1. Introduction
Nono is a spontaneously lactic acid fermented milk that originates from the northern parts of Nigeria. It is predominantly prepared and sold in local markets by the Hausa/Fulani cattle herdsmen [
1,
2]. Generally,
nono is prepared in homes from unpasteurised cow’s milk collected in a calabash and allowed to ferment naturally for 24 h [
3,
4,
5] before being sold to rural and urban people on markets [
6]. Depending on uncontrolled production processes and too poor hygiene during preparation, bacteria other than lactic acid bacteria (LAB), including foodborne pathogenic bacteria such as
Escherichia coli,
Salmonella, Shigella, Staphylococcus aureus, and
Bacillus cereus can occur in these products [
2,
5,
6,
7,
8]. The presence of foodborne pathogens in such products suggests that the
nono products are probably not fermented to a pH sufficiently fast and low enough to inhibit such bacteria. Furthermore, the production of
nono by spontaneous fermentation with unpasteurized milk leads to varying products with different levels of quality inconsistency and safety. In order to obtain a well-controlled nono fermentation with a fast and deep acidification, one would benefit from the use of starter cultures. Upon inoculation of pasteurized or non-pasteurized milk with suitable starters at high concentration, the pH could be rapidly lowered to pH 4.50–4.0, at which many foodborne pathogens will not grow [
9].
Thus far, there have not been many studies on the diversity of LAB found during fermentation of
nono. Banwo et al. [
10] showed that quite a diverse range of LAB were associated with
nono production, identifying
Lactobacillus (
Lb.)
plantarum,
Enterococcus spp., and
Pediococcus spp., while Okagbue and Bankole [
1] identified
Lactococcus (
Lc.)
lactis subsp.
lactis biovar diacetylactis (in the original publication termed
Streptococcus diacetilactis),
Lb. brevis, and
Saccharomyces (
S.) cerevisiae to be associated with the fermentation. The authors of [
11] isolated
Lb. plantarum, Lb. brevis, Lb. delbrueckii subsp.
bulgaricus, and
Limosilactobacillus fermentum from
nono. In addition, the authors [
11] indicated that yeasts, especially
S. cerevisiae, were also associated with the fermentation. The aim of this study was, therefore, to characterize the predominant LAB and yeast species associated with
nono fermentation and to perform a preliminary investigation of their technological properties in order to select potential starter cultures for controlled
nono production.
2. Materials and Methods
2.1. Microbiological Analysis of Nono Samples from the Nigerian Market
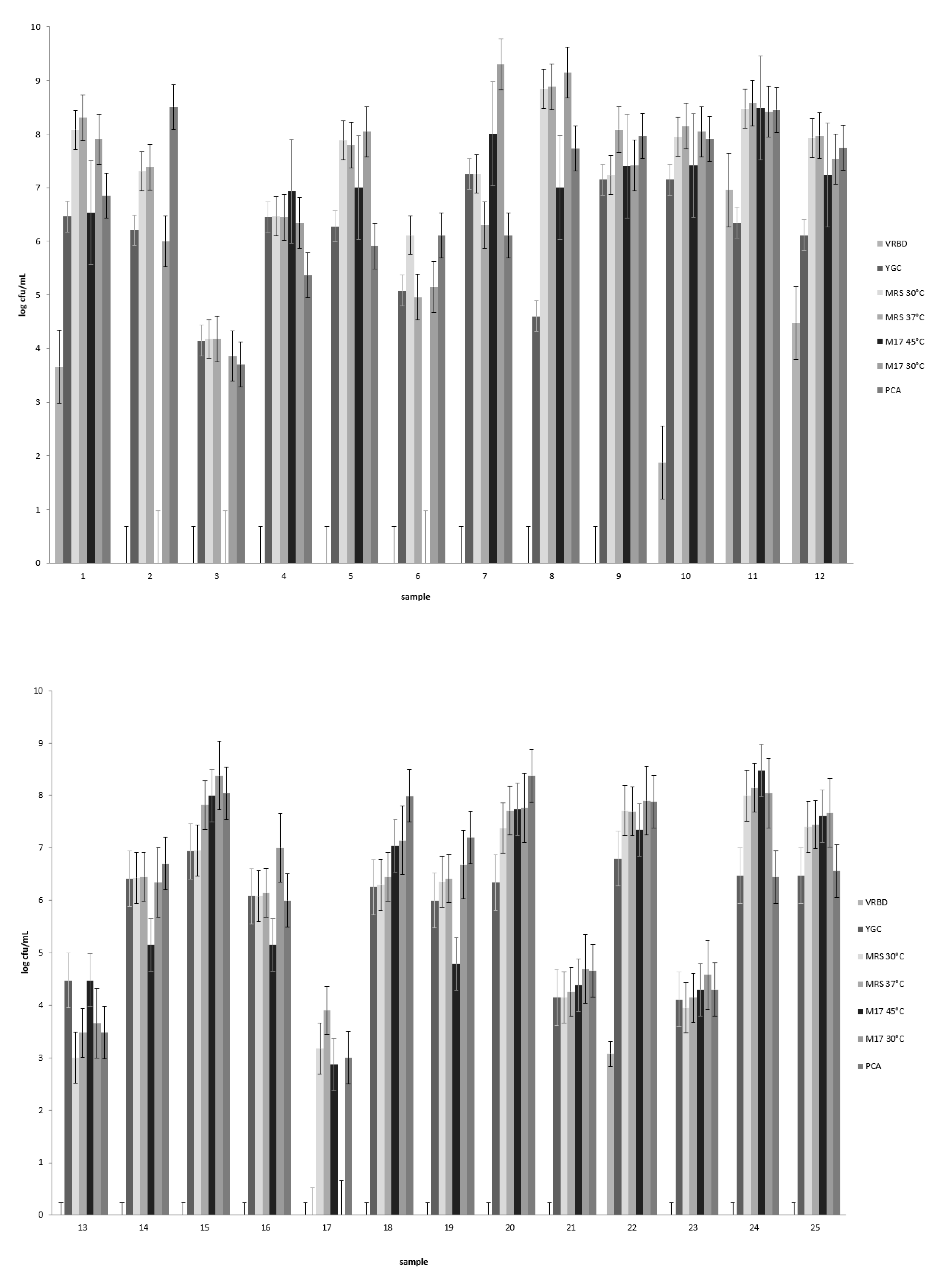
For a general microbiological analysis, 25 nono samples were bought from Fulani processors and local markets in Kano, Katsina, Jigawa, and Bauchi states in northern Nigeria. The samples were transported under cooled conditions to the laboratory, and the pH of the nono samples was measured using a pH electrode (Mettler Tuledo Inlab Expert Pt 1000, Gießen, Germany). Samples were then diluted in a serial tenfold-dilution series using quarter-strength Ringers solution (Merck, Darmstadt, Germany). After this, 100 µL aliquots of appropriate dilutions were plated out in duplicate onto different selective culture media in order to determine the total amount of aerobic, mesophilic bacteria on plate count agar (VWR Chemicals, Leuven, Belgium); LAB on de Man, Rogosa, and Sharpe agar (MRS agar, Merck, Darmstadt, Germany); and Gram-negative bacteria on violet red bile dextrose agar (VRBD agar, Merck, Darmstadt, Germany) incubated at 30 °C and streptococci on M17 medium incubated at 45 °C. In addition to selective enumeration of bacteria, the numbers of yeasts in the samples were determined using yeast extract–glucose–chloramphenicol agar (YGC agar, Merck, Darmstadt, Germany).
2.2. S Amplicon Sequencing Analysis of the Nono Bacterial Community
The bacterial community composition of the
nono samples was also determined using tag-encoded, 16S rRNA gene MiSeq-based (Illumina, San Diego, CA, USA) high-throughput sequencing. The total bacterial DNA was extracted using the repeated bead beating plus column (RBB+C) method as described previously by Yu and Morrison [
12] from 20 mL of each of the products. The DNA concentration of the samples was measured using the Qubit 3.0 fluorometer with the high sensitivity kit (Thermo Scientific, Darmstadt, Germany) and was then diluted to 5 ng/µL. DNA amplification was carried out following the two-stage PCR protocol provided by Illumina. The first PCR step was carried out in a final volume of 25 µL containing 2.5 µL template DNA (5 ng/µL), 5 µL of the primers 16S Fw (5′-TCG TCG GCA GCG TCA GAT GTG TAT AAG AGA CAG CCT ACG GGN GGC WGC AG-3′) and 16S Rev (5′-GTC TCG TGG GCT CGG AGA TGT GTA TAA GAG ACA GGA CTA CHV GGG TAT CTA ATC C-3′) binding to the the V3 and V4 hypervariable regions of the 16S rRNA gene, respectively (final concentration 1 µM) (Illumina 16S Metagenomic Sequencing Library Preparation protocol), and 12.5 µL of KAPA HiFi HotStart Ready Mix (Kapa Biosystems, Inc. Wilmington, MA, USA). The PCR reactions were performed in a PeqSTAR thermal cycler (Peqlab, Erlangen, Germany) using the following program: initial denaturation at 95 °C for 3 min, followed by 25 cycles of 95 °C for 30 s, 55 °C for 30 s, and 72 °C for 30 s, with a final elongation at 72 °C for 5 min. The sizes of the 16S rRNA gene PCR products were confirmed by automated electrophoresis using the Experion Automated Electrophoresis System (BioRad, Munich, Germany). The PCR products were cleaned using AMPure XP magnetic beads (Agencourt) and then used as a template for the second PCR, the index PCR, to attach dual indices with the Nextera XT Index kit (Illumina). PCR was performed using 25 µL KAPA HiFi HotStart Ready Mix (KAPPA BIOSYSTEMS), with an initial denaturation at 95 °C for 3 min, followed by 8 amplification cycles (denaturing at 95 °C for 30 s; annealing at 55 °C for 30 s; extension at 72 °C for 30 s) with a final extension at 72 °C for 5 min. The index PCR products were cleaned with AMPure XP beads (Agencourt) and the final DNA concentration was measured using the Qubit 3.0 fluorometer with the broad range kit (Thermo Scientific). The final DNA concentration of each sample was adjusted to 4 nM with 10 mM Tris (pH 8.5), and 5 µL of diluted DNA from each sample were mixed together for pooling the libraries with unique indices. The pooled samples were further diluted using the Illumina Hybridization Buffer (HT1) to a final loading concentration of 6 pM. Sequencing of the 16S rRNA gene-based metagenome was performed in Miseq Illumina following the Illumina instructions for metagenomics workflows. The resulting fastq files were analyzed using the
de novo analysis pipeline at the IMNGS platform for ecology and diversity studies [
13]. The demultiplexed Illumina fastq files were pre-processed using the remultiplexer pearl script recommended by the IMNGS platform and unknown sequences belonging to ambiguous sequences, archaea, eukaryota, chloroplasts, and mitochondria were excluded. The processed sequences were uploaded to the MG-RAST and analyzed using the RDP taxonomic database. The relative abundance of OTUs were calculated and visualized using the implanted pipelines of MG-RAST.
2.3. Isolation and Identification of Predominant Lactic Acid Bacteria and Yeasts from Nono
In order to accurately characterize the predominant lactic acid bacteria and yeasts associated with
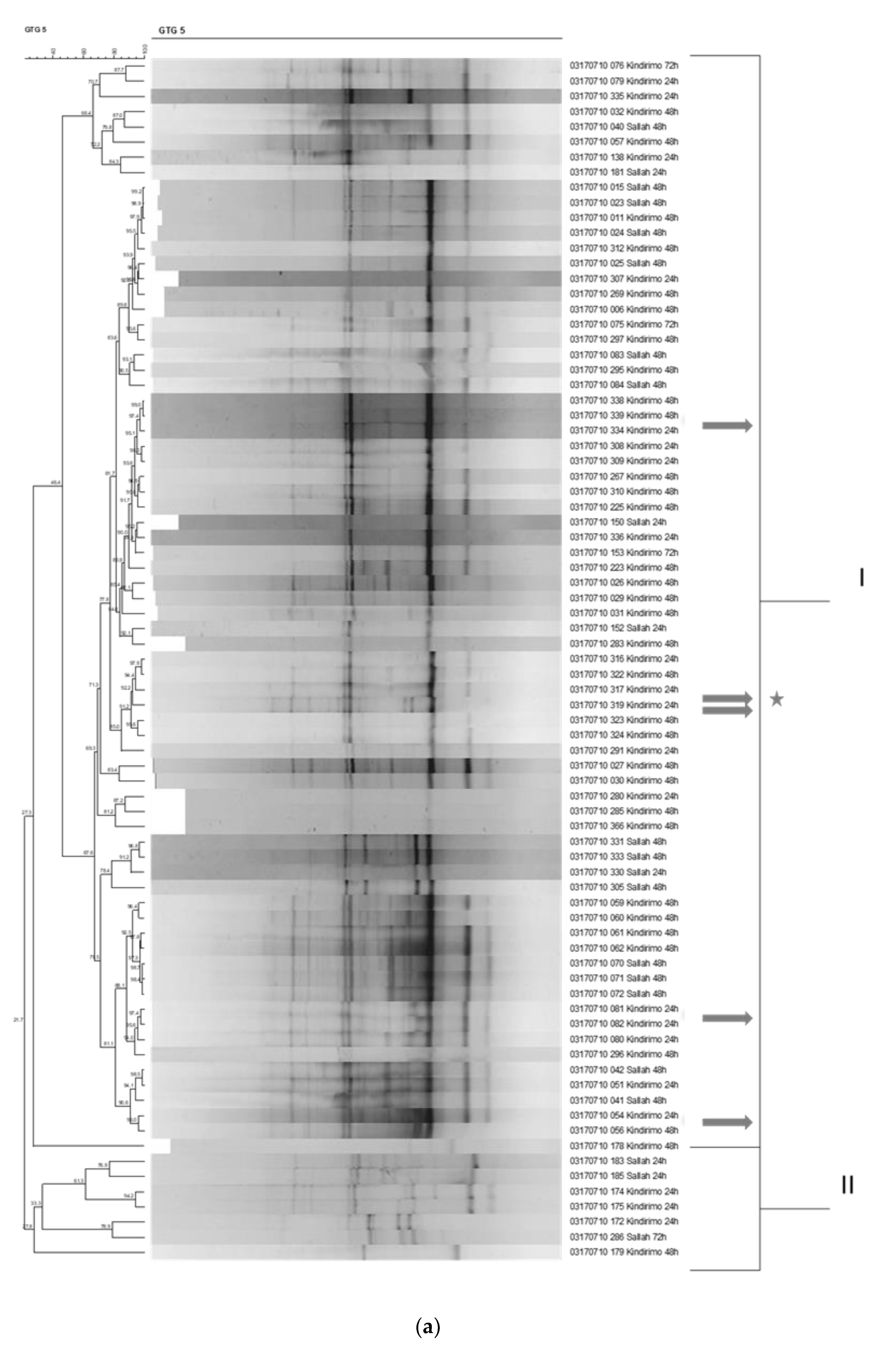
nono fermentation, we isolated colonies from MRS agar and YGC agar plates used to determine the lactic acid bacterial and yeast counts, respectively. For the LAB identification, a polyphasic approach to identify the strains was adopted. For this, strains were first presumptively identified as LAB by phenotypic tests and then grouped by repetitive element PCR (rep-PCR) fingerprinting to the species level [
14,
15]. Representative isolates of the major groups were then characterized by 16S rRNA gene sequencing, indicating the rest of the closely grouped isolates to presumptively be the same species. Of these isolates, the genomes of selected strains with potential for use as starter culture were sequenced in order to determine the presence of technologically important genes for the fermentation. Colonies were randomly selected from the plates onto which the highest dilutions were plated. For LAB, colonies were purified by streaking onto MRS agar plates (at least 3 times). A total of 223 presumptive lactic acid bacteria strains thus isolated were tested for Gram-reaction using the 3% KOH method, for the presence of the enzyme catalase, and for production of CO
2 from glucose fermentation, as described previously [
14,
15]. Of these, 147 were selected for further characterization by rep-PCR.
Yeasts were purified by streaking onto YGC agar, cultured on malt extract agar (2% malt extract, Carl Roth; 0.1% peptone from soybean, Fluka; 1.5% agar-agar, VWR) at 25 °C. A total of 43 isolates were identified using a commercial test kit (API BioMérieux ID 32 C, Nürtingen, Germany), which tests for utilization of different sugars, and the results were transformed into an API code that is analyzed using the database of the apiweb (BioMérieux) and the Westerdijk Fungal biodiversity institute.
The total genomic DNA was isolated from each of the 147 presumptive LAB strains according to the method of [
16] as modified by [
17], and the bacteria were fingerprinted using rep-PCR fingerprinting as previously described [
14]. Rep-PCR was performed using the primer GTG
5 (5′-GTG GTG GTG GTG GTG-3′), and PCR products were separated by electrophoresis on a 1.8% (w/v) agarose gel using 1 × TBE buffer. Groupings of rep-PCR fingerprints were performed by means of the Pearson product-moment correlation coefficient (
r) and the unweighted pair-group method using arithmetic averages clustering algorithm (UPGMA) [
18] using the Bionumerics (version 7.2.6) software package (Applied Maths, Sint-Martens-Latem, Belgium).
The almost complete 16S rRNA gene (maximal 1540 bp) was also amplified by PCR from total genomic DNA extracted as described above from 16 isolates characteristic of major rep-PCR groups. The PCR products were amplified in 50 µL volumes containing 100 ng template DNA, 1 × Taq DNA polymerase buffer (Genaxxon, Ulm, Germany), 125 µM of each dNTP (CarlRoth, Karlsruhe, Germany), 25 pM of each forward and reverse primer (16S fw 5′-AGA GTT TGA TCM TGG CTC AG-3′ and 16S rev 5′-TAC GGY TAC CTT GTT ACG ACT-3′), and 1.5 U Taq DNA polymerase (Genaxxon, Ulm, Germany). The PCR reaction was performed using an initial denaturation step at 94 °C for 3 min, followed by 32 cycles of 94 °C for 30 s, 55 °C primer annealing for 30 s, 72 °C extension for 1 min 30 s, followed by a final extension step at 72 °C for 5 min. All PCR products were purified using PCR cleaning columns (Peqlab) and subsequently sequenced at Eurofins (Eruofin, Cologne, Germany). The sequences of 16S rRNA gene PCR products were compared to those present in the EzTaxon database [
19].
The total genomic DNA of each of the selected potential starter cultures was isolated using the peqGOLD bacterial DNA kit (Peqlab, Erlangen, Germany) after growing the strain overnight at 30 °C in MRS broth. The sequencing library was constructed using the Nextera Flex DNA library kit (Illumina, San Diego, CA, USA) and sequenced with 2 × 251 bp reads on an Illumina MiSeq platform. The Trimmomatic version 0.36 for trimming of Illumina paired-end [
20] software was used to remove adapter sequences with the following parameters (sliding window; 4:15 and leading;3) as well as low-quality reads, and reads with a minimum contig length of 500 bp and fivefold coverage were then
de novo assembled using SPAdes v3.13.1 with careful and k-mer 77 parameters [
21]. Annotation of the draft genome sequences was performed with the NCBI Prokaryotic Genome Annotation Pipeline v4.10 using default parameters [
22]. The whole genome sequences of the
Lb. helveticus strain 314 and
Streptococcus (
S.)
thermophilus strain 252 have been deposited in DDB/GenBank under the accession numbers JAFIWI000000000 and JAFIWH000000000.
2.4. Milk Fermentation with Starter Lactic Acid Bacteria and Yeast Strains
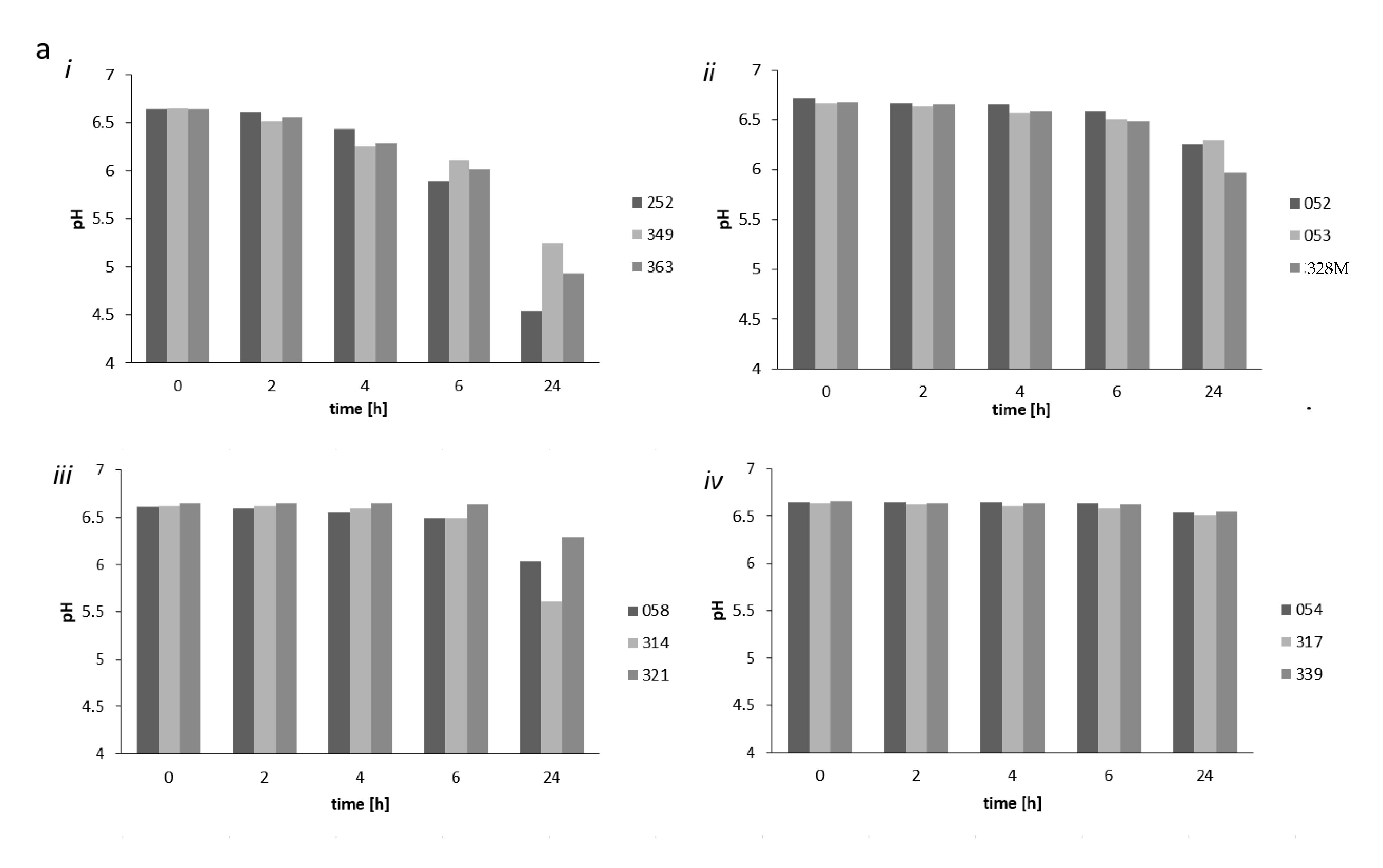
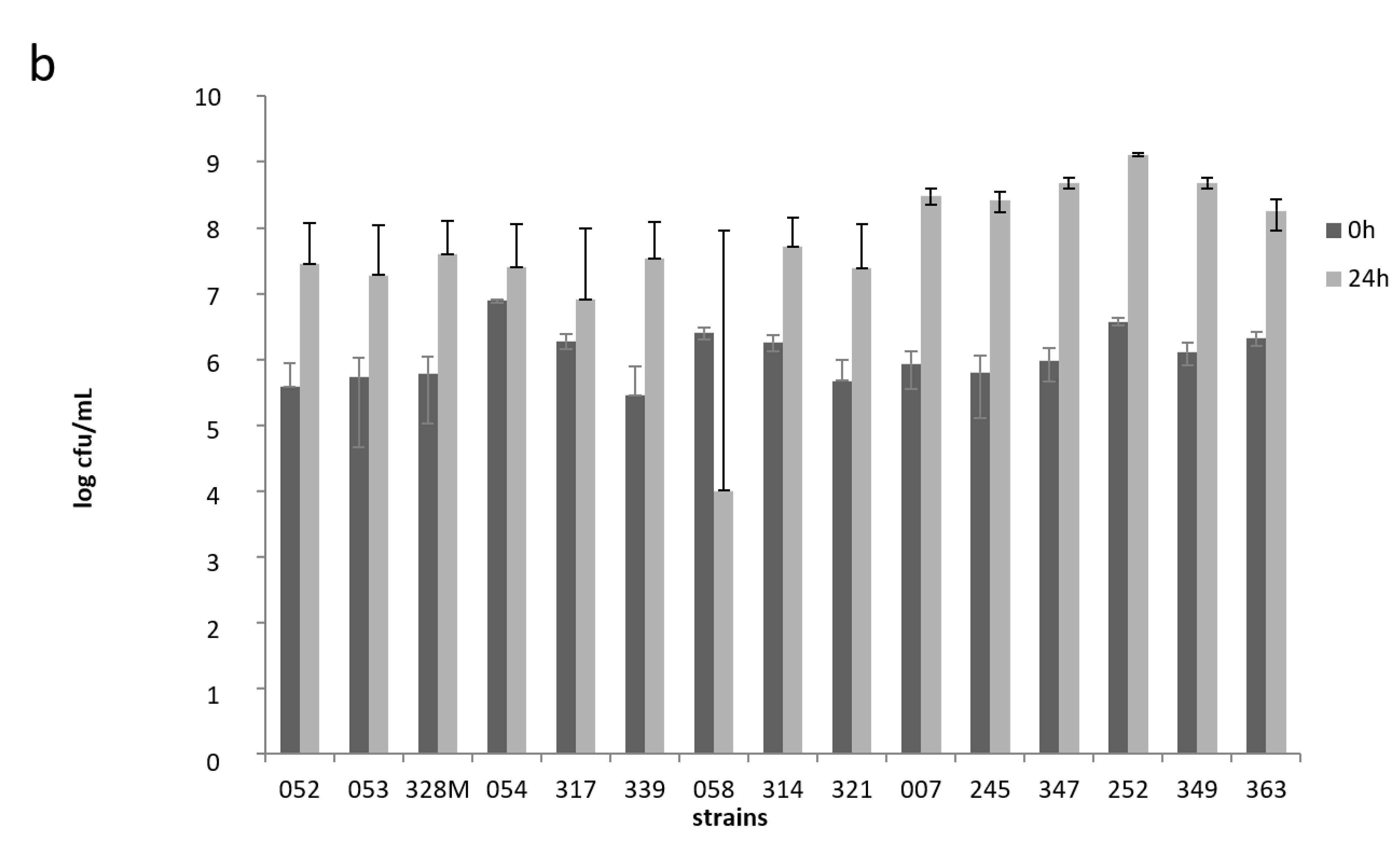
To determine the growth and acidification ability of potential starter strains, we obtained pasteurized cow’s milk used in fermentation experiments from the Max Rubner-Institute’s experimental farm located in Schädtbek just outside the city of Kiel. The acidifying capacity of selected strains (S. thermophilus strains 252, 349, and 363; Lb. helveticus strains 058, 314, and 321; L. fermentum strains 054, 317 and 339; as well as Lb. delbrueckii strains 052, 053, and 328M) was tested in freshly pasteurized (90–95 °C for 15 min) whole milk at 30 °C for 24 h. Strains from these 4 species were chosen, as they represent the predominant LAB species associated with nono samples (see below). An incubation temperature of 30 °C was chosen as nono fermentation was performed according to traditional local methods, where the milk is left to ferment at ambient temperatures that can reach 30 °C in Nigeria. For the acidification experiments, the strains were inoculated at 1 × 106 cfu/mL in single-strain fermentations. The milk pH was measured after 2, 4, 6, and 24 h fermentation. The growth of the potential starter strains was assessed upon inoculation of the milk at 0 and after 24 h fermentation by plating 100 µL of appropriate tenfold dilutions of the milk in quarter-strength Ringers solution onto MRS agar, and incubating plates for 48 h at 30 °C under gas pack anaerobically.
On the basis of pH reducing capacity and growth ability in the single-strain fermentation tests, we chose the
Lb. helveticus 314,
L. fermentum 317 [
23],
S. thermophilus 252,
Lb. delbrueckii 328M, and the yeast
S. cerevisiae 370 strains as possible starter cultures to test
nono production at laboratory scale in 50 mL volumes. These strains were tested as multi-strain starters for fermentation in co-cultures and included all 4 bacterial strains inoculated to a level of 1 × 10
6 cfu/mL and the yeast inoculated at 1 × 10
4 cfu/mL (starter A), as well as the 4 bacterial strains-inoculated at a level of 1 × 10
6 cfu/mL without the yeast (starter B). The yeast was chosen because Okagbue and Bankole [
1] reported that
S. cerevisiae was consistently isolated from
nono.
The milk was fermented in 50 mL volumes at 30 °C for 48 h, and the pH and numbers of bacteria were determined after 0 h (immediately after inoculation), 24 h, and 48 h. For enumeration, 1 mL of milk was collected and diluted in quarter-strength Ringer’s solution in a tenfold dilution series. Hundred microliter aliquots from appropriate dilutions were plated out onto MRS agar plates for determining the Lactobacillus sp. counts and on M17 agar for the S. thermophilus counts (only for the fermentation that contained S. thermophilus 252). The yeast counts were determined by plating onto YGC agar, to which 0.2% of a 10% tartaric acid solution was added after autoclaving in order to adjust the pH to 4.6. Both MRS agar (anaerobic) and M17 agar plates (aerobic) were incubated at 30 °C for 48 to 72 h, while YGC agar plates were incubated at 25 °C for 72 h.
2.5. Determination of Rheological Properties
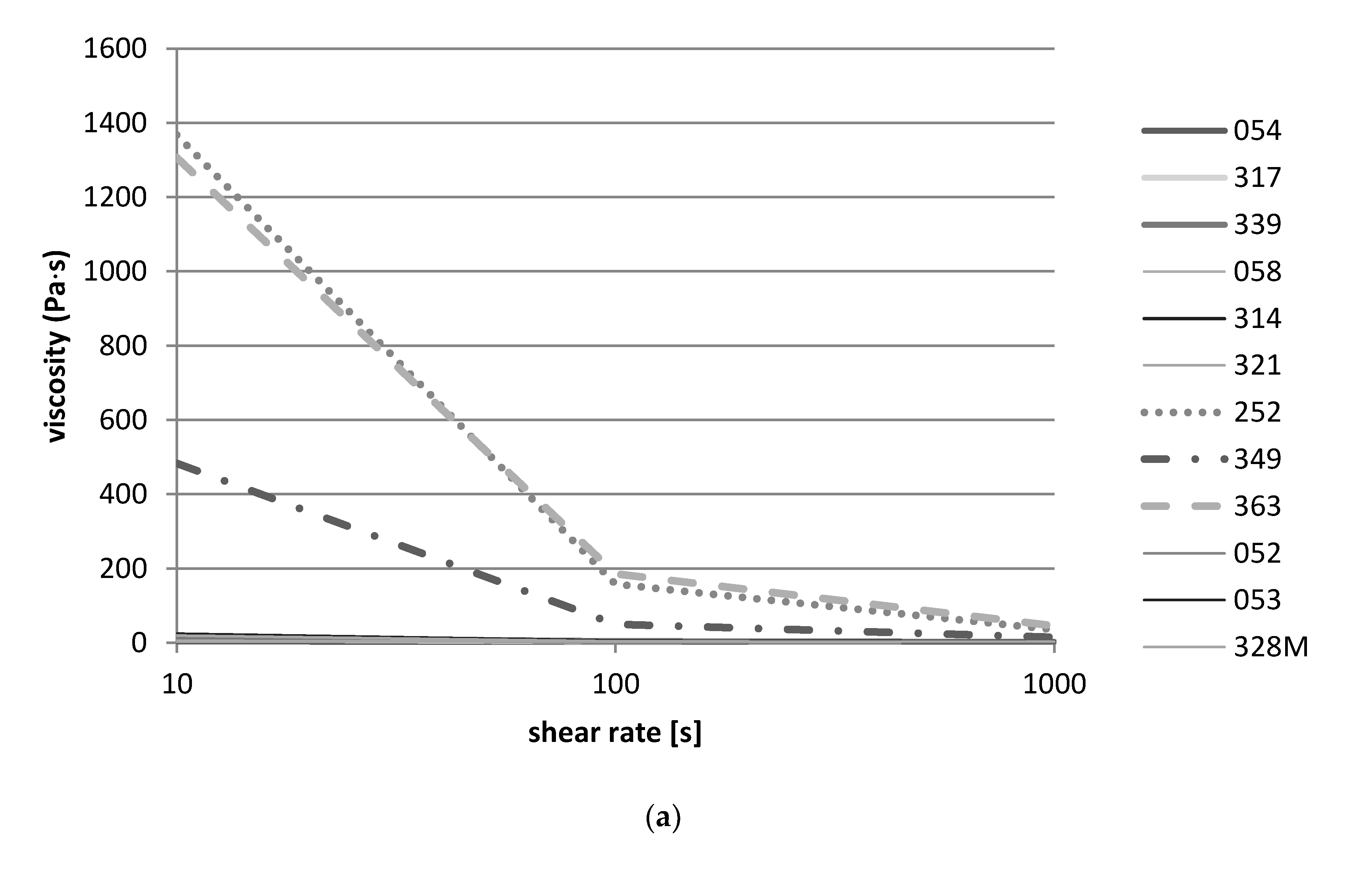
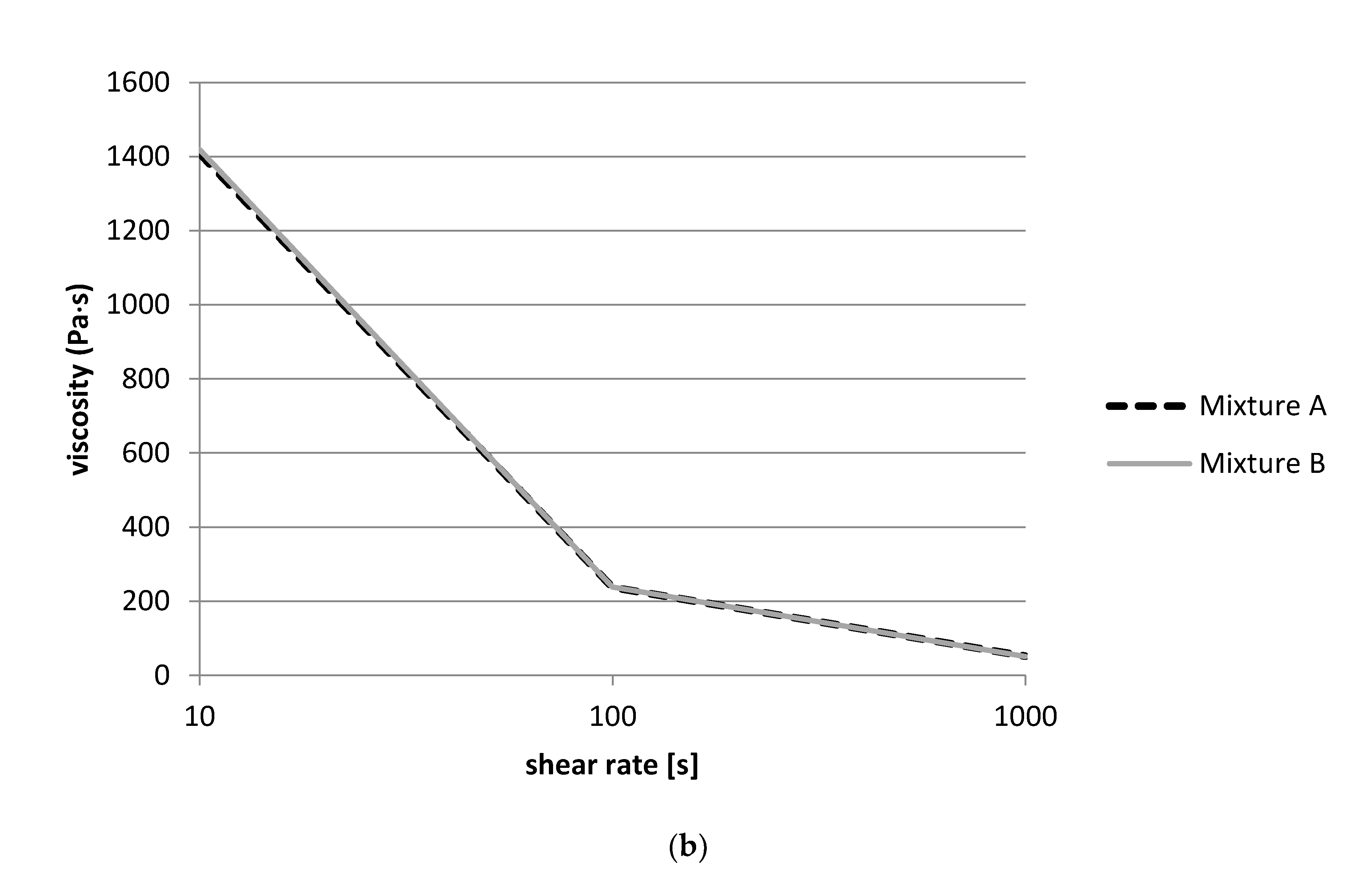
For the measurement of the viscosity, a rheometer MCR 302 (Anton Paar, Ostfildern, Germany) was used. Aliquots of 0.7 mL of the fermented product were placed into the cone and plate measuring system (diameter 50 mm) using a syringe without needle. A flow curve was recorded at a temperature of 20 °C using a logarithmic ramp of the shear rate (10 to 1000 s−1). These measurements were performed after 24 h and 48 h of fermentation.
4. Discussion
Generally, the bacterial counts determined on M17 medium incubated at 45 °C in this study were similar to those determined on the same M17 incubated at 30 °C, although in some samples the counts were higher on the latter medium by up to 1 cfu/mL. This indicates that not all the bacteria growing on M17 at 30 °C were streptococci, as the temperature of 45 °C is selective for these bacteria. M17 also supports the growth of lactococci at 30 °C. Similar to our results, Okonkwo [
2] reported highly variable counts of different bacterial groups in
nono samples from Nigerian markets, and in that study, the mesophilic, aerobic plate counts ranged from log 2.53 to log 4.89 per mL, while enterobacterial counts ranged from log 1.37 to log 3.29 per mL. Yeasts were found to be present at levels ranging from log 0.81 to log 1.61 per mL [
2]. Overall, our results indicate that lactic acid bacteria (including presumptive streptococci) and yeasts represent predominant microorganisms in
nono products and that product occasionally also contained Gram-negative bacteria, which are perceived as a hygienic risk. The presence of various pathogenic bacteria in other studies [
2,
5,
6,
7,
8] confirmed this risk, and this may reflect the fact that
nono is traditionally produced from non-pasteurized milk under uncontrolled conditions without the use of starter cultures at a household scale.
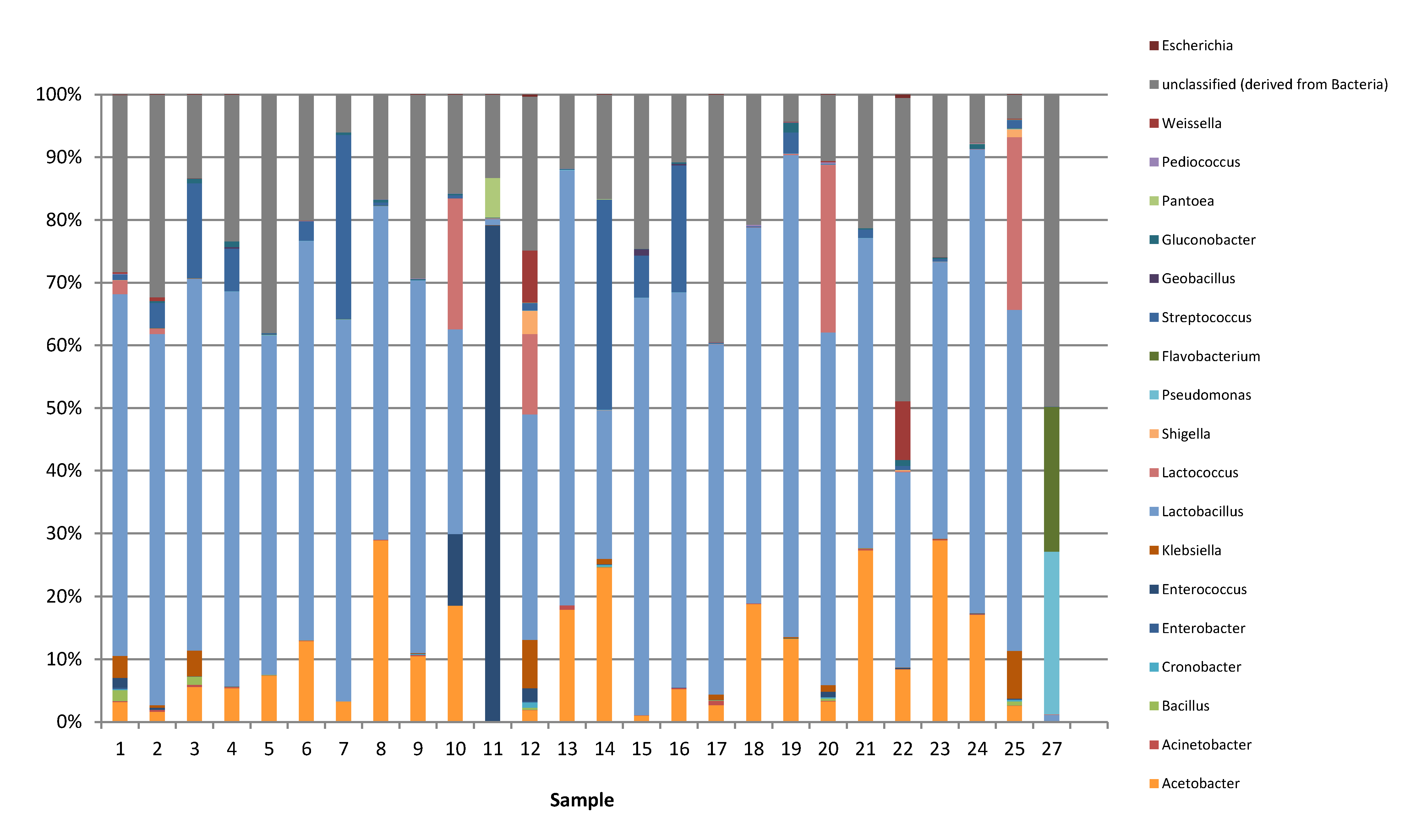
Metagenomics based on next generation sequencing with an Illumina MiSeq sequencer was performed to investigate the bacterial diversity of
nono samples. It should be mentioned here that streptococci and lactococci are phylogenetically closely related and that these may not be unequivocally identified on the basis of high throughput sequencing metagenomic studies. This is because only a small portion of the 16S rRNA gene (in this case the V3/V4 region) is sequenced, which may not be sufficiently discriminatory to identify these bacteria to genus level. The varying abundances of LAB in the
nono products indicate that they were not uniformly fermented and differed regarding the LAB population composition, which exemplifies that the drawback that the milk used in these fermentations was not inoculated with starter bacteria, as the use of an industrial starter would result in a more uniform population if the fermentation process was controlled. It was further noticed that enterobacteria such as
Shigella,
Escherichia, and
Klebsiella occasionally occurred in the fermented products, as was previously reported [
2,
5,
6,
7]. It is unlikely that these bacteria will be able to grow and multiply at a pH of 4.0 or less. Nevertheless, the results show that there is an inherent risk of infection by potentially pathogenic bacteria from these products whenever
nono is not properly fermented. The work of Okonkwo [
2] showed that the pH values of
nono samples from Nigerian markets varied from pH 4.0 to pH 5.4. As suggested by Holzapfel [
9], acidification to pH values of less than pH 4.2 constitutes a major safety concern for fermented foods. These data clearly show that there is a need for a controlled fermentation of
nono type products in order to guarantee their safety.
In order to determine the species composition of the lactic acid bacterial populations more accurately, we identified isolates to the species level by a polyphasic approach based on phenotypic and genotypic methods [
14,
15]. Repetitive element PCR (rep-PCR) was used as a major fingerprinting method. Overall, the results thus showed that the predominant LAB involved in the
nono fermentation included
L. fermentum, together with
Lb. helveticus,
S. thermophilus, and
Lb. delbrueckii strains. The last two microorganisms are well known for their role in yoghurt production and are utilized extensively as starter cultures for the production of this dairy product in western countries. Only few studies have been conducted on the lactic acid bacteria and their role in
nono fermentation.
L. fermentum, however, has been often isolated from African fermented foods [
25,
26,
27,
28,
29,
30], possibly as it is associated with vegetable fermentations performed in the same household/production facility. Previously, Okagbue and Bankole [
1] also reported
L. fermentum as the predominant lactic acid bacterium isolated from
nono. Interestingly, Adekosan et al. and Aforijiku and Onilude [
10,
29] also isolated
L. fermentum from
nono, in addition to other lactic acid bacteria including
Lb. delbrueckii subsp.
bulgaricus, Lb. plantarum, Lb. brevis, and
Lb. casei.
In addition to the lactic acid bacteria, the predominant yeasts isolated from the fermentation in this study were predominantly
Saccharomyces cerevisiae, as well as, to a lesser extent,
Candida novegensis strains. This confirms the report of Okagbue and Bankole [
1] that
Saccharomyces cerevisiae can be consistently isolated from
nono fermentations.
The use of single-strain starter cultures, especially when choosing a predominant
L. fermentum strain, would probably not lead to a sufficiently deep acidification to ensure the safety of the
nono product. This could be especially problematic in view of the fact that some potentially pathogenic Gram-negative bacteria were found in these products through the metagenomic microbiota analysis. Thus, for the development of a starter culture for
nono preparation, it seems necessary to incorporate representative strains of all the predominant lactic acid bacteria species shown to occur in the fermentation. As
Streptococcus thermophilus strains were noticeably capable of good growth and acid production in the milk, the incorporation of this species appears especially important. Even though
L. fermentum was revealed as a poor acid producer, it is known, on the other hand, to be involved in aroma production of African fermented foods, as well as for production of extracellular polysaccharides, which may play a role as bio-thickener in the product [
27].
Corroborating this position, Okagbue and Bankole [
1] also reported the development of starter cultures for
nono production. The work, however, based the starter cultures for
nono fermentation on
Lactococcus lactis subsp.
lactis biovar diacetylactis (referred to as
Streptococcus diacetilactis in the original publication),
Lactococcus lactis subsp.
cremoris (referred to as
Streptococcus cremoris in the original publication),
Lb. brevis, and
Saccharomyces cerevisiae. Diacetyl production is an important trait influencing the sensory properties and palatability of this product [
1]. In their study,
L. lactis subsp.
lactis biovar diacetylactis produced
nono with better taste when compared to
L. lactis subsp.
cremoris. Diacetyl production may thus be a key technological property for the starter bacteria used in the production of
nono. They confirmed that the individual strains showed marked differences in pH-reducing potential and concluded that
Lb. brevis was a poor acidifier of milk, while the
Lactococcus strains produced sufficient levels of acidity. The authors concluded therefore that single microorganism starter cultures should be avoided and that
L. lactis could be used together with
Lb. brevis as starter culture. As similar results were obtained also in our study, various starter cultures containing one representative strain of each of the four predominant LAB species were tested to determine their effect on functional aspects of
nono production, including milk acidification and rheological properties. Similar to Okagbue and Bankole [
1], we also opted to test these starter cultures either with or without addition of the yeast
Saccharomyces cerevisiae.
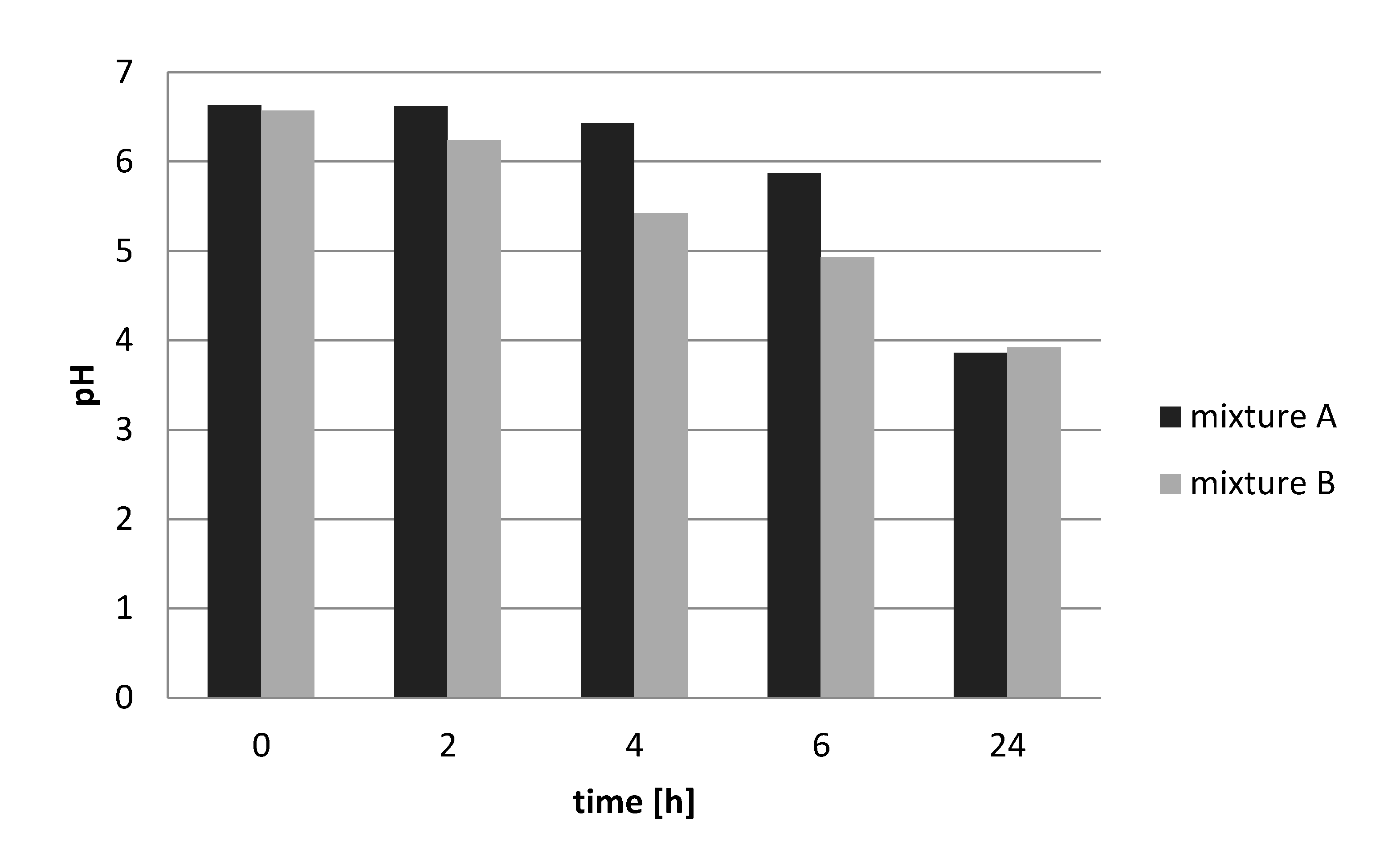
Both starter combinations tested achieved a pH reduction from 6.5 to less than 4.0, a pH level that according to Holzapfel [
9] would suffice in terms of guaranteeing the safety of these fermented products. As seen from single-strain incubations, this was probably the result of the high acid-producing capacity of the
S. thermophilus strain 252. These results indicate that the use of starter culture strains could result in products with high levels of these bacteria that stabilized the products by acidification to a pH of 4.0. Furthermore, varying levels of microorganisms of different bacterial groups, as determined in products obtained on Nigerian markets, could be avoided by the use of starter cultures, which would quickly dominate the fermentation. We did not examine yeast as single starter culture. Hence, whether or not
Saccharomyces cerevisiae plays an important role in the fermentation has still to be determined in more detail. Previously, Okagbule and Bankole [
1] had suggested that the incorporation of the yeast did not result in better sensory characteristics or higher diacetyl levels, and concluded that it was relatively unimportant in
nono production and that its use in a commercial situation would be unnecessary.
Data from the rheological study corresponds well with the results of the pH measurements, which showed that Lb. delbrueckii, L. fermentum, and Lb. helveticus strains from nono did not lower the pH below 5.0 and were thus not able to gel the milk. Again, the flow thinning behavior of S. thermophilus could be explained by the higher acid-producing capacity of the S. thermophilus strains, which could lower the pH of the milk during fermentation to below pH 5.0, causing the milk to gel and to thicken.
The
Lb. helveticus 314 and
L. fermentum 317 strains possessed the gene for citrate lyase [
23], which can be important for citrate metabolism and for the potential production of the aroma compound diacetyl. The genomes of strain
L. fermentum 317 and
S. thermophilus 252 also contained genes for an acetolactate synthase that may also be involved in diacetyl production. Only the genome of
Lb. delbrueckii 328M did not show the presence of any genes that may be related to diacetyl production. Diacetyl production has previously been described to be important for starter bacteria involved in
nono production [
1], and thus the starters selected in this study appeared to possess the genes required for this important technological function. The presence of the signature genes for folate biosynthesis (dihydropteroate synthase (
folP) and 2-amino-4-hydroxy-6-hydroxymethyldihydropteridine diphosphokinase (
folK)) indicates that the starter (culture) strains may hypothetically be capable of folate production. This is another functionally important trait for starter bacteria for African fermented foods [
31]. The
Lb. helveticus 314 and
S. thermophilus 252 strains possessed the bacteriocin genes for a class III bacteriocin and class IIb bacteriocin, respectively. The production of bacteriocin may hypothetically be helpful for inhibiting Gram-positive pathogenic bacteria, and thus these starter cultures may contribute to the safety of
nono products.
5. Conclusions
Overall, our metagenomic and, more importantly, strain typing study showed that lactobacilli and streptococci dominated the lactic acid microbiota of fermented nono. The bacterial species predominantly determined to occur were Lb. helveticus, L. fermentum, Lb. delbrueckii, and S. thermophilus strains, while the predominant yeast occurring in nono was Saccharomyces cerevisiae. Starter cultures development for nono production should thus be based on a combination of these predominant strains. Lb. delbrueckii and S. thermophilus are typical yoghurt starter bacteria that grow synergistically. Using metagenomics, pathogens such as Shigella and potential pathogens such as enterobacteria could be detected at low abundances. Starter selection for nono fermentation should include suitable technological strains of the bacterial species that are predominantly occurring in nono, i.e., Lb. helveticus, L. fermentum, Lb. delbrueckii, and S. thermophilus strains. However, the predominant L. fermentum, Lb. helveticus, and Lb. bulgaricus strains did not show a high capacity for acid production and consequently for milk gelling as a result of pH decrease. S. thermophilus strains, on the other hand, showed good acidification properties, and rheological results showed the strains’ capacity for gelling of milk to a yogurt consistency. As Lb. delbrueckii and S. thermophilus are known to support each other’s growth in yoghurt, Lb. delbrueckii was considered an important starter also for the production of nono. L. fermentum and Lb. helveticus may, on the other hand, also be important for production of flavor metabolites such as diacetyl. In this study, therefore, predominant strains were selected for further development as starter bacteria. These strains show that when used in combination as starters, they are effective in growing and lowering the pH of milk, causing gelling of the product to a yoghurt-like consistency. All strains, except for the Lb. delbrueckii strain, possess genes potentially involved in diacetyl production and folate biosynthesis signature genes. These strains should therefore be further studied in Nigeria to test for their success in nono fermentations under local manufacturing conditions.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








