
In Vitro Modulatory Effect of Stevioside, as a Partial Sugar Replacer in Sweeteners, on Human Child Microbiota



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Fermentation Model
2.1.1. Fecal Samples
2.1.2. Simulation Conditions
2.2. Determination of Microbiota Pattern
2.3. Determination of Metabolomic Response
2.4. Determination of Antioxidant Response
2.5. Statistical Analysis
3. Results
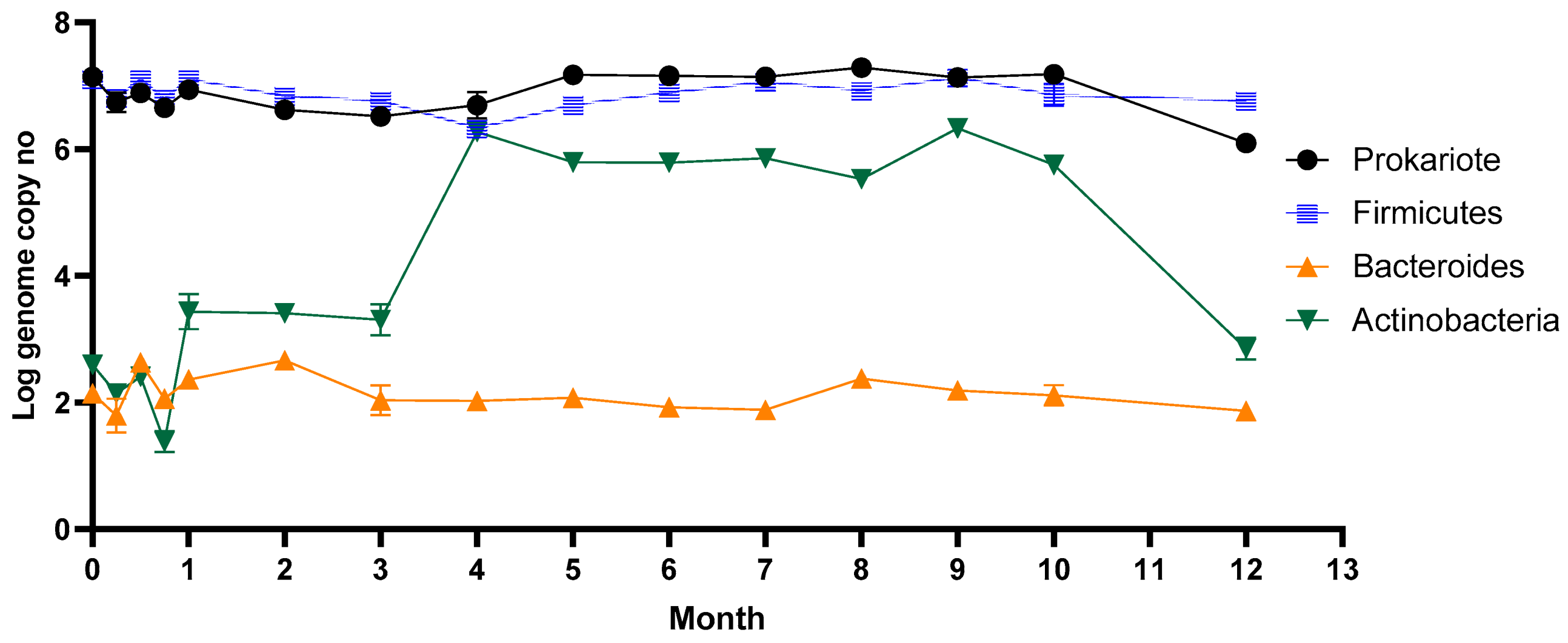
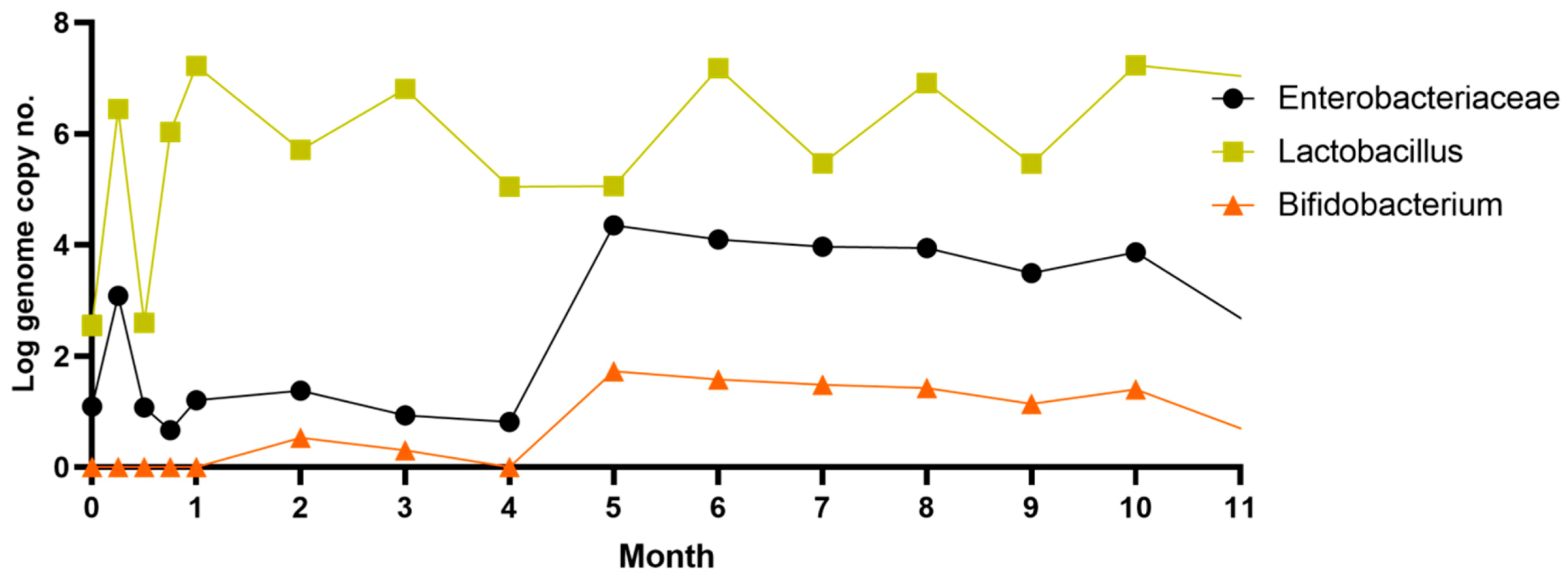
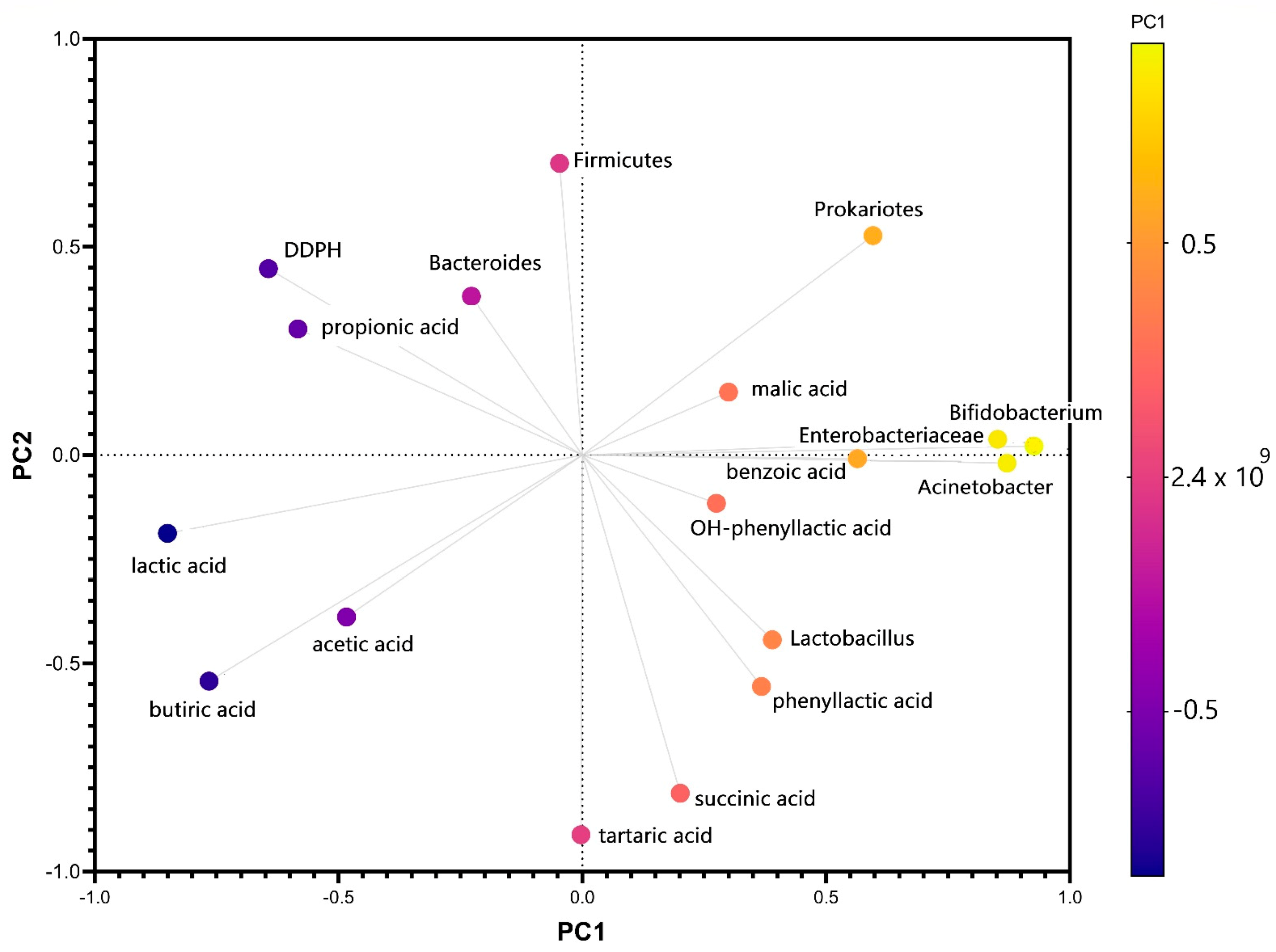
3.1. The Response of the Microbiota Pattern after Stevia In Vitro Consumption
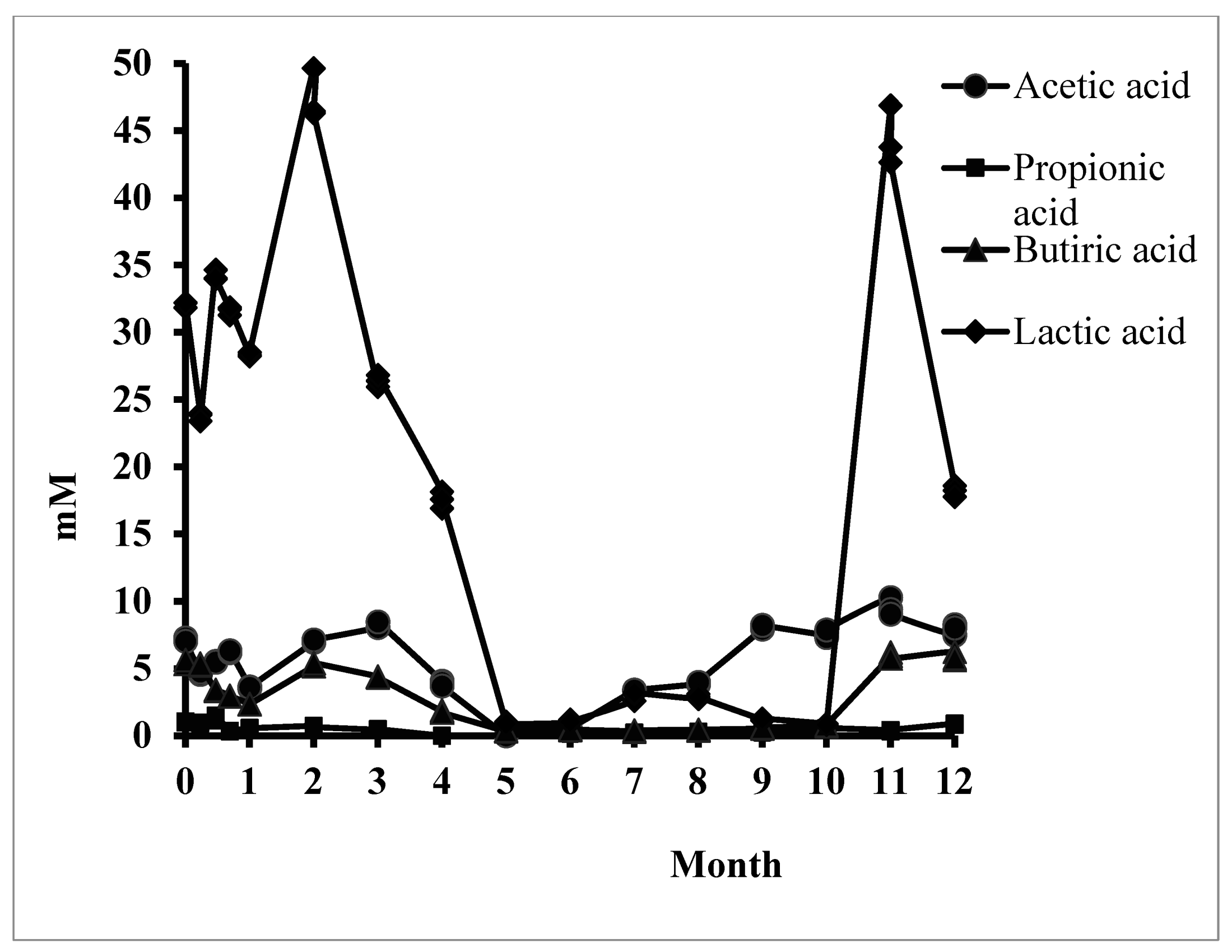
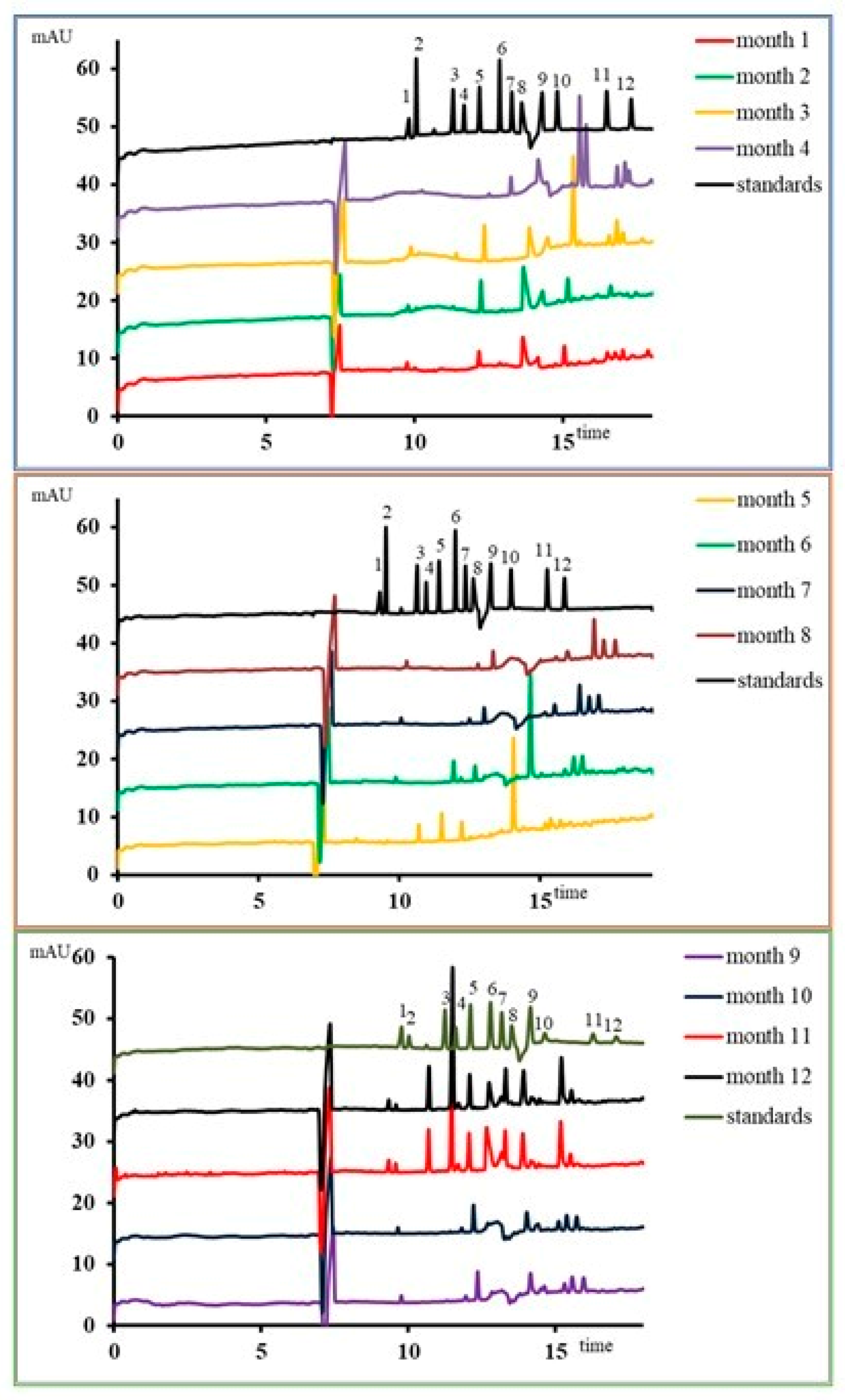
3.2. The Response of the Metabolomic Pattern after Stevia In Vitro Consumption
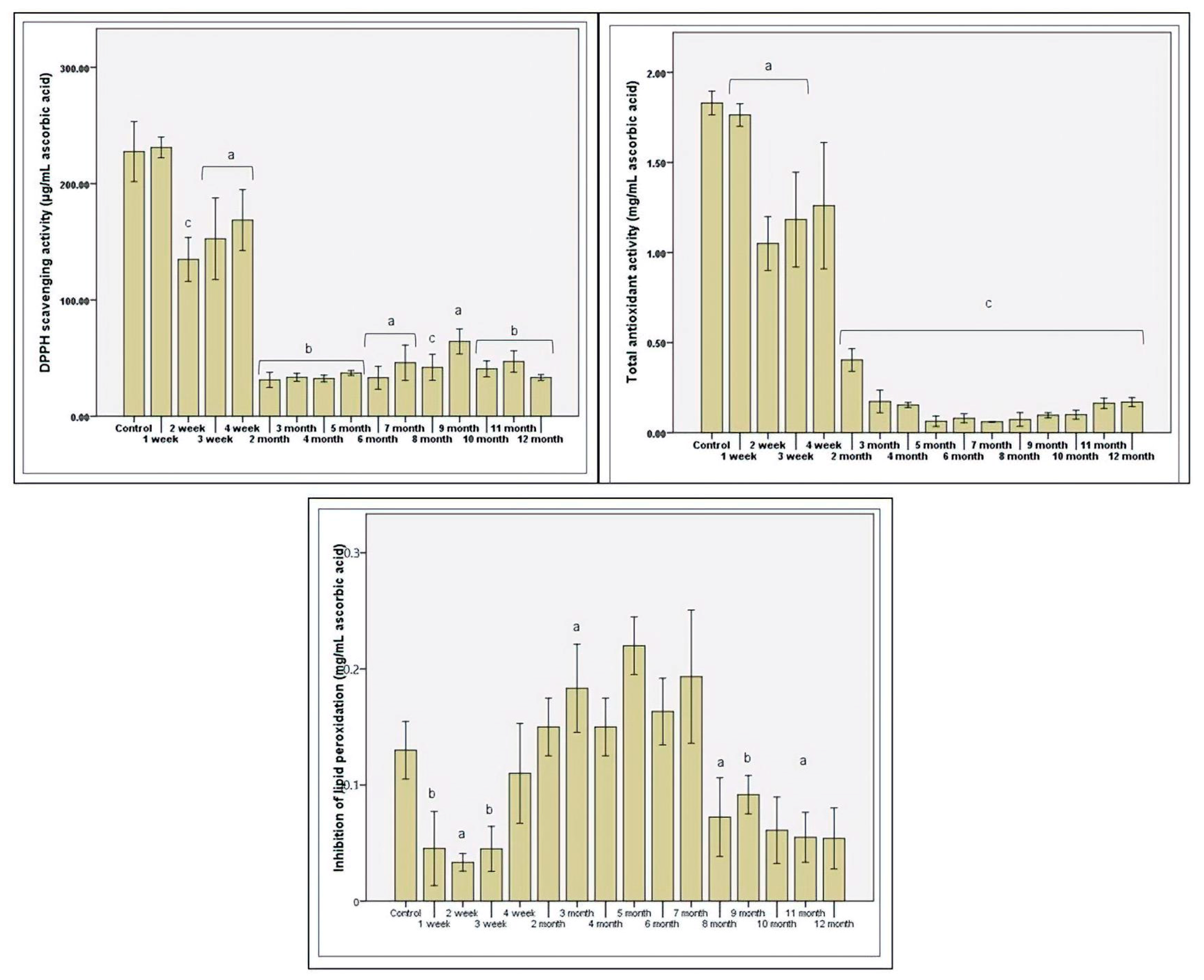
3.3. Antioxidant Status after Stevia In Vitro Consumption
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kho, Z.Y.; Lal, S.K. The Human Gut Microbiome—A Potential Controller of Wellness and Disease. Front. Microbiol. 2018, 9, 1835. [Google Scholar] [CrossRef] [Green Version]
- Sommer, F.; Anderson, J.; Bharti, R.; Raes, J.; Rosenstiel, P. The resilience of the intestinal microbiota influences health and disease. Nat. Rev. Microbiol. 2017, 15, 630–638. [Google Scholar] [CrossRef] [PubMed]
- Srikantha, P.; Mohajeri, M.H. The Possible Role of the Microbiota-Gut-Brain-Axis in Autism Spectrum Disorder. Int. J. Mol. Sci. 2019, 20, 2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, M.D.; Goossens, G.H.; Blaak, E.E. The Impact of Artificial Sweetenerson Body Weight Control and Glucose Homeostasis. Front. Nutr. 2021, 7, 598340. [Google Scholar] [CrossRef]
- Saraiva, A.; Carrascosa, C.; Dele Raheem, D.; Fernando Ramos, F.; Raposo, A. Natural Sweeteners: The Relevance of Food Naturalness for Consumers, Food Security Aspects, Sustainability and Health Impacts. Int. J. Environ. Res. Public Health 2020, 17, 6285. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, H.; Wang, Y.; Lau, H.; Zhou, W.; Chen, C.; Tan, S. A review of stevia as a potential healthcare product: Up-to-date functional characteristics, administrative standards and engineering techniques. Trends Food Sci. Technol. 2020, 103, 264–281. [Google Scholar] [CrossRef]
- Lemusmondaca, R.; Vegagalvez, A.; Rojas, P.; Stucken, K.; Delporte, C.; Valenzuela Barra, G.; Jagus, R.J.; Agüero, M.V.; Pasten, A. Antioxidant, antimicrobial and anti-inflammatory potential of Stevia rebaudiana leaves: Effect of different drying methods. J. Appl. Res. Med. Arom. Plants 2018, 11, 37–46. [Google Scholar] [CrossRef]
- Brijesh, K.; Kamath, M. Experimental evaluation of anti-hyperglycemic and hypolipidemic effects of stevia rebaudiana, Anacardium occidentale on wistar rats. Int. J. Basic Clin. Pharmacol. 2016, 5, 2463–2467. [Google Scholar] [CrossRef] [Green Version]
- Panagiotou, C.; Mihailidou, C.; Brauhli, G.; Katsarou, O.; Moutsatsou, P. Effect of steviol, steviol glycosides and stevia extract on glucocorticoid receptor signaling in normal and cancer blood cells. Mol. Cell. Endocrin. 2018, 460, 189–199. [Google Scholar] [CrossRef]
- Rashad, N.M.; Abdelsamad, M.A.E.; Amer, A.M.; Sitohy, M.Z.; Mousa, M.M. The impact of stevioside supplementation on glycemic control and lipid profile in patients with type 2 diabetes: A controlled clinical trial. Egypt. J. Int. Med. 2019, 31, 22–30. [Google Scholar] [CrossRef]
- Available online: https://www.efsa.europa.eu/en/events/event/technical-stakeholder-event-re-evaluation-authorised-food-additives (accessed on 2 January 2021).
- Singh, D.P.; Kumari, M.; Prakash, H.G.G.P.; Solomon, S. Phytochemical and Pharmacological Importance of Stevia: A Calorie-Free Natural Sweetener. Sugar Tech. 2019, 21, 227–234. [Google Scholar] [CrossRef]
- Bundgaard, A.C.C.; Rafiq, S.; Jeppesen, P.B. Effect of Steviol Glycosides on Human Health with Emphasis on Type 2 Diabetic Biomarkers: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2019, 11, 1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bueno-Hernández, N.; Vázquez-Frías, R.; Abreu, A.T.; Almeda-Valdés, P.; Barajas-Nava, L.A.; Carmona-Sánchez, R.I.; Chávez-Sáenz, J.; Consuelo-Sánchez, A.; Espinosa-Flores, A.J.; Hernández-Rosiles, V.; et al. Review of the scientific evidence and technical opinion onnoncaloric sweetener consumption in gastrointestinal diseases. J. Gastroenterol. Mex. 2019, 84, 492–510. [Google Scholar] [CrossRef]
- Verduci, E.; Carbone, M.T.; Borghi, E.; Ottaviano, E.; Burlina, A.; Biasucci, G. Nutrition, Microbiota and Role of Gut-Brain Axis in Subjects with Phenylketonuria (PKU): A Review. Nutrients 2020, 12, 3319. [Google Scholar] [CrossRef]
- Liauchonak, I.; Qorri, B.; Dawoud, F.; Riat, Y.; Szewczuk, M.R. Non-Nutritive Sweeteners and Their Implications on the Development of Metabolic Syndrome. Nutrients 2019, 11, 644. [Google Scholar] [CrossRef] [Green Version]
- Majzoobi, M.; Mohammadi, M.; Farahnaky, A. Simultaneous reduction of fat and sugar in cake production; effects of changing sucrose, oil, water, inulin, and Rebaudioside A on cake batter properties. J. Food Process. Preserv. 2020, 44, 10. [Google Scholar] [CrossRef]
- Urban, J.D.; Carakostas, M.C.; Taylor, S.L. Stevioside glycoside safety: Are highly purified Stevioside glycoside sweeteners food allergens? Food Chem. Toxicol. 2015, 71–78. [Google Scholar] [CrossRef]
- De Carvalhoa, N.M.; Waltonb, G.E.; Povedab, C.G.; Silvaa, S.N.; Amorima, M.; Madureiraa, A.R.; Pintadoa, M.E.; Gibsonb, G.R.; Jauregi, P. Study of in vitro digestion of Tenebrio molitor flour for evaluation of its impact on the human gut microbiota. J. Funct. Foods 2019, 59, 101–109. [Google Scholar] [CrossRef]
- Tsitko, I.; Wiik-Miettinen, F.; Mattila, O.; Rosa-Sibakov, N.; Seppänen-Laakso, T.; Maukonen, J.; Nordlund, E.; Saarela, M. A Small In Vitro Fermentation Model for Screening the Gut Microbiota Effects of Different Fiber Preparations. Int. J. Mol. Sci. 2019, 20, 1925. [Google Scholar] [CrossRef] [Green Version]
- Okumura, R.; Takeda, K. Roles of intestinal epithelial cells in the maintenance of gut homeostasis. Exp. Mol. Med. 2017, 49, e338. [Google Scholar] [CrossRef] [Green Version]
- Alexeev, E.E.; Lanis, J.M.; Kao, D.J.; Campbell, E.L.; Kelly, C.J.; Battista, K.D.; Gerich, M.E.; Jenkins, B.R.; Walk, S.T.; Kominsky, D.J.; et al. Microbiota-Derived Indole Metabolites Promote Human and Murine Intestinal Homeostasis through Regulation of Interleukin-10 Receptor. Am. J. Pathol. 2018, 188, 5. [Google Scholar] [CrossRef] [Green Version]
- Piotr Konopelski, P.; Ufnal, M. Indoles—Gut Bacteria Metabolites of Tryptophan with Pharmacotherapeutic Potential. Curr. Drug Metab. 2018, 19, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Powell, D.N.; Swimm, A.; Sonowal, R.; Bretin, A.; Gewirtz, A.T.; Jones, R.M.; Kalman, D. Indoles from the commensal microbiota act via the AHR and IL-10 to tune the cellular composition of the colonic epithelium during aging. Proc. Natl. Acad. Sci. USA 2020, 117, 21519–21526. [Google Scholar] [CrossRef] [PubMed]
- Hendrikx, T.; Schnabl, B. Indoles: Metabolites produced by intestinal bacteria capable of controlling liver disease manifestation. J. Int. Med. 2019, 286, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Heinken, A.; Ravcheev, D.A.; Baldini, F.; Heirendt, L.; Fleming, R.M.T.; Thiele, I. Systematic assessment of secondary bile acid metabolism in gut microbes reveals distinct metabolic capabilities in inflammatory bowel disease. Microbiome 2019, 7, 75. [Google Scholar] [CrossRef]
- Winston, J.A.; Theriot, C.M. Diversification of host bile acids by members of the gut microbio-ta. Gut Microbes 2020, 11, 158–171. [Google Scholar] [CrossRef] [PubMed]
- Van Best, N.; Rolle-Kampczyk, U.; Schaap, F.G.; Basic, M.; Olde Damink, S.W.M.; Bleich, A.; Savelkoul, P.H.M.; von Bergen, M.; Penders, J.; Hornef, M.W. Bile acids drive the newborn’s gut microbiota maturation. Nat. Commun. 2020, 11, 3692. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.L.; Shu, C.C.; Chen, Y.M.; Lu, J.J.; Wu, T.S.; Lai, W.F.; Tzeng, C.M.; Lai, H.C.; Lu, C.C. Like Cures Like: Pharmacological Activity of Anti-Inflammatory Lipopolysaccharides from Gut Microbiome. Front. Pharmacol. 2020, 11, 554. [Google Scholar] [CrossRef] [PubMed]
- d’Hennezel, E.; Abubucker, S.; Murphy, L.O.; Cullen, T.W. Total lipopolysaccharide from the human gut microbiome silences Toll-like receptor signaling. mSystems 2017, 2, e00046-17. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, M.C.; Lankelma, J.M.; Wolff, N.S.; Hugenholtz, F.; de Vos, A.F.; van der Poll, T.; Joost Wiersinga, W. Effect of antibiotic gut microbiota disruption on LPS-induced acute lung inflammation. PLoS ONE 2020, 15, e0241748. [Google Scholar] [CrossRef]
- Vamanu, E.; Pelinescu, D.; Gatea, F.; Sârbu, I. Altered in Vitro Metabolomic Response of the Human Microbiota to Sweeteners. Genes 2019, 10, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: https://www.efsa.europa.eu/en/press/news/ans100414 (accessed on 1 January 2020).
- Vamanu, E.; Ene, M.; Biță, B.; Ionescu, C.; Crăciun, L.; Sârbu, I. In Vitro Human Microbiota Response to Exposure to Silver Nanoparticles Biosynthesized with Mushroom Extract. Nutrients 2018, 10, 607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatea, F.; Teodor, E.D.; Paun, G.; Matei, A.O.; Radu, G.L. Capillary Electrophoresis Method Validation for Organic Acids Assessment in Probiotics. Food Anal. Meth. 2015, 8, 1335–1340. [Google Scholar] [CrossRef]
- Zhang, J.; Zhao, X.; Jiang, Y.; Zhao, W.; Guo, T.; Cao, Y.; Teng, J.; Hao, X.; Zhao, J.; Yang, Z. Antioxidant status and gut microbiota change in an aging mouse model as influenced by exopolysaccharide produced by Lactobacillus plantarum YW11 isolated from Tibetan kefir. J. Dairy Sci. 2017, 8, 6025–6041. [Google Scholar] [CrossRef] [PubMed]
- Coman, M.M.; Oancea, A.M.; Verdenelli, M.C.; Cecchini, C.; Bahrim, G.E.; Orpianesi, C.; Cresci, A.; Silvi, S. Polyphenol content and in vitro evaluation of antioxidant, antimicrobial and prebiotic properties of red fruit extracts. Eur. Food Res. Technol. 2018, 244, 735–745. [Google Scholar] [CrossRef]
- Wang, J.; Yang, Z.; Celi, P.; Yan, L.; Ding, X.; Bai, S.; Zeng, Q.; Mao, X.; Feng, B.; Xu, S.; et al. Alteration of the Antioxidant Capacity and Gut Microbiota under High Levels of Molybdenum and Green Tea Polyphenols in Laying Hens. Antioxidants 2019, 8, 503. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Chen, T.; Dong, S.; Xiong, Y.; Hua, W.; Xu, F. The effects of rebaudioside A onmicrobial diversity in mouse intestine. Food Sci. Technol. Res. 2014, 20, 459–467. [Google Scholar] [CrossRef] [Green Version]
- Cho, E.J.; Leem, S.; Kim, S.A.; Yang, J.; Lee, Y.B.; Kim, S.S.; Cheong, J.Y.; Cho, S.W.; Kim, J.W.; Kim, S.M.; et al. Circulating Microbiota-Based Metagenomic Signature for Detection of Hepatocellular Carcinoma. Sci. Rep. 2019, 9, 7536. [Google Scholar] [CrossRef]
- Filippone, A.; Lanza, M.; Campolo, M.; Casili, G.; Paterniti, I.; Cuzzocrea, S.; Esposito, E. The Anti-Inflammatory and Antioxidant Effects of Sodium Propionate. Int. J. Mol. Sci. 2020, 21, 3026. [Google Scholar] [CrossRef]
- Olivier-Van Stichelen, S.; Rother, K.I.; Hanover, J.A. Maternal Exposure to Non-nutritive Sweeteners Impacts Progeny’s Metabolism and Microbiome. Front. Microbiol. 2019, 10, 1360. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, S.; Maikowski, U.; Blankenstein, O.; Biebermann, H.; Tarnow, P.; Grüters, A. Type 2 diabetes and impaired glucose tolerance in European children and adolescents with obesity—A problem that is no longer restricted to minority groups. Eur. J. Endocrinol. 2004, 151, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Gardana, C.; Simonetti, P.; Canzi, E.; Zanchi, R.; Pietta, P. Metabolims of stevioside and rebaudioside A from Stevia rebaudiana extracts by human microflora. J. Agric. Food Chem. 2003, 51, 6618–6622. [Google Scholar] [CrossRef]
- Markus, V.; Share, O.; Teralı, K.; Ozer, N.; Marks, R.S.; Kushmaro, A.; Golberg, K. Anti-Quorum Sensing Activity of Stevia Extract, Stevioside, Rebaudioside A and Their Aglycon Stevioside. Molecules 2020, 25, 5480. [Google Scholar] [CrossRef]
- Wan, M.L.Y.; Co, V.A.; El-Nezami, H. Dietary polyphenol impact on gut health and microbiota. Crit. Rev. Food Sci. Nutr. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, G. Insulin and insulin resistance. Clin. Biochem. Rev. 2005, 26, 19–39. [Google Scholar]
- Gomes, A.C.; Bueno, A.A.; de Souza, R.G.; Mota, J.F. Gut microbiota, probiotics and diabetes. Nutr. J. 2014, 13, 60. [Google Scholar] [CrossRef] [Green Version]
- Vamanu, E.; Pelinescu, D.; Sarbu, I. Comparative Fingerprinting of the Human Microbiota in Diabetes and Cardiovascular Disease. J. Med. Food. 2016, 19, 1188–1195. [Google Scholar] [CrossRef]
- Wakita, Y.; Shimomura, Y.; Kitada, Y.; Yamamoto, H.; Ohashi, Y.; Matsumoto, M. Taxonomic classification for microbiome analysis, which correlates well with the metabolite milieu of the gut. BMC Microbiol. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Tapia, M.; Miller, A.W.; Granados -Portillo, O.; Tovar, A.R.; Torres, N. The development of metabolic endotoxemia is dependent on the type of sweetener and the presence of saturated fat in the diet. Gut Microbes 2020, 12, 1801301. [Google Scholar] [CrossRef] [PubMed]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijingoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Zhang, P.; Shen, L.; Niu, L.; Tan, Y.; Chen, L.; Zhao, Y.; Bai, L.; Hao, X.; Li, X.; et al. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2020, 21, 6356. [Google Scholar] [CrossRef]
- Wei, Y.; Liang, J.; Su, Y.; Wang, J.; Amakye, W.K.; Pan, J.; Chu, X.; Ma, B.; Song, Y.; Li, Y.; et al. The associations of the gut microbiome composition and short-chain fatty acid concentrations with body fat distribution in children. Clin. Nutr. 2020. [Google Scholar] [CrossRef] [PubMed]
- Hellerstein, M.K.; Christiansen, M.; Kaempfer, S.; Kletke, C.; Wu, K.; Reid, J.S.; Mulligan, K.; Hellerstein, N.S.; Shackleton, C.H. Measurement of de novo hepatic lipogenesis in humans using stable isotopes. J. Clin. Investig. 1991, 87, 1841–1852. [Google Scholar] [CrossRef]
- Wolever, T.M.; Spadafor, A.P.; Eshuis, H. Interaction between colonic acetate and propionate in humans. Am. J. Clin. Nutr. 1991, 53, 681–687. [Google Scholar] [CrossRef]
- Corsi, G.C.; Yoshime, L.T.; Corrêa, T.A.F.; Rogero, M.M. Bioactive compounds and inflammation: An overview. Nutrire 2020, 45, 14. [Google Scholar] [CrossRef]
- Alexander, C.; Swanson, K.S.; Fahey, G.C., Jr.; Garleb, K.A. Perspective: Physiologic Importance of Short-Chain Fatty Acids from Nondigestible Carbohydrate Fermentation. Adv. Nutr. 2019, 10, 576–589. [Google Scholar] [CrossRef]
- Pahl, H.L. Activators and target genes of Rel/NF-kappaB transcription factors. Oncogene 1999, 18, 6853–6866. Available online: https://www.nature.com/articles/1203239.pdf (accessed on 2 January 2021). [CrossRef] [Green Version]
- Meijer, K.; Vonk, R.J.; Priebe, M.G.; Roelofsen, H. Cell-based screening assay for anti-inflammatory activity of bioactive compounds. Food Chem. 2015, 166, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Beloborodova, N.V.; Bairamov, I.T.; Olenin, A.Y.; Shubina, V.S.; Teplova, V.V.; Fedotcheva, N. Effect of phenolic acids of microbial origin on production of reactive oxygen species in mitochondria and neutrophils. J. Biomed. Sci. 2012, 19, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beloborodova, N.V.; Osipov, A.A.; Bedova, A.Y.; Khabib, O.N. Low-Molecular Weight Bacterial Metabolites in Host-Microbial Interaction. J. Infect. NonInfect. Dis. 2016, 2. [Google Scholar] [CrossRef]
- Chernevskaya, E.; Beloborodova, N.; Klimenko, N.; Pautova, A.; Shilkin, D.; Gusarov, V.; Tyakht, A. Serum and fecal profiles of aromatic microbial metabolites reflect gut microbiota disruption in critically ill patients: A prospective observational pilot study. Crit. Care 2020, 24, 312. [Google Scholar] [CrossRef] [PubMed]
- Caussy, C.; Hsu, C.; Lo, M.T.; Liu, A.; Bettencourt, R.; Ajmera, V.H.; Bassirian, S.; Hooker, J.; Sy, E.; Richards, L.; et al. Genetics of NAFLD in Twins Consortium Link between gut-microbiome derived metabolite and shared gene-effects with hepatic steatosis and fibrosis in NAFLD. Hepatology 2018, 68, 918–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beloborodova, N.; Pautova, A.; Sergeev, A.; Fedotcheva, N. Serum levels of mitochondrial and microbial metabolites reflect mitochondrial dysfunction in different stages of sepsis. Metabolites 2019, 9, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Veledo, S.; Joan Vendrell, J. Gut microbiota-derived succinate: Friend or foe in human metabolic diseases? Rev. End. Metab. Dis. 2019, 20, 439–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Björck, I.; Bäckhed, F. Dietary Fiber-Induced Improvement in Glucose Metabolism Is Associated with Increased Abundance of Prevotella. Cell Metab. 2015. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gatea, F.; Sârbu, I.; Vamanu, E. In Vitro Modulatory Effect of Stevioside, as a Partial Sugar Replacer in Sweeteners, on Human Child Microbiota. Microorganisms 2021, 9, 590. https://doi.org/10.3390/microorganisms9030590
Gatea F, Sârbu I, Vamanu E. In Vitro Modulatory Effect of Stevioside, as a Partial Sugar Replacer in Sweeteners, on Human Child Microbiota. Microorganisms. 2021; 9(3):590. https://doi.org/10.3390/microorganisms9030590
Chicago/Turabian StyleGatea, Florentina, Ionela Sârbu, and Emanuel Vamanu. 2021. "In Vitro Modulatory Effect of Stevioside, as a Partial Sugar Replacer in Sweeteners, on Human Child Microbiota" Microorganisms 9, no. 3: 590. https://doi.org/10.3390/microorganisms9030590
APA StyleGatea, F., Sârbu, I., & Vamanu, E. (2021). In Vitro Modulatory Effect of Stevioside, as a Partial Sugar Replacer in Sweeteners, on Human Child Microbiota. Microorganisms, 9(3), 590. https://doi.org/10.3390/microorganisms9030590