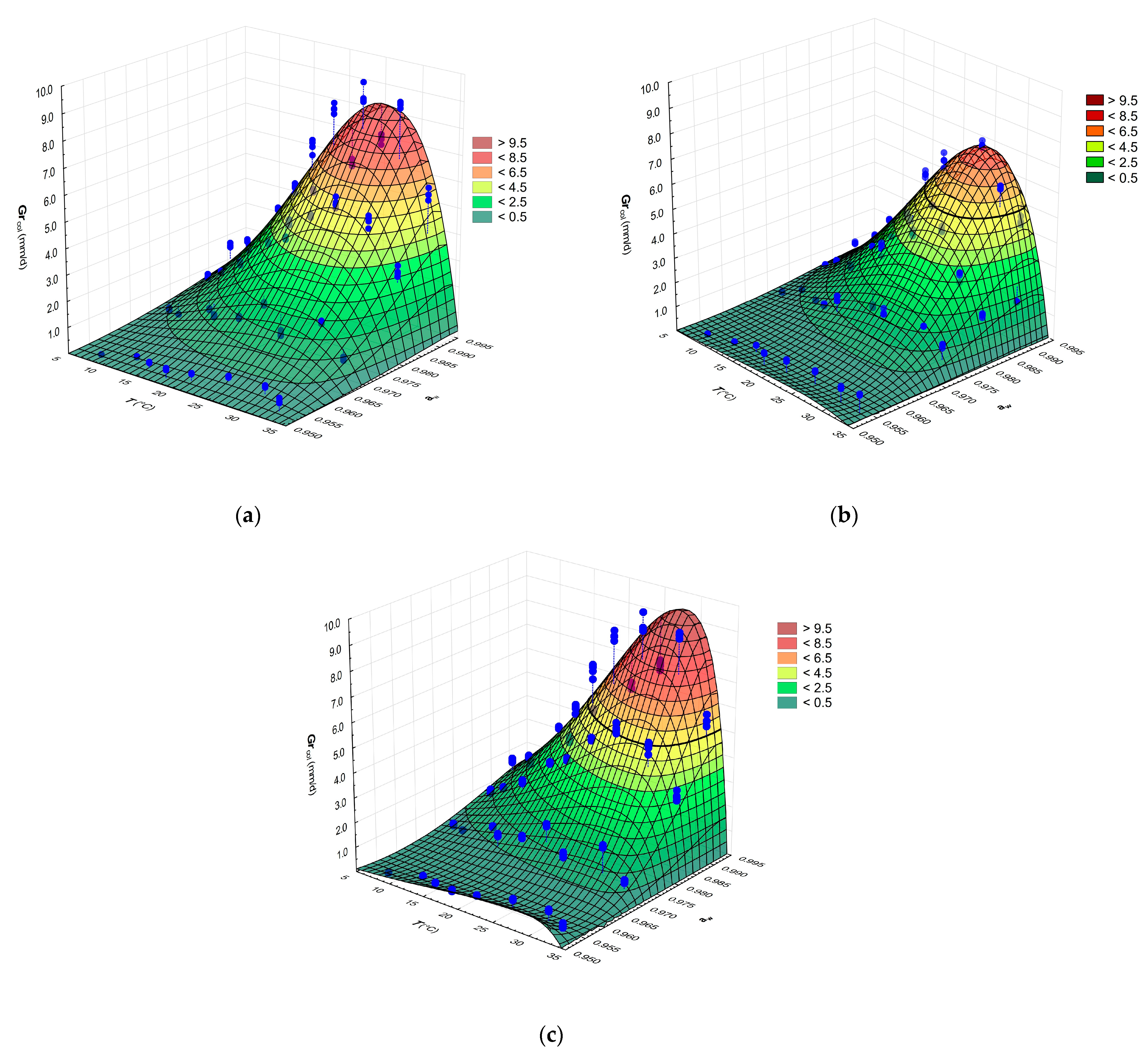
3.2.2. Combined Effects of aw and Temperature on the Growth of G. candidum
3.3. Prediction of the Time Required to Achieve Visible Colonies
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pitt, J.I.; Hocking, A.D. Fungi and Food Spoilage, 3rd ed.; Springer Science+Business Media: New York, NY, USA, 2009; pp. 53–142. [Google Scholar]
- Deák, T. Handbook of Food Spoilage Yeast, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 2–13. [Google Scholar]
- Hudecová, A.; Valík, Ľ.; Liptáková, D. Effect of lactic acid bacteria on growth dynamics of Geotrichum candidum during co-culture in milk. Slovak J. Food Sci. 2010, 4, 387–395. [Google Scholar]
- Laurenčík, M.; Sulo, P.; Sláviková, E.; Piecková, E.; Seman, M.; Ebringer, L. The diversity of eukaryotic microbiota in the traditional Slovak sheep cheese—Bryndza. Int. J. Food Microbiol. 2008, 127, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Boutrou, R.; Guéguen, M. Interests in Geotrichum candidum for cheese technology. Int. J. Food Microbiol. 2005, 102, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Koňuchová, M.; Lehotová, V.; Šípková, A.; Piecková, E.; Valík, Ľ. Biodiversity Evaluation of Geotrichum candidum Link. Is arthrosporic nucleus number in Geotrichum candidum related to the fungus biodiversity? Sci. Agric. Bohemica 2016, 47, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Kornacki, J.L.; Flowers, R.S.; Bradley, R.L. Microbiology of butter and related products. In Applied Dairy Microbiology, 2nd ed.; Marth, E.H., Steele, J.L., Eds.; Marcel Dekker: New York, NY, USA, 2001; pp. 127–150. [Google Scholar]
- Eliskases-Lechner, F.; Guéguen, M.; Panoff, J.M. Geotrichum candidum. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Fox, P.F., Mcsweeney, P.L.H., Eds.; Academic Press: London, UK, 2011; Volume 4, pp. 765–771. [Google Scholar]
- Aziza, M.; Couriol, C.; Amrane, A.; Boutrou, R. Evidences for synergistic effects of Geotrichum candidum on Penicillium camembertii growing on cheese juice. Enzym. Microb. Technol. 2005, 37, 218–224. [Google Scholar] [CrossRef]
- Boutrou, R.; Kerriou, L.; Gassi, J.-Y. Contribution of Geotrichum candidum to the proteolysis of soft cheese. Int. Dairy J. 2006, 16, 775–783. [Google Scholar] [CrossRef]
- Cogan, T.M.; Goerges, S.; Gelsomino, R.; Larpin, S.; Hohenegger, M.; Bora, N.; Jamet, E.; Rea, M.C.; Mounier, J.; Vancanneyt, M.; et al. Biodiversity of the surface microbial consortia from Limburger, Reblochon, Livarot, Tilsit, and Gubbeen cheeses. Microbiol. Spectrum. 2014, 2, 1–28. [Google Scholar]
- Desmasures, N. Mold-Ripened Varieties. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortello, M.-R., Eds.; Academic Press: Cambridge, UK, 2014; pp. 409–416. [Google Scholar]
- Grygier, A.; Majcher, M.; Myszka, K. Galactomyces geotrichum–moulds from dairy products with high biotechnological potential. Acta Sci. Pol. Technol. Aliment. 2017, 16, 5–16. [Google Scholar] [CrossRef]
- Hudecová, A.; Valík, Ľ.; Liptáková, D. Quantification of Geotrichum candidum growth in co-culture with lactic acid bacteria. Czech J. Food Sci. 2009, 27, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Šipošová, P.; Koňuchová, M.; Valík, Ľ.; Medveďová, A. Growth dynamics of lactic acid bacteria and dairy microscopic fungus Geotrichum candidum during their co-cultivation in milk. Food Sci. Technol. Int. 2020, in press. [Google Scholar]
- Bakry, A.M.; Campelo, P.H. Mini-review on functional characteristics of viili and manufacturing process. J. Food Biotechnol. Res. 2018, 2, 1–6. [Google Scholar]
- Witthuhn, R.C.; Schoeman, T.; Britz, T.J. Characterisation of the microbial population at different stages of Kefir production and Kefir grain mass cultivation. Int. Dairy J. 2005, 15, 383–389. [Google Scholar] [CrossRef]
- Kazou, M.; Grafakou, A.; Tsakalidou, E.; Georgalaki, M. Zooming into the microbiota of home-made and industrial kefir produced in Greece using classical microbiological and amplicon-based metagenomics analyses. Front. Microbiol. 2021, 12, 621069. [Google Scholar] [CrossRef] [PubMed]
- Dantigny, P.; Marín, S.; Beyer, M.; Magan, N. Mould germination: Data treatment and modelling. Int. J. Food Microbiol. 2007, 114, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Ramos, A.J.; Sanchis, V.; Marín, S. Predicting mycotoxins in foods: A review. Food Microbiol. 2009, 26, 757–769. [Google Scholar] [CrossRef]
- Koňuchová, M.; Valík, Ľ. Evaluation of radial growth dynamics variability of Geotrichum candidum: A quantitative approach. J. Food Nutr. Res. 2017, 56, 155–166. [Google Scholar]
- Baranyi, J.; Roberts, T.A. Mathematics of predictive food microbiology. Int. J. Food Microbiol. 1995, 26, 199–218. [Google Scholar] [CrossRef] [Green Version]
- Rosso, L.; Lobry, J.R.; Flandrois, J.P. An unexpected correlation between cardinal temperatures of microbial growth highlighted by a new model. J. Theor. Biol. 1993, 162, 447–463. [Google Scholar] [CrossRef]
- Zwietering, M.H.; Wijtzes, T.; De Wit, J.C.; van’T Riet, K. A decision support system for prediction of the microbial spoilage in foods. J. Food Protect. 1992, 55, 973–979. [Google Scholar] [CrossRef]
- Rosso, L.; Robinson, T.P. A cardinal model to describe the effect of water activity on the growth of moulds. Int. J. Food Microbiol. 2001, 63, 265–273. [Google Scholar] [CrossRef]
- Pérez-Rodríguez, F.; Valero, A. Predictive Microbiology in Foods, 5th ed.; Springer Science+Business Media: New York, NY, USA, 2013; pp. 25–55. [Google Scholar]
- Rosso, L.; Lobry, J.R.; Bajard, S.; Flandrois, J.P. Convenient model to describe the combined effects of temperature and pH on microbial growth. Appl. Environ. Microbiol. 1995, 61, 610–616. [Google Scholar] [CrossRef] [Green Version]
- Gougouli, M.; Koutsoumanis, K.P. Modelling growth of Penicillium expansum and Aspergillus niger at constant and fluctuating temperature conditions. Int. J. Food Microbiol. 2010, 140, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Greco, M.; Pardo, A.; Pose, G.; Patriarca, A. Effect of water activity and temperature on the growth of Eurotium species isolated from animal feeds. Rev. Iberoam. Micol. 2018, 35, 39–48. [Google Scholar] [CrossRef]
- Dagnas, S.; Gougouli, M.; Onno, B.; Koutsoumanis, K.P.; Membré, J.-M. Quantifying the effect of water activity and storage temperature on single spore lag times of three moulds isolated from spoiled bakery products. Int. J. Food Microbiol. 2017, 240, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Huchet, V.; Pavan, S.; Lochardet, A.; Divanac’h, M.L.; Postollec, F.; Thuault, D. Development and application of a predictive model of Aspergillus candidus growth as a tool to improve shelf life of bakery products. Food Microbiol. 2013, 36, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Astoreca, A.; Vaamonde, G.; Dalcero, A.; Ramos, A.J.; Marín, S. Modelling the effect of temperature and water activity of Aspergillus flavus isolates from corn. Int. J. Food Microbiol. 2012, 156, 60–67. [Google Scholar] [CrossRef]
- Samapundo, S.; Devlieghere, F.; Geeraerd, A.H.; De Meulenaer, B.; Van Impe, J.F.; Debevere, J. Modelling of the individual and combined effects of water activity and temperature on the radial growth of Aspergillus flavus and A. parasiticus on corn. Food Microbiol. 2007, 24, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Panagou, E.Z.; Chelonas, S.; Chatzipavlidis, I.; Nychas, G.-J.E. Modelling the effect of temperature and water activity on the growth rate and growth/no growth interface of Byssochlamys fulva and Byssochlamys nivea. Food Microbiol. 2010, 27, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Nevarez, L.; Vasseur, V.; Le Madec, A.; Le Bras, M.; Coroller, L.; Leguérinel, I.; Barbier, G. Physiological traits of Penicillium glabrum strain LCP 08.5568, a filamentous fungus isolated from bottled aromatised mineral water. Int. J. Food Microbiol. 2009, 130, 166–171. [Google Scholar] [CrossRef] [Green Version]
- Baert, K.; Valero, A.; De Meulenaer, B.; Samapundo, S.; Ahmed, M.M.; Bo, L.; Debevere, J.; Devlieghere, F. Modelling the effect of temperature on the growth rate and lag phase of Penicillium expansum in apples. Int. J. Food Microbiol. 2007, 118, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Parra, R.; Magan, N. Modelling the effect of temperature and water activity on growth of Aspergillus niger strains and applications for food spoilage moulds. J. Appl. Microbiol. 2004, 97, 429–438. [Google Scholar] [CrossRef]
- Sautour, M.; Rouget, A.; Dantigny, P.; Divies, C.; Bensoussan, M. Application of Doehlert design to determine the combined effects of temperature, water activity and pH on conidial germination of Penicillium chrysogenum. Int. J. Food Microbiol. 2001, 91, 900–906. [Google Scholar] [CrossRef] [Green Version]
- Marín, S.; Cuevas, D.; Ramos, A.J.; Sanchis, V. Fitting of colony diameter and ergosterol as indicators of food borne mould growth to known growth models in solid medium. Int. J. Food Microbiol. 2008, 121, 139–149. [Google Scholar] [CrossRef]
- Sant’ana, A.; Franco, B.D.G.M.; Schaffner, D.W. Modeling the growth rate and lag time of different strains of Salmonella enterica and Listeria monocytogenes in ready-to-eat lettuce. Food Microbiol. 2012, 30, 267–273. [Google Scholar] [CrossRef]
- Valík, Ľ.; Medveďová, A.; Liptáková, D. Characterization of the growth of Lactobacillus rhamnosus GG in milk at suboptimal temperatures. J. Food Nutr. Res. 2008, 47, 60–67. [Google Scholar]
- Gougouli, M.; Kalantzi, K.; Beletsiotis, E.; Koutsoumanis, K.P. Development and application of predictive models for fungal growth as tools to improve quality control in yogurt production. Food Microbiol. 2011, 28, 1453–1462. [Google Scholar] [CrossRef]
- Marín, S.; Colom, C.; Sanchis, V.; Ramos, A.J. Modelling of growth of aflatoxigenic A. flavus isolates from red chilli powder as a function of water availability. Int. J. Food Microbiol. 2009, 128, 491–496. [Google Scholar] [CrossRef]
- Dagnas, S.; Onno, B.; Membré, J.M. Modeling growth of three bakery product spoilage molds as a function of water activity, temperature and pH. Int. J. Food Microbiol. 2014, 186, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Belbahi, A.; Leguerinel, I.; Méot, J.-M.; Loiseau, G.; Madani, K.; Bohuon, P. Modelling the effect of temperature, water activity and carbon dioxide on the growth of Aspergillus niger and Alternaria alternata isolated from fresh date fruit. J. Appl. Microbiol. 2016, 121, 1685–1698. [Google Scholar] [CrossRef] [PubMed]
- Samapundo, S.; Devlieghere, F.; De Meulenaer, B.; Debevere, J. Growth kinetics of cultures from single spores of Aspergillus flavus and Fusarium verticillioides on yellow dent corn meal. Food Microbiol. 2007, 24, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Hudecová, A.; Valík, Ľ.; Liptáková, D. Influence of temperature on the surface growth of Geotrichum candidum. Acta Chim. Slovaca. 2009, 2, 75–87. [Google Scholar]
- Hudecová, A.; Valík, Ľ.; Liptáková, D. Surface growth of Geotrichum candidum: Effect of the environmental factors on its dynamics. Slovak J. Food Sci. 2011, 5, 17–22. [Google Scholar] [CrossRef]
- Domsch, K.H.; Gams, W.; Anderson, T.-H. Compendium of Soil Fungi, 1st ed.; Academic Press: London, UK, 1980; Volume 2, pp. 1–859. [Google Scholar]
- Šípková, A.; Valík, L.; Čižniar, M.; Liptáková, D. Characterization of mutual relations between Geotrichum candidum and Lactobacillus rhamnosus GG in milk: A quantitative approach. Food Sci. Technol. Int. 2014, 20, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Tornadijo, M.E.; Fresno, J.M.; Sarmiento, R.M.; Carballo, J. Study of the yeasts during the ripening process of Armada cheeses from raw goat’s milk. Le Lait 1998, 78, 647–659. [Google Scholar] [CrossRef] [Green Version]
- Plaza, P.; Usall, J.; Teixidó, N.; Vièas, I. Effect of water activity and temperature on germination and growth of Penicillium digitatum, P. italicum and Geotrichum candidum. J. Appl. Microbiol. 2003, 94, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Marcellino, S.N.; Benson, D.R. The good, the bad, and the ugly: Tales of mold-ripened cheese. In Cheese and Microbes, 1st ed.; Donnelly, C.W., Ed.; ASM Press: Washington, DC, USA, 2014; pp. 95–132. [Google Scholar]
- Tempel, T.; Nielsen, M.S. Effects of atmospheric conditions, NaCl and pH on growth and interactions between moulds and yeasts related to blue cheese production. Int. J. Food Microbiol. 2000, 57, 193–199. [Google Scholar] [CrossRef]
- Uraz, T.; Özer, B.H. Molds Employed in Food Processing. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortello, M.-R., Eds.; Academic Press: Cambridge, UK, 2014; pp. 522–528. [Google Scholar]
- Dantigny, P. Applications of predictive modeling techniques to fungal growth in foods. Curr. Opin. Food Sci. 2021, 38, 86–90. [Google Scholar] [CrossRef]
- Aldars-García, L.; Sanchis, V.; Ramos, A.J.; Marín, S. Time-course of germination, initiation of mycelium proliferation and probability of visible growth and detectable AFB1 production of an isolate of Aspergillus flavus on pistachio extract agar. Food Microbiol. 2017, 64, 104–111. [Google Scholar] [CrossRef]
- Burgain, A.; Bensoussan, M.; Dantigny, P. Effect of inoculum size and water activity on the time to visible growth of Penicillium chrysogenum colony. Int. J. Food Microbiol. 2013, 163, 180–183. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameters | Isolate G | Isolate I | Strain CBS 577.83 |
|---|---|---|---|
| 1/λopt (1/d) | 3.30 ± 0.25 | 3.64 ± 0.68 | 2.97 ± 1.06 |
| Tmin (°C) * | −5.15 ± 0.10 | −5.45 ± 0.14 | −1.95 ± 0.03 |
| Topt (°C) | 33.13 ± 0.32 | 34.56 ± 0.70 | 35.83 ± 0.94 |
| Tmax (°C) | 37.35 ± 0.09 | 37.09 ± 0.11 | 37.06 ± 0.27 |
| awmin | 0.927 ± 0.003 | 0.938 ± 0.003 | 0.925 ± 0.003 |
| awopt | 0.998 (fixed value) | 0.998 (fixed value) | 0.997 (fixed value) |
| awmax | 0.999 (fixed value) | 0.999 (fixed value) | 1 (fixed value) |
| RMSE | 0.268 | 0.280 | 0.173 |
| %MRE | 9.5 | 7.6 | 4.9 |
| %SEP | 32.0 | 32.4 | 23.3 |
| n | 187 | 159 | 152 |
| R2 | 0.707 | 0.685 | 0.808 |
| Parameters | Isolate G | Isolate I | Strain CBS 577.83 |
|---|---|---|---|
| RGRopt (mm/d) | 7.85 ± 0.15 | 6.87 ± 0.15 | 9.13 ± 0.20 |
| Tmin (°C) * | −1.46 ± 0.01 | −0.43 ± 0.01 | −5.17 ± 0.05 |
| Topt (°C) | 25.92 ± 0.04 | 25.41 ± 0.04 | 28.03 ± 0.37 |
| Tmax (°C) | 35.63 ± 0.07 | 34.17 ± 0.38 | 37.57 ± 0.06 |
| awmin | 0.9479 ± 0.0014 | 0.9557 ± 0.0016 | 0.9591 ± 0.0012 |
| awopt | 0.9934 ± 0.0003 | 0.9919 ± 0.0003 | 0.9916 ± 0.0002 |
| awmax | 0.9977 ± 0.0010 | 0.9988 ± 0.0011 | 0.9991 ± 0.0001 |
| RMSE | 0.415 | 0.405 | 0.278 |
| %MRE | 4.2 | 4.3 | 4.4 |
| %SEP | 16.4 | 20.0 | 19.7 |
| n | 192 | 184 | 188 |
| R2 | 0.985 | 0.954 | 0.980 |
| Temperature (°C) | % NaCl | t3 (d) | RMSE for the t3 Predictions Based on Both Cultures Data (n = 6) | |
|---|---|---|---|---|
| Strain CBS 557.83 | Isolate G | |||
| 4 | 0.995 | 8.1 ± 0.3 | 9.6 ± 0.4 | 0.9 |
| 0.99 | 8.6 ± 0.5 | 10.6 ± 0.2 | 1.1 | |
| 0.98 | 13.8 ± 0.6 | 15.9 ± 0.05 | 1.2 | |
| 0.97 | 32.3 ± 1.9 | 28.9 ± 0.9 | 2.4 | |
| 5 | 0.995 | 6.6 ± 0.3 | 7.4 ± 0.3 | 0.5 |
| 0.99 | 7.1 ± 0.4 | 8.0 ± 0.1 | 0.6 | |
| 0.98 | 11.3 ± 0.5 | 12.9 ± 0.04 | 0.6 | |
| 0.97 | 26.4 ± 1.5 | 21.9 ± 0.7 | 2.8 | |
| 6 | 0.995 | 5.5 ± 0.2 | 5.9 ± 0.2 | 0.3 |
| 0.99 | 5.9 ± 0.3 | 6.5 ± 0.1 | 0.4 | |
| 0.98 | 9.4 ± 0.4 | 9.7 ± 0.04 | 0.3 | |
| 0.97 | 22.0 ± 1.3 | 17.3 ± 0.5 | 2.8 | |
| 7 | 0.995 | 4.7 ± 0.2 | 4.8 ± 0.2 | 0.2 |
| 0.99 | 5.0 ± 0.3 | 5.3 ± 0.1 | 0.2 | |
| 0.98 | 7.9 ± 0.4 | 7.9 ± 0.4 | 0.2 | |
| 0.97 | 18.6 ± 1.1 | 14.1 ± 0.4 | 2.7 | |
| 8 | 0.995 | 4.0 ± 0.2 | 4.0 ± 0.2 | 0.1 |
| 0.99 | 4.3 ± 0.3 | 4.4 ± 0.1 | 0.2 | |
| 0.98 | 6.8 ± 0.3 | 6.6 ± 0.04 | 0.2 | |
| 0.97 | 15.9 ± 1.0 | 11.7 ± 0.3 | 2.5 | |
| 9 | 0.995 | 3.5 ± 0.1 | 3.4 ± 0.1 | 0.1 |
| 0.99 | 3.7 ± 0.2 | 3.8 ± 0.1 | 0.2 | |
| 0.98 | 5.9 ± 0.3 | 5.6 ± 0.04 | 0.2 | |
| 0.97 | 13.8 ± 0.8 | 9.9 ± 0.3 | 2.3 | |
| 10 | 0.995 | 3.0 ± 0.1 | 2.9 ± 0.1 | 0.1 |
| 0.99 | 3.2 ± 0.2 | 3.3 ± 0.1 | 0.1 | |
| 0.98 | 5.2 ± 0.2 | 4.9 ± 0.04 | 0.2 | |
| 0.97 | 12.1 ± 0.7 | 8.6 ± 0.2 | 2.1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koňuchová, M.; Valík, Ľ. Modelling the Radial Growth of Geotrichum candidum: Effects of Temperature and Water Activity. Microorganisms 2021, 9, 532. https://doi.org/10.3390/microorganisms9030532
Koňuchová M, Valík Ľ. Modelling the Radial Growth of Geotrichum candidum: Effects of Temperature and Water Activity. Microorganisms. 2021; 9(3):532. https://doi.org/10.3390/microorganisms9030532
Chicago/Turabian StyleKoňuchová, Martina, and Ľubomír Valík. 2021. "Modelling the Radial Growth of Geotrichum candidum: Effects of Temperature and Water Activity" Microorganisms 9, no. 3: 532. https://doi.org/10.3390/microorganisms9030532
APA StyleKoňuchová, M., & Valík, Ľ. (2021). Modelling the Radial Growth of Geotrichum candidum: Effects of Temperature and Water Activity. Microorganisms, 9(3), 532. https://doi.org/10.3390/microorganisms9030532