
Taxonomic Re-Examination of Nine Rosellinia Types (Ascomycota, Xylariales) Stored in the Saccardo Mycological Collection

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimens Sampling and Morphology
2.2. DNA Extraction, PCR Amplification, Sequencing and Data Analysis
2.3. Phylogenetic Analysis
3. Results
3.1. Phylogenetic Analysis
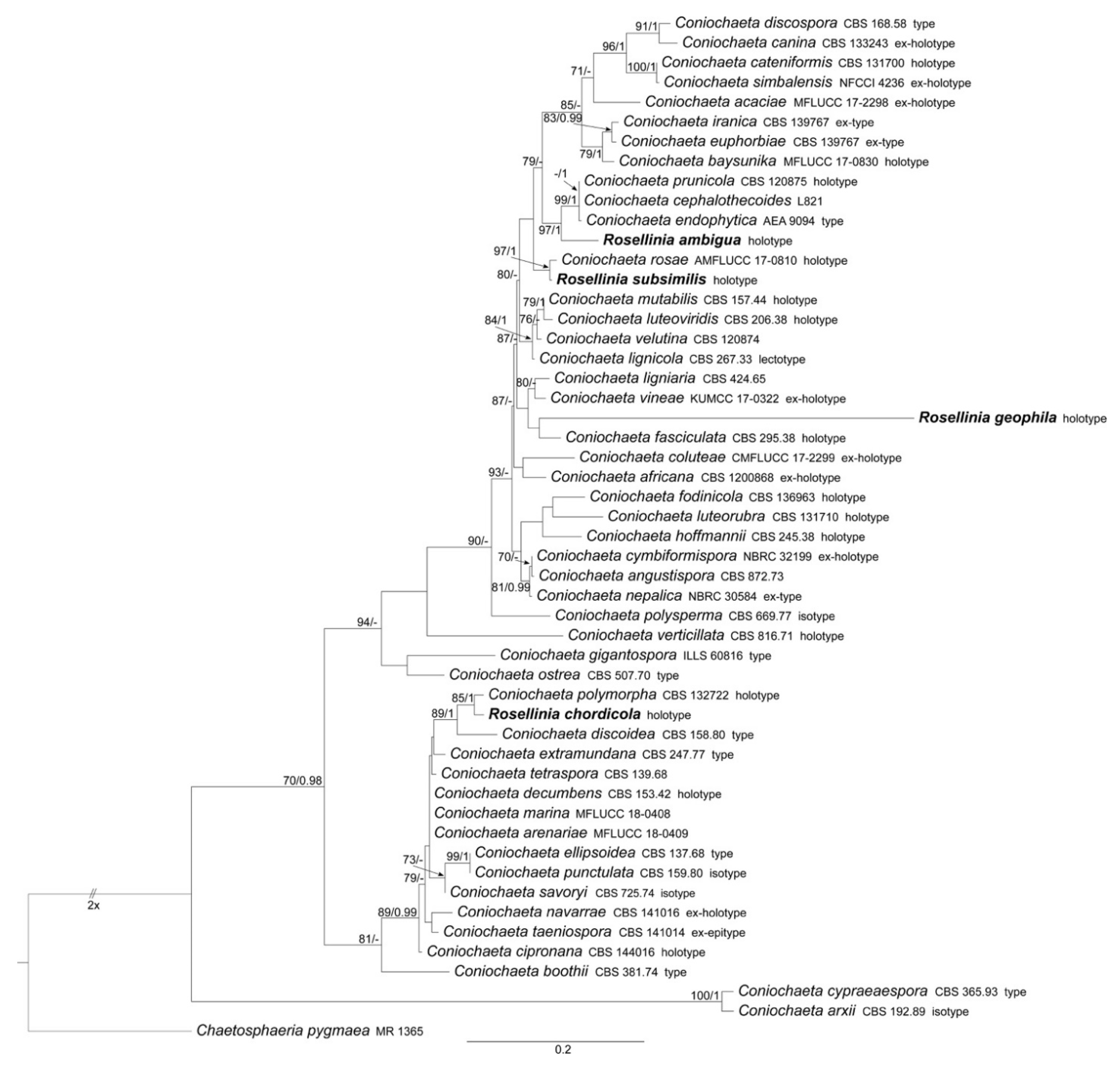
3.1.1. Coniochaeta
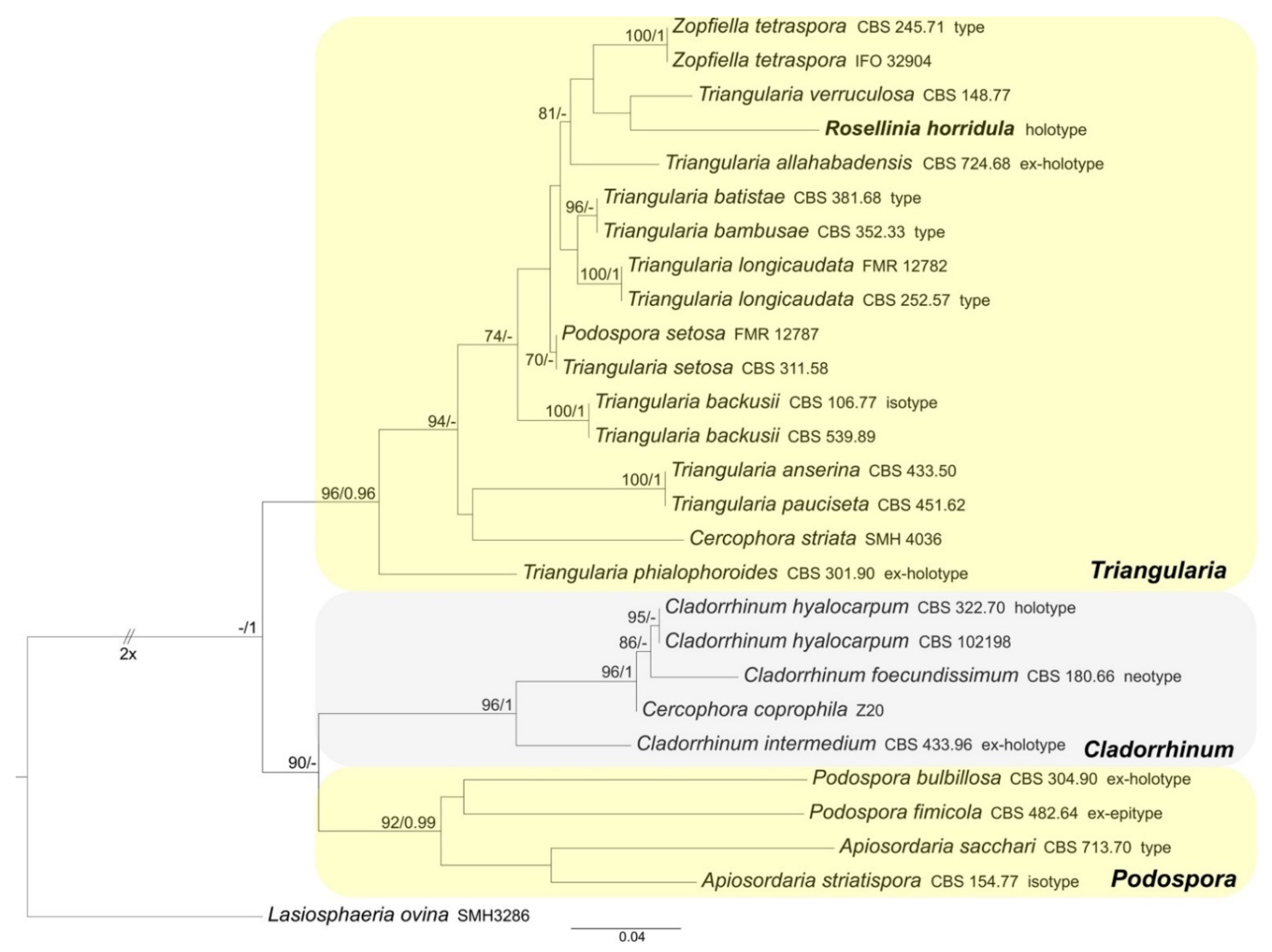
3.1.2. Podosporaceae
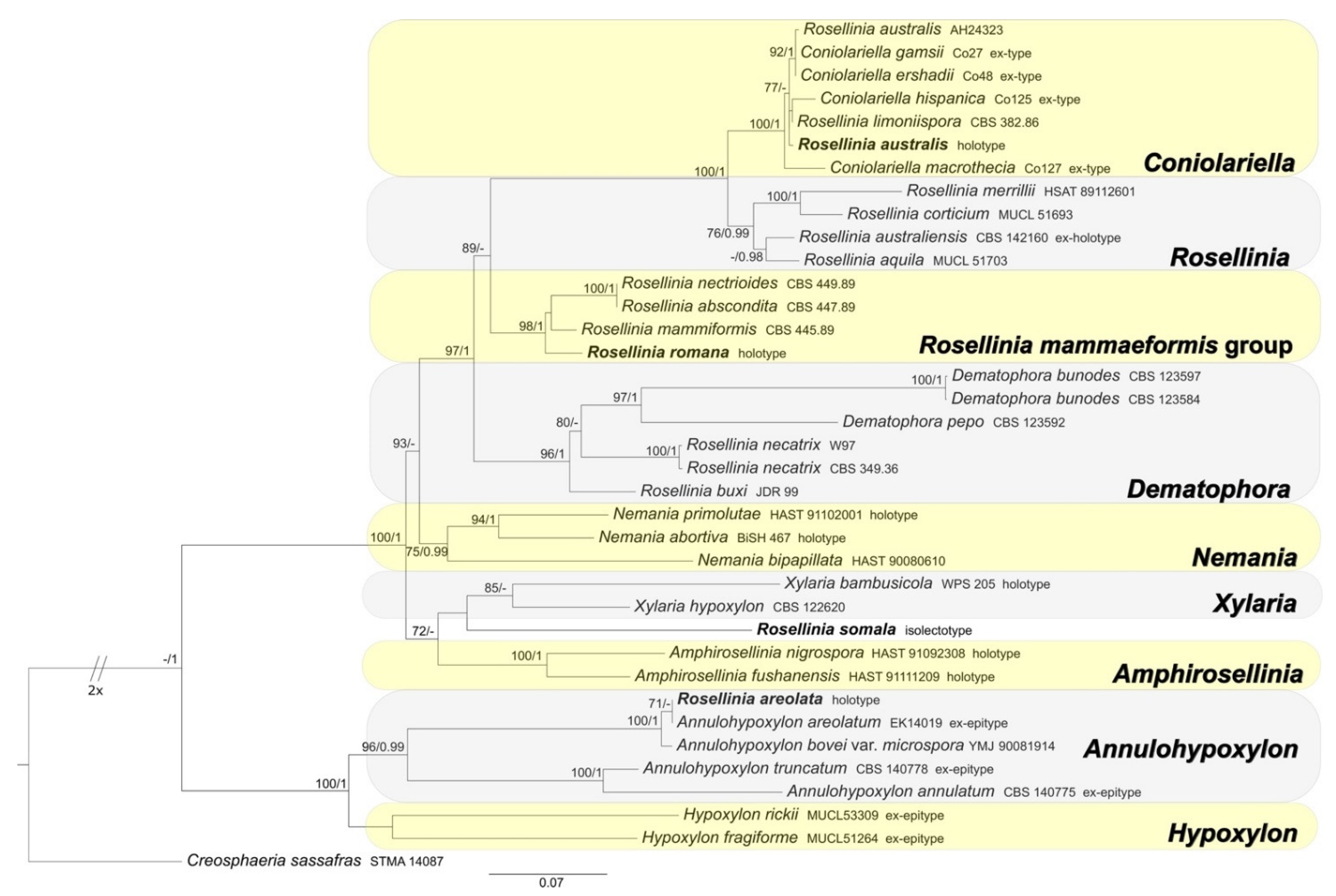
3.1.3. Xylariaceae and Hypoxylaceae
3.2. Taxonomy
3.2.1. Rosellinia ambigua
3.2.2. Rosellinia areolata
3.2.3. Rosellinia australis
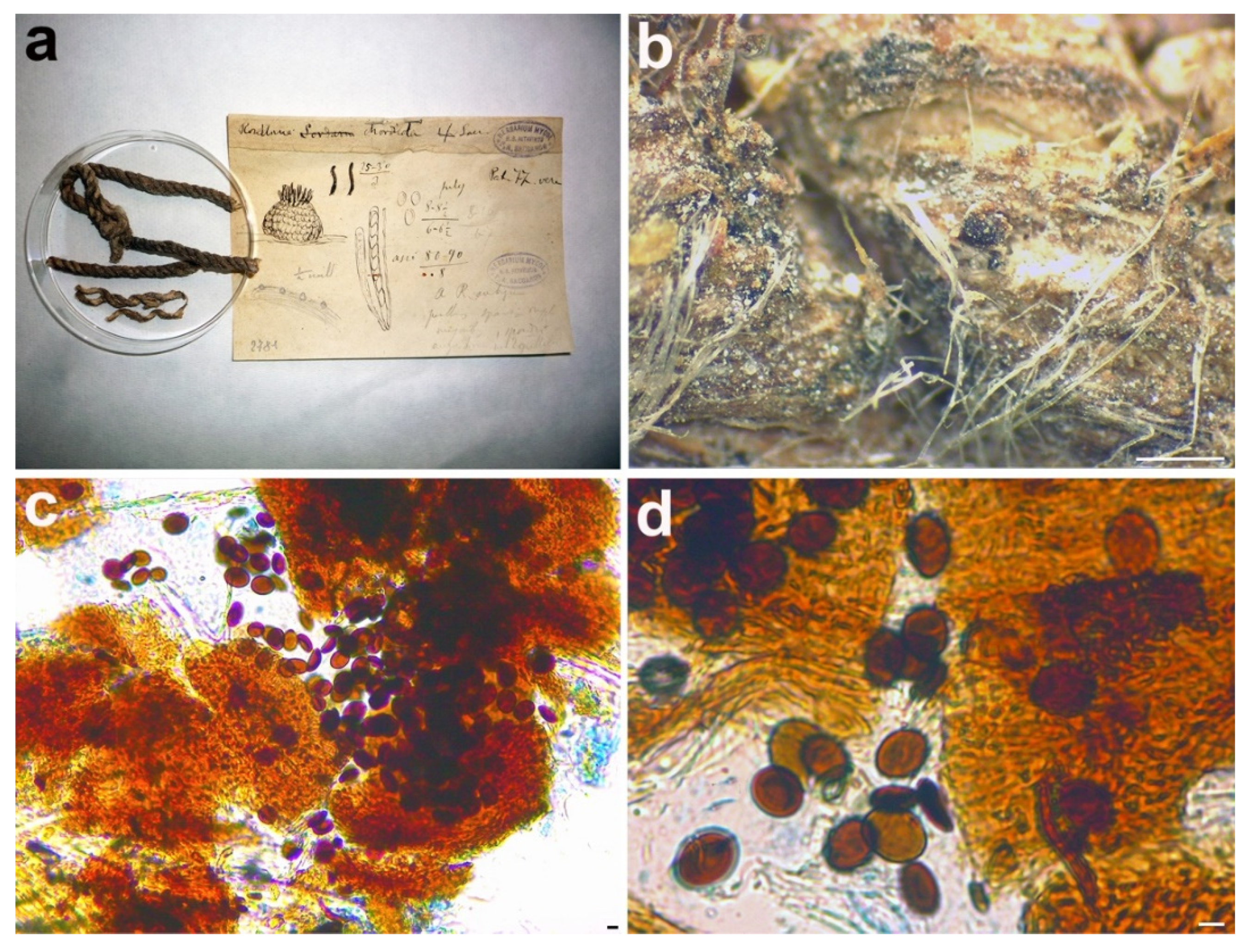
3.2.4. Rosellinia chordicola
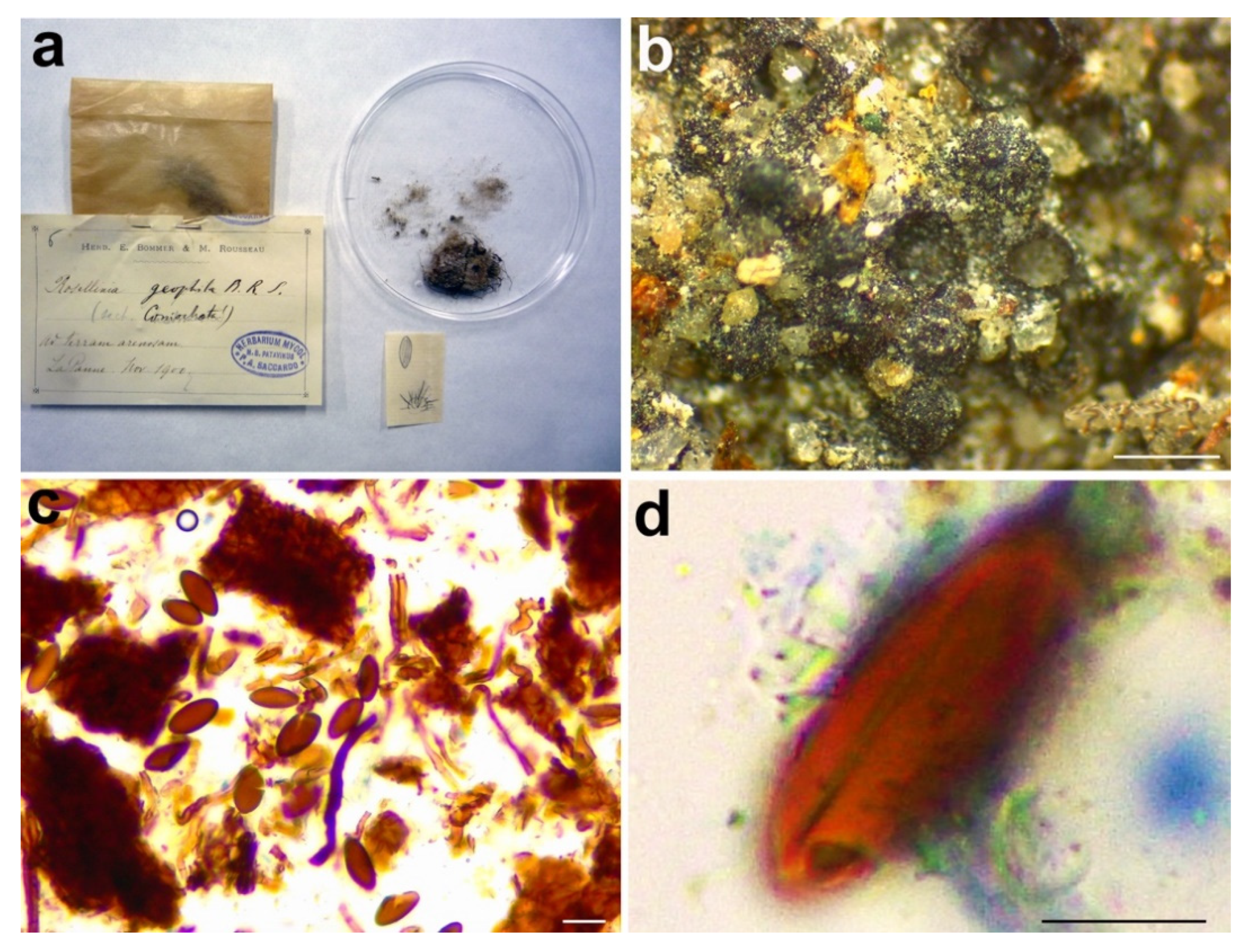
3.2.5. Rosellinia geophila
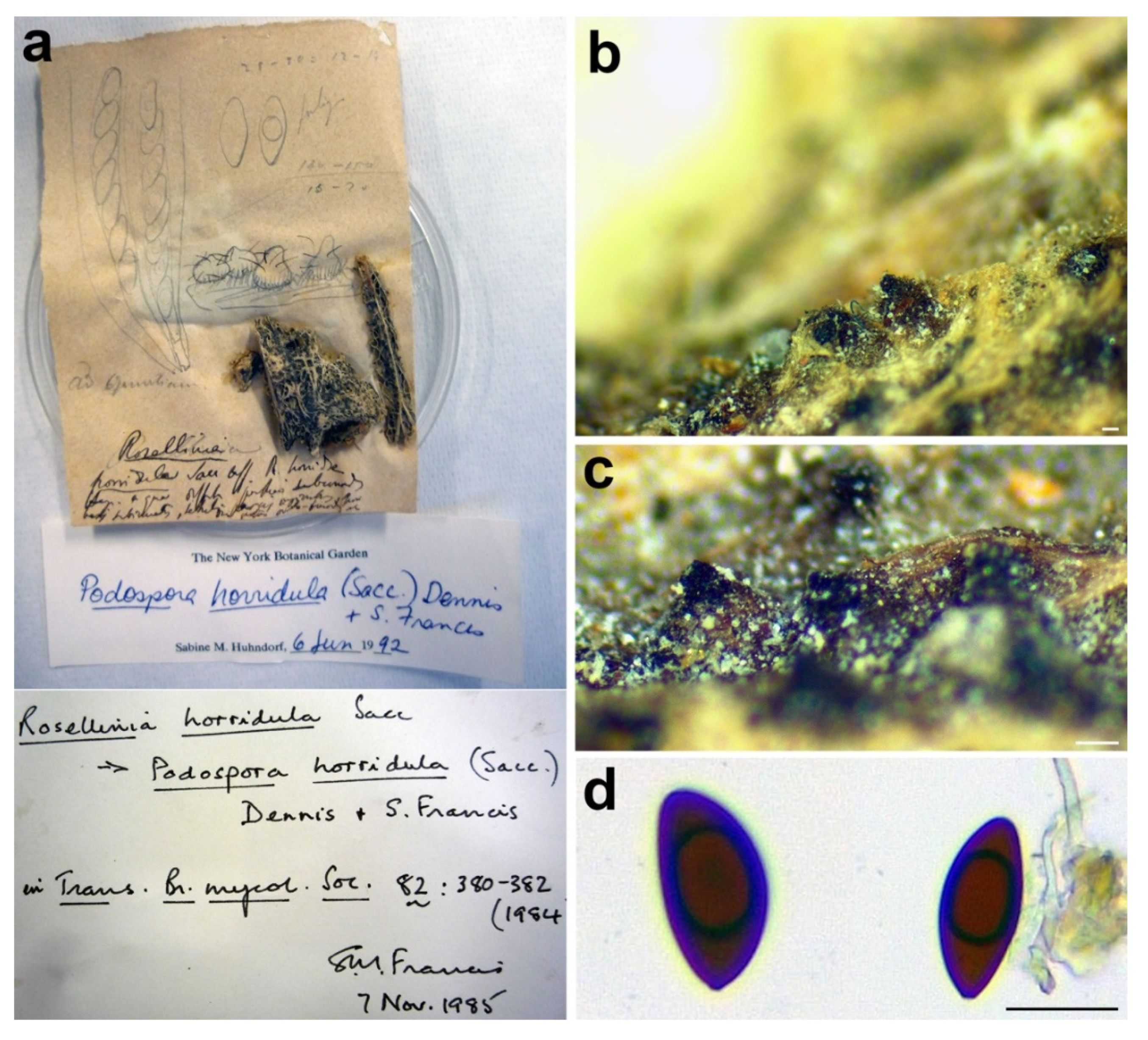
3.2.6. Rosellinia horridula
3.2.7. Rosellinia romana
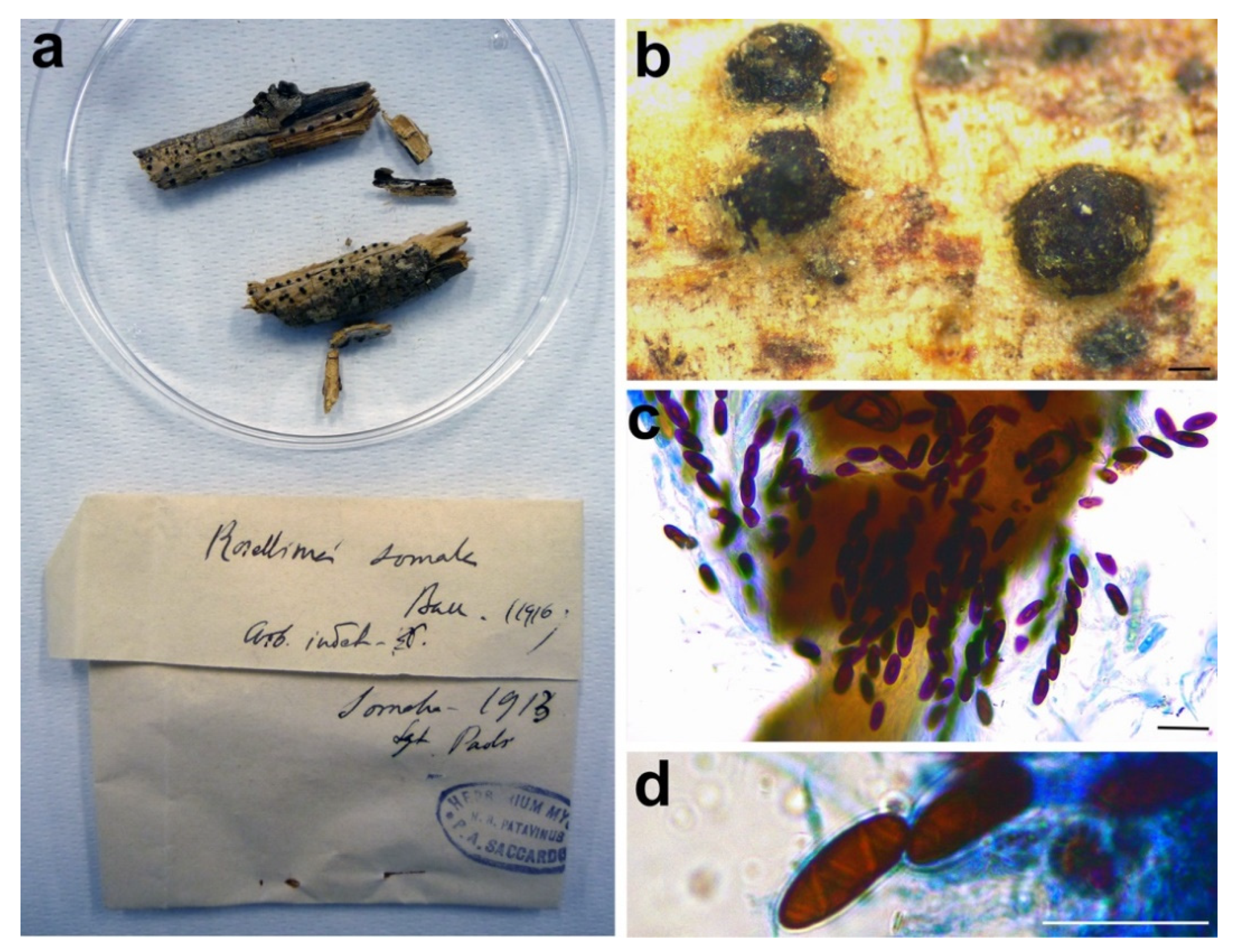
3.2.8. Rosellinia somala
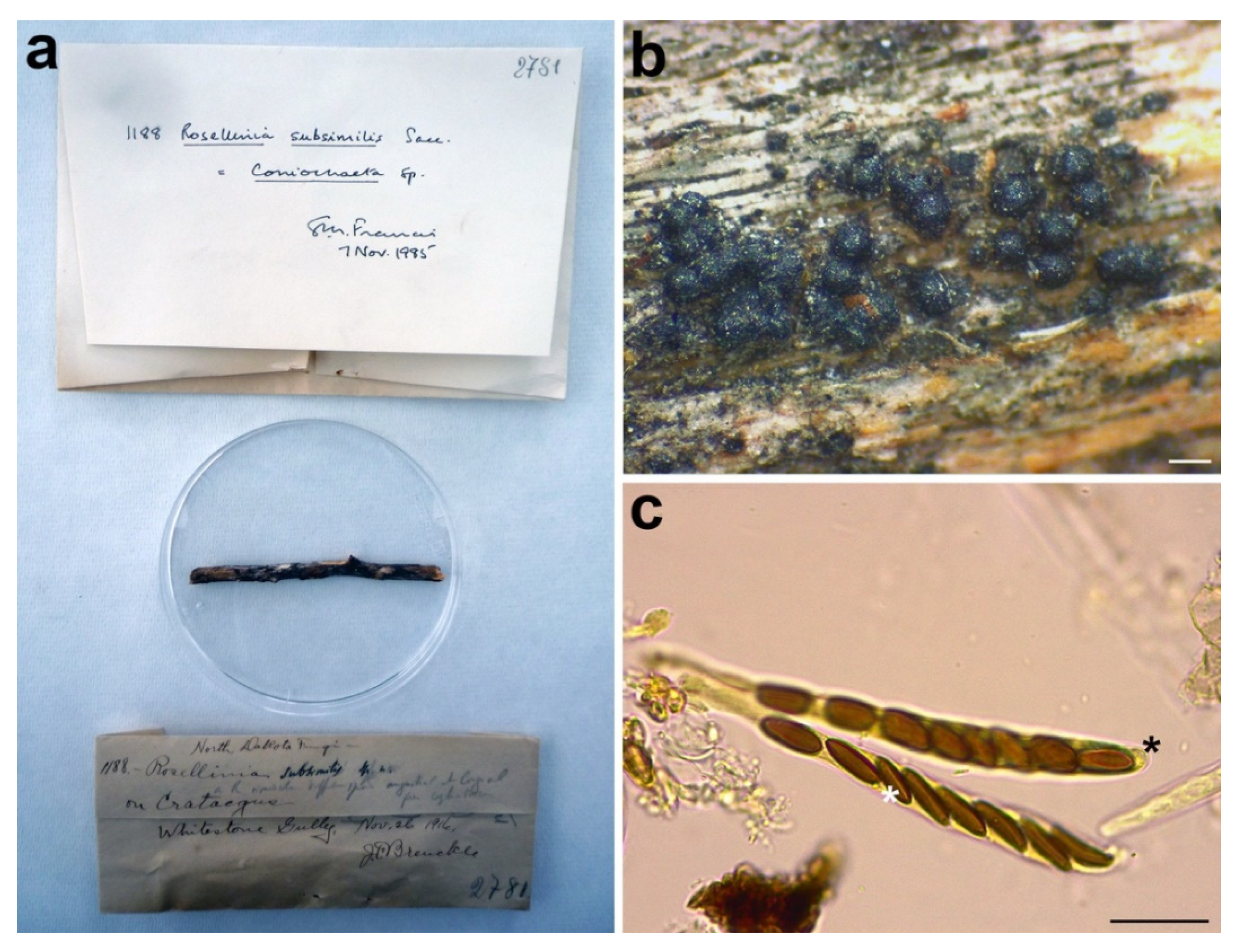
3.2.9. Rosellinia subsimilis
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Notaris, G. Cenni sulla tribù dei pirenomiceti sferiacei e descrizione di alcuni generi spettanti alla medesima. Giornale Botanico Italiano 1844, 1, 322–355. [Google Scholar]
- Petrini, L.E. Rosellinia species of the temperate zones. Sydowia 1992, 44, 169–281. [Google Scholar]
- Petrini, L.E. Rosellinia—A World Monograph. In Bibliotheca Mycologica, 1st ed.; J. Cramer: Stuttgart, DE, USA, 2013; Volume 205. [Google Scholar]
- Wittstein, K.; Cordsmeier, A.; Lambert, C.; Wendt, L.; Sir, E.B.; Weber, J.; Wurzler, N.; Petrini, L.E.; Stadler, M. Identification of Rosellinia species as producers of cyclodepsipeptide PF1022 A and resurrection of the genus Dematophora as inferred from polythetic taxonomy. Stud. Mycol. 2020, 96, 1–16. [Google Scholar] [CrossRef]
- Miller, J. Biologic studies in the Sphaeriales. Mycologia 1928, 20, 305–339. [Google Scholar]
- Wendt, L.; Sir, E.B.; Kuhnert, E.; Heitkämper, S.; Lambert, C.; Hladki, A.I.; Romero, A.I.; Luangsa-ard, J.J.; Srikitikulchai, P.; Peršoh, D.; et al. Resurrection and emendation of the Hypoxylaceae, recognised from a multigene phylogeny of the Xylariales. Mycol. Prog. 2018, 17, 115–154. [Google Scholar] [CrossRef] [Green Version]
- Saccardo, P.A. Sphaeriaceae, Phaeosporae, Rosellinia. Sylloge Fungorum 1882, 1, 252–277. [Google Scholar]
- Gola, G. L’erbario Micologico di P.A. Saccardo; Antoniana: Padova, Italy, 1930; pp. 214–215. [Google Scholar]
- Cooke, M.C. Synopsis pyrenomycetum. Grevillea 1887, 16, 16–19. [Google Scholar]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Forin, N.; Nigris, S.; Voyron, S.; Girlanda, M.; Vizzini, A.; Casadoro, G.; Baldan, B. Next generation sequencing of ancient fungal specimens: The case of the Saccardo mycological herbarium. Front. Ecol. Evol. 2018, 6, 129. [Google Scholar] [CrossRef] [Green Version]
- Forin, N.; Vizzini, A.; Nigris, S.; Ercole, E.; Voyron, S.; Girlanda, M.; Baldan, B. Illuminating type collections of nectriaceous fungi in Saccardo’s fungarium. Persoonia 2020, 45, 221–249. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications, 1st ed.; Innis, M.A., Gelfand, D.H., Sninsky, J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. Peer J. 2016, 4, e2584. [Google Scholar] [CrossRef]
- Bengtsson-Palme, J.; Ryberg, M.; Hartmann, M.; Branco, S.; Wang, Z.; Godhe, A.; Wit, P.D.; Sa, M.; Amend, A.S.; Jumpponen, A.; et al. Improved software detection and extraction of ITS1 and ITS2 from ribosomal ITS sequences of fungi and other eukaryotes for analysis of environmental sequencing data. Methods Ecol. Evol. 2013, 6. [Google Scholar] [CrossRef]
- UNITE Community. Full UNITE+INSD Dataset for Fungi. Version 18.11.2018; 2019. [Google Scholar] [CrossRef]
- Altschul, S. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A fast, scalable and user-friendly tool for maximum likelihood phylogenetic inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: A community resource for phylogenetic analyses. In TeraGrid Conference: Extreme Digital Discovery; San Diego Supercomputer Center: San Diego, CA, USA, 2011; Volume 41, pp. 1–8. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Vu, D.; Groenewald, M.; de Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Marin-Felix, Y.; Miller, A.N.; Cano-Lira, J.F.; Guarro, J.; García, D.; Stadler, M.; Huhndorf, S.M.; Stchigel, A.M. Re-evaluation of the order Sordariales: Delimitation of Lasiosphaeriaceae s. str., and introduction of the new families Diplogelasinosporaceae, Naviculisporaceae, and Schizotheciaceae. Microorganisms 2020, 8, 1430. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Han, T.; Li, W.; Jia, M.; Xue, L.; Rahman, K.; Qin, L. Geographic and tissue influences on endophytic fungal communities of Taxus chinensis var. mairei in China. Curr. Microbiol. 2013, 66, 40–48. [Google Scholar] [CrossRef]
- Réblová, M.; Winka, K. Phylogeny of Chaetosphaeria and its anamorphs based on morphological and molecular data. Mycolgia 2000, 92, 939–954. [Google Scholar] [CrossRef]
- Wang, X.W.; Bai, F.Y.; Bensch, K.; Meijer, M.; Sun, B.D.; Han, Y.F.; Crous, P.W.; Samson, R.A.; Yang, F.Y.; Houbraken, J. Phylogenetic re-evaluation of Thielavia with the introduction of a new family Podosporaceae. Stud. Mycol. 2019, 93, 155–252. [Google Scholar] [CrossRef]
- Samarakoon, M.C.; Gafforov, Y.; Liu, N.; Maharachchikumbura, S.S.N.; Bhat, J.D.; Liu, J.-K.; Promputtha, I.; Hyde, K.D. Combined multi-gene backbone tree for the genus Coniochaeta with two new species from Uzbekistan. Phytotaxa 2018, 336, 43–56. [Google Scholar] [CrossRef]
- Damm, U.; Fourie, P.H.; Crous, P.W. Coniochaeta (Lecythophora), Collophora gen. nov. and Phaeomoniella species associated with wood necroses of Prunus trees. Persoonia 2010, 24, 60–80. [Google Scholar] [CrossRef] [Green Version]
- Dayarathne, M.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Devadatha, B.; Sarma, V.V.; Khongphinitbunjong, K.; Chomnunti, P.; Hyde, K.D. Morpho-molecular characterization of microfungi associated with marine based habitats. Mycosphere 2020, 11, 1–188. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Phukhamsakda, C.; Hyde, K.D.; Jeewon, R.; Lee, H.B.; Jones, G.E.B.; Tibpromma, S.; Tennakoon, D.S.; Dissanayake, A.J.; Jayasiri, S.C.; et al. Fungal Diversity Notes 709–839: Taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N.; et al. Finding needles in haystacks: Linking scientific names, reference specimens and molecular data for Fungi. Database Oxford 2014. [Google Scholar] [CrossRef]
- Han, J.; Liu, C.; Li, L.; Zhou, H.; Liu, L.; Bao, L.; Chen, Q.; Song, F.; Zhang, L.; Li, E.; et al. Decalin-containing tetramic acids and 4-Hydroxy-2-Pyridones with antimicrobial and cytotoxic activity from the fungus Coniochaeta cephalothecoides collected in Tibetan Plateau (Medog). J. Org. Chem. 2017, 82, 11474–11486. [Google Scholar] [CrossRef]
- Coronado-Ruiz, C.; Avendaño, R.; Escudero-Leyva, E.; Conejo-Barboza, G.; Chaverri, P.; Chavarría, M. Two new cellulolytic fungal species isolated from a 19th-Century art collection. Sci. Rep. 2018, 8, 7492. [Google Scholar] [CrossRef] [Green Version]
- Perdomo, H.; García, D.; Gené, J.; Cano, J.; Sutton, D.A.; Summerbell, R.; Guarro, J. Phialemoniopsis, a new genus of Sordariomycetes, and new species of Phialemonium and Lecythophora. Mycologia 2013, 105, 398–421. [Google Scholar] [CrossRef]
- Harrington, A.H.; Del Olmo-Ruiz, M.; U’Ren, J.M.; Garcia, K.; Pignatta, D.; Wespe, N.; Sandberg, D.C.; Huang, Y.-L.; Hoffman, M.T.; Arnold, A.E. Coniochaeta endophytica sp. nov., a foliar endophyte associated with healthy photosynthetic tissue of Platycladus orientalis (Cupressaceae). Plant Fungal Syst. 2019, 64, 65–79. [Google Scholar] [CrossRef] [Green Version]
- Nasr, S.; Bien, S.; Soudi, M.R.; Alimadadi, N.; Shahzadeh Fazeli, S.A.; Damm, U. Novel Collophorina and Coniochaeta species from Euphorbia polycaulis, an endemic plant in Iran. Mycol. Prog. 2018, 17, 755–771. [Google Scholar] [CrossRef]
- Vázquez-Campos, X.; Kinsela, A.S.; Waite, T.D.; Collins, R.N.; Neilan, B.A. Fodinomyces uranophilus gen. nov. sp. nov. and Coniochaeta fodinicola sp. nov., two uranium mine-inhabiting Ascomycota fungi from northern Australia. Mycologia 2014, 106, 1073–1089. [Google Scholar] [CrossRef]
- Leonhardt, S.; Büttner, E.; Gebauer, A.M.; Hofrichter, M.; Kellner, H. Draft Genome Sequence of the Sordariomycete Lecythophora (Coniochaeta) hoffmannii CBS 245.38. Genome Announc. 2018, 6, e01510-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, E.B.G.; Devadatha, B.; Abdel-Wahab, M.A.; Dayarathne, M.C.; Zhang, S.-N.; Hyde, K.D.; Liu, J.-K.; Bahkali, A.H.; Sarma, V.V.; Tibell, S.; et al. Phylogeny of new marine Dothideomycetes and Sordariomycetes from mangroves and deep-sea sediments. Bot. Mar. 2020, 63, 155–181. [Google Scholar] [CrossRef] [Green Version]
- Friebes, G.; Jaklitsch, W.M.; García, S.; Voglmayr, H. Lopadostoma taeniosporum revisited and a new species of Coniochaeta. Sydowia 2016, 68, 87–97. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef] [Green Version]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.-G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.; et al. Fungal Diversity Notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef] [Green Version]
- Miller, A.N.; Huhndorf, S.M. Using phylogenetic species recognition to delimit species boundaries within Lasiosphaeria. Mycologia 2004, 96, 1106. [Google Scholar] [CrossRef]
- Hsieh, H.-M.; Lin, C.-R.; Fang, M.-J.; Rogers, J.D.; Fournier, J.; Lechat, C.; Ju, Y.-M. Phylogenetic status of Xylaria subgenus Pseudoxylaria among taxa of the subfamily Xylarioideae (Xylariaceae) and phylogeny of the taxa involved in the subfamily. Mol. Phylogenet. Evol. 2010, 54, 957–969. [Google Scholar] [CrossRef]
- Kuhnert, E.; Sir, E.B.; Lambert, C.; Hyde, K.D.; Hladki, A.I.; Romero, A.I.; Rohde, M.; Stadler, M. Phylogenetic and chemotaxonomic resolution of the genus Annulohypoxylon (Xylariaceae) including four new species. Fungal Divers. 2017, 85, 1–43. [Google Scholar] [CrossRef]
- Hsieh, H.-M.; Ju, Y.-M.; Rogers, J.D. Molecular phylogeny of Hypoxylon and closely related genera. Mycologia 2005, 97, 844–865. [Google Scholar] [CrossRef]
- Zare, R.; Asgari, B.; Gams, W. The species of Coniolariella. Mycologia 2010, 102, 1383–1388. [Google Scholar] [CrossRef]
- Daranagama, D.A.; Camporesi, E.; Tian, Q.; Liu, X.; Chamyuang, S.; Stadler, M.; Hyde, K.D. Anthostomella is polyphyletic comprising several genera in Xylariaceae. Fungal Divers. 2015, 73, 203–238. [Google Scholar] [CrossRef]
- Kuhnert, E.; Fournier, J.; Per, D.; Luangsa-ard, J.J.D.; Stadler, M. New Hypoxylon species from Martinique and new evidence on the molecular phylogeny of Hypoxylon based on ITS rDNA and β-Tubulin data. Fungal Divers. 2014, 64, 181–203. [Google Scholar] [CrossRef]
- Ju, Y.-M.; Hsieh, H.-M.; Ho, M.-C.; Szu, D.-H.; Fang, M.-J. Theissenia rogersii sp. nov. and phylogenetic position of Theissenia. Mycologia 2007, 99, 612–621. [Google Scholar] [CrossRef] [PubMed]
- Vicente, F.; Basilio, A.; Platas, G.; Collado, J.; Bills, G.F.; González Del Val, A.; Martín, J.; Tormo, J.R.; Harris, G.H.; Zink, D.L.; et al. Distribution of the antifungal agents aordarins across filamentous fungi. Mycol. Res. 2009, 113, 754–770. [Google Scholar] [CrossRef]
- Peláez, F.; González, V.; Platas, G.; Sánchez-Ballesteros, J.; Rubio, V. Molecular phylogenetic studies within the Xylariaceae based on ribosomal DNA sequences. Fungal Divers. 2008, 31, 111–134. [Google Scholar]
- Checa, J.; Arenal, F.; Blanco, N.; Rogers, J.D. Coniolariella hispanica sp. nov. and other additions to Coniolariella. Mycol. Res. 2008, 112, 795–801. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Guarro, J.; Cheewangkoon, R.; van der Bank, M.; Swart, W.J.; Stchigel, A.M.; Cano-Lira, J.F.; Roux, J.; Madrid, H.; et al. Fungal Planet Description Sheets: 154–213. Persoonia 2013, 31, 188–296. [Google Scholar] [CrossRef]
- Sir, E.; Lambert, C.; Wendt, L.; Hladki, A.I.; Romero, A.I.; Stadler, M. A new species of Daldinia (Xylariaceae) from the Argentine subtropical montane forest. Mycosphere 2016, 7, 1378–1388. [Google Scholar] [CrossRef]
- Popuschoi, I.S. Mikoflora Plodorykh Derev’ev SSSR; 1971; p. 90. [Google Scholar]
- Kamiya, S.; Uchiyama, S.; Udagawa, S.-I. Two new species of Coniochaeta with a cephalothecoid peridium wall. Mycoscience 1995, 36, 377–383. [Google Scholar] [CrossRef]
- García, D.; Stchigel, A.M.; Cano, J.; Calduch, M.; Hawksworth, D.L.; Guarro, J. Molecular phylogeny of Coniochaetales. Mycol. Res. 2006, 110, 1271–1289. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Gené, J.; Ahmad, S.; Cano, J.; Al-Sweih, N.; Joseph, L.; Chandy, R.; Guarro, J. Coniochaeta polymorpha, a new species from endotracheal aspirate of a preterm neonate, and transfer of Lecythophora species to Coniochaeta. Antonie Van Leeuwenhoek 2013, 104, 243–252. [Google Scholar] [CrossRef] [PubMed]
- García, D.; Stchigel, A.M.; Guarro, J. A new species of Poroconiochaeta from Russian soils. Mycologia 2003, 95, 525–529. [Google Scholar] [CrossRef]
- Weber, E. The Lecythophora-Coniochaeta complex I. Morphological studies on Lecythophora species isolated from Picea abies. Nova Hedwigia 2002, 74, 159–185. [Google Scholar] [CrossRef]
- Mahoney, D.P.; LaFavre, J.S. Coniochaeta extramundana, with a synopsis of other Coniochaeta species. Mycologia 1981, 73, 931–952. [Google Scholar] [CrossRef]
- Checa, J.; Barrasa, J.M.; Moreno, G.; Fort, F.; Guarro, J. The genus Coniochaeta (Sacc.) Cooke (Coniochaetaceae, Ascomycotina) in Spain. Cryptog. Mycol. 1988, 9, 1–34. [Google Scholar]
- Francis, S.M.; Sparrow, J.R. Podospora horridula. Trans. Br. Mycol. Soc. 1984, 82, 380–382. [Google Scholar] [CrossRef]
- Hawksworth, D.L.; Lodha, B.C. Helicogermslita, a new stromatic xylariaceous genus with a spiral germ slit from India. Trans. Br. Mycol. Soc. 1983, 81, 91–96. [Google Scholar] [CrossRef]
- Laessöe, T.; Spooner, B.M. Rosellinia & Astrocystis (Xylariaceae): New species and generic concepts. Kew Bull. 1993, 49, 1–70. [Google Scholar] [CrossRef]
- Petrini, L.E. Rosellinia and related genera in New Zealand. N. Zeal. J. Bot. 2003, 41, 71–138. [Google Scholar] [CrossRef]
- Lee, S.; Crous, P.W. A new species of Helicogermslita from South Africa. Sydowia 2003, 55, 109–114. [Google Scholar]
- Li, Q.; Kang, J.; Hyde, K.D. Two new Rosellinia species from southwest China. Mycotaxon 2015, 130, 563–567. [Google Scholar] [CrossRef]
- Seifert, K.A.; Rossman, A.Y. How to describe a new fungal species. IMA Fungus 2010, 1, 109–111. [Google Scholar] [CrossRef] [PubMed]
- Lücking, R.; Aime, M.C.; Robbertse, B.; Miller, A.N.; Ariyawansa, H.A.; Aoki, T.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Unambiguous identification of fungi: Where do we stand and how accurate and precise is fungal DNA barcoding? IMA Fungus 2020, 11, 1–32. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GenBank Accession Numbers | |||||
|---|---|---|---|---|---|
| Original Identification | Name to Be Used | Herbarium/Strain | ITS1 | ITS2 | ITS |
| Apiosordaria sacchari | Podospora sacchari | CBS 713.70, type of Echinopodospora sacchari [26] | - | - | MH859915 |
| Apiosordaria striatispora | Podospora striatispora | CBS 154.77, isotype of Triangularia striatispora [27] | - | - | MT784137 |
| Arnium arizonense | Triangularia arizonensis | CBS 120289 | - | - | KU955584 |
| Cercophora coprophila | Cladorrhinum coprophilum | Z20 [28] | - | - | JN198495 |
| Cercophora striata | Triangularia striata | SMH 4036 | - | - | KX348038 |
| Chaetosphaeria pygmaea | Chaetosphaeria pygmaea | MR 1365 [29] | - | - | AF178545 |
| Cladorrhinum foecundissimum | Cladorrhinum foecundissimum | CBS 180.66, neotype [30] | - | - | MK926856 |
| Cladorrhinum hyalocarpum | Cladorrhinum hyalocarpum | CBS 322.70, holotype of Thielavia hyalocarpa [30] | - | - | MK926857 |
| CBS 102198 [30] | - | - | MK926858 | ||
| Cladorrhinum intermedium | Cladorrhinum intermedium | CBS 433.96, ex-holotype of Thielavia intermedia [30] | - | - | MK926859 |
| Coniochaeta acaciae | Coniochaeta acaciae | MFLUCC 17-2298, ex-holotype [31] | - | - | MG062735 |
| Coniochaeta africana | Coniochaeta africana | CBS 1200868, ex-holotype [32] | - | - | NR_137725 |
| Coniochaeta angustispora | Coniochaeta angustispora | CBS 872.73 [26] | - | - | MH860817 |
| Coniochaeta arenariae | Coniochaeta arenariae | MFLUCC 18-0409 [33] | - | - | MN047126 |
| Coniochaeta arxii | Coniochaeta arxii | CBS 192.89, isotype [26] | - | - | MH862164 |
| Coniochaeta baysunika | Coniochaeta baysunika | MFLUCC 17-0830, holotype [34] | - | - | NR_157508 |
| Coniochaeta boothii | Coniochaeta boothii | CBS 381.74, type of Thielavia boothii [26] | - | - | NR_159776 |
| Coniochaeta canina | Coniochaeta canina | CBS 133243, ex-holotype of Lecythophora canina [35] | - | - | NR_120211 |
| Coniochaeta cateniformis | Coniochaeta cateniformis | CBS 131709, holotype of Lecythophora cateniformis [26] | - | - | MH865902 |
| Coniochaeta cephalothecoides | Coniochaeta cephalothecoides | L821 [36] | - | - | KY064029 |
| Coniochaeta cipronana | Coniochaeta cipronana | CBS 144016, holotype [37] | - | - | NR_157478 |
| Coniochaeta coluteae | Coniochaeta coluteae | MFLUCC 17-2299, ex-holotype [31] | - | - | MG137251 |
| Coniochaeta cymbiformispora | Coniochaeta cymbiformispora | NBRC 32199, ex-holotype | - | - | LC146726 |
| Coniochaeta cypraeaespora | Coniochaeta cypraeaespora | CBS 365.93, type [26] | - | - | MH862420 |
| Coniochaeta decumbens | Coniochaeta decumbens | CBS 153.42, holotype of Margarinomyces decumbens [38] | - | - | NR_144912 |
| Coniochaeta discoidea | Coniochaeta discoidea | CBS 158.80, type of Poroconiochaeta discoidea [26] | - | - | NR_159779 |
| Coniochaeta discospora | Coniochaeta discospora | CBS 168.58, type of Sordaria discospora [26] | - | - | MH857740 |
| Coniochaeta ellipsoidea | Coniochaeta ellipsoidea | CBS 137.68, type [26] | - | - | MH859091 |
| Coniochaeta endophytica | Coniochaeta endophytica | AEA 9094, type [39] | - | - | EF420005 |
| Coniochaeta euphorbiae | Coniochaeta euphorbiae | CBS 139768, ex-type [40] | - | - | KP941076 |
| Coniochaeta extramundana | Coniochaeta extramundana | CBS 247.77, type [26] | - | - | MH861057 |
| Coniochaeta fasciculata | Coniochaeta fasciculata | CBS 205.38, holotype of Margarinomyces fasciculatus [38] | - | - | NR_154770 |
| Coniochaeta fodinicola | Coniochaeta fodinicola | CBS 136963, holotype [41] | - | - | JQ904603 |
| Coniochaeta gigantospora | Coniochaeta gigantospora | ILLS 60816, type [35] | - | - | NR_121521 |
| Coniochaeta hoffmannii | Coniochaeta hoffmannii | CBS 245.38, holotype of Margarinomyces hoffmannii [42] | - | - | NR_167688 |
| Coniochaeta iranica | Coniochaeta iranica | CBS 139767 [40] | - | - | KP941078 |
| Coniochaeta ligniaria | Coniochaeta ligniaria | CBS 424.65 [26] | - | - | MH858650 |
| Coniochaeta lignicola | Coniochaeta lignicola | CBS 267.33, lectotype of Lecythophora lignicola [35] | - | - | NR_111520 |
| Coniochaeta luteorubra | Coniochaeta luteorubra | CBS 131710, holotype Lecythophora luteorubra [26] | - | - | MH865901 |
| Coniochaeta luteoviridis | Coniochaeta luteoviridis | CBS 206.38, holotype of Margarinomyces luteoviridis [38] | - | - | NR_154769 |
| Coniochaeta marina | Coniochaeta marina | MFLUCC 18-0408, ex-holotype [43] | - | - | MK458764 |
| Coniochaeta mutabilis | Coniochaeta mutabilis | CBS 157.44, holotype of Margarinomyces mutabilis [35] | - | - | NR_111519 |
| Coniochaeta navarrae | Coniochaeta navarrae | CBS 141016, ex-holotype [44] | - | - | NR_154808 |
| Coniochaeta nepalica | Coniochaeta nepalica | NBRC 30584, ex-type | - | - | LC146727 |
| Coniochaeta ostrea | Coniochaeta ostrea | CBS 507.70, type of Coniochaetidium ostreum [26] | - | - | NR_159772 |
| Coniochaeta polymorpha | Coniochaeta polymorpha | CBS 132722, holotype [35] | - | - | NR_121473 |
| Coniochaeta polysperma | Coniochaeta polysperma | CBS 669.77, isotype [26] | - | - | MH861109 |
| Coniochaeta prunicola | Coniochaeta prunicola | CBS 120875, holotype [32] | - | - | NR_137037 |
| Coniochaeta punctulata | Coniochaeta punctulata | CBS 159.80, isotype of Poroconiochaeta punctulata [26] | - | - | MH861254 |
| Coniochaeta rosae | Coniochaeta rosae | MFLUCC 17-0810, holotype [34] | - | - | NR_157509 |
| Coniochaeta savoryi | Coniochaeta savoryi | CBS 725.74, isotype of Thielavia savoryi [26] | - | - | MH860890 |
| Coniochaeta simbalensis | Coniochaeta simbalensis | NFCCI 4236, ex-holotype [45] | - | - | NR_164024 |
| Coniochaeta taeniospora | Coniochaeta taeniospora | CBS 141014, ex-epitype [44] | - | - | KU762324 |
| Coniochaeta tetraspora | Coniochaeta tetraspora | CBS 139.68 [26] | - | - | MH859093 |
| Coniochaeta velutina | Coniochaeta velutina | CBS 120874 [32] | - | - | GQ154542 |
| Coniochaeta verticillata | Coniochaeta verticillata | CBS 816.71, holotype of Ephemeroascus verticillatus [26] | - | - | NR_159774 |
| Coniochaeta vineae | Coniochaeta vineae | KUMCC 17-0322, ex-holotype [46] | - | - | MN473469 |
| Lasiosphaeria ovina | Lasiosphaeria ovina | SMH3286 [47] | - | - | AY587931 |
| Podospora bulbillosa | Podospora bulbillosa | CBS 304.90, ex-holotype of Cladorrhinum bulbillosum [30] | - | - | MK926861 |
| Podospora fimicola | Podospora fimicola | CBS 482.64, ex-epitype of Schizothecium fimicola [30] | - | - | MK926862 |
| Podospora setosa | Triangularia setosa | FMR 12787 [27] | - | - | MT784144 |
| Rosellinia ambigua | Coniochaeta ambigua | PADS00027: fungarium Saccardo, holotype | MW626895 | MW626903 | - |
| Rosellinia chordicola | Coniochaeta chordicola (Sacc.) Forin, Fainelli & Vizzini | PAD S00030: fungarium Saccardo, holotype | MW626898 | MW626905 | - |
| Rosellinia geophila | Coniochaeta geophila (E. Bommer, M. Rousseau & Sacc.) Forin, Fainelli & Vizzini | PAD S00031: fungarium Saccardo, holotype | - | MW626906 | - |
| Rosellinia horridula | Triangularia horridula (Sacc.) Forin, Fainelli & Vizzini | PAD S00032: fungarium Saccardo, holotype | MW626899 | MW626907 | - |
| Rosellinia subsimilis | Coniochaeta dakotensis Forin, Fainelli & Vizzini | PAD S00035: fungarium Saccardo, holotype | MW626902 | MW626910 | - |
| Triangularia allahabadensis | Triangularia allahabadensis | CBS 724.68, ex-holotype of Sordaria allahabadensis [30] | - | - | MK926865 |
| Triangularia anserina | Triangularia anserina | CBS 433.50 [30] | - | - | MK926864 |
| Triangularia backusii | Triangularia backusii | CBS 539.89, isotype [30] | - | - | MK926866 |
| CBS 106.77 [30] | - | - | MK926867 | ||
| Triangularia bambusae | Triangularia bambusae | CBS 352.33, type of Trigonia bambusae [30] | - | - | MK926868 |
| Triangularia batistae | Triangularia batistae | CBS 381.68, type [27] | - | - | MT784140 |
| Triangularia longicaudata | Triangularia longicaudata | CBS 252.57, type [30] | - | - | MK926869 |
| FMR 12782 [27] | - | - | MT784142 | ||
| Triangularia pauciseta | Triangularia pauciseta | CBS 451.62 [30] | - | - | MK926870 |
| Triangularia phialophoroides | Triangularia phialophoroides | CBS 301.90, ex-holotype of Cladorrhinum phialophoroides [30] | - | - | MK926871 |
| Triangularia setosa | Triangularia setosa | CBS 311.58 [30] | - | - | MK926872 |
| Triangularia verruculosa | Triangularia verruculosa | CBS 148.77 [30] | - | - | MK926874 |
| Zopfiella tetraspora | Triangularia tetraspora | CBS 245.71, type of Podospora buffonii [26] | - | - | MH860097 |
| IFO 32904 | - | - | AY999130 | ||
| GenBank Accession Numbers | |||||||
|---|---|---|---|---|---|---|---|
| Original Identification | Name to Be Used | Herbarium/Strain | ITS1 | ITS2 | ITS | LSU | TUB2 |
| Amphirosellinia fushanensis | Amphirosellinia fushanensis | HAST 91111209, holotype [48] | - | - | GU339496 | - | GQ495950 |
| Amphirosellinia nigrospora | Amphirosellinia nigrospora | HAST 91092308, holotype [48] | - | - | GU322457 | - | GQ495951 |
| Annulohypoxylon annulatum | Annulohypoxylon annulatum | CBS 140775, ex-epitype [6] | - | - | KY610418 | KY610418 | - |
| Annulohypoxylon areolatum | Annulohypoxylon areolatum | MFLUCC 14-1233, EK14019 ex-epitype [49] | - | - | KX376327 | - | KX376344 |
| Annulohypoxylon bovei var. microspora | Annulohypoxylon bovei var. microspora | YMJ 90081914 [48,50] | - | - | EF026141 | - | AY951654 |
| Annulohypoxylon truncatum | Annulohypoxylon truncatum | CBS 140778, ex-epitype [6,49] | - | - | KY610419 | KY610419 | KX376352 |
| Coniolariella ershadii | Coniolariella ershadii | Co48, CBS 119785, ex-type [51] | - | - | GU553328 | GU553331 | - |
| Coniolariella gamsii | Coniolariella gamsii | Co27, CBS 114379, ex-type [51] | - | - | GU553325 | GU553329 | - |
| Coniolariella hispanica | Coniolariella hispanica | Co125, CBS 124506, ex-type [51] | - | - | GU553323 | GU553353 | - |
| Coniolariella macrothecia | Coniolariella macrothecia | Co127, CBS 125772, ex-type [51] | - | - | GU553324 | GU553354 | - |
| Creosphaeria sassafras | Creosphaeria sassafras | STMA 14087 [6] | - | - | KY610411 | KY610468 | KX271258 |
| Dematophora bunodes | Dematophora bunodes | CBS 123584 [4] | - | - | MN984617 | - | MN987243 |
| CBS 123597 [4] | - | - | MN984618 | - | MN987244 | ||
| Dematophora pepo | Dematophora pepo | CBS 123592 [4] | - | - | MN984620 | - | MN987246 |
| Hypoxylon fragiforme | Hypoxylon fragiforme | MUCL51264, ex-epitype [52] | - | - | KM186294 | KM186295 | KM186293 |
| Hypoxylon rickii | Hypoxylon rickii | MUCL 53309, ex-epitype [53] | - | - | KC968932 | - | KC977288 |
| Nemania abortiva | Nemania abortiva | BiSH 467, holotype [48] | - | - | GU292816 | - | GQ470219 |
| Nemania bipapillata | Nemania bipapillata | HAST 90080610 [48] | - | - | GU292818 | - | GQ470221 |
| Nemania primolutae | Nemania primolutae | HAST 91102001 [48,54] | - | - | EF026121 | - | EF025607 |
| Rosellinia abscondita | Rosellinia abscondita | CBS 447.89 [55] | - | - | FJ175180 | KF719208 | - |
| Rosellinia aquila | Rosellinia aquila | MUCL 51703 [6] | - | - | KY610392 | KY610460 | KX271253 |
| Rosellinia areolata | Annulohypoxylon areolatum | PAD S00028: fungarium Saccardo, holotype | MW626896 | - | - | - | - |
| Rosellinia australis | Coniolariella limoniispora | PAD S00029: fungarium Saccardo, holotype | MW626897 | MW626904 | - | - | - |
| AH24323 [56,57] | - | - | AY908997 | EF489464 | - | ||
| Rosellinia australiensis | Rosellinia australiensis | CBS 142160, ex-holotype [58] | - | - | KY979742 | KY979797 | - |
| Rosellinia buxi | Dematophora buxi | JDR 99 [48] | - | - | GU300070 | - | GQ470228 |
| Rosellinia corticum | Rosellinia corticum | MUCL 51693 [6] | - | - | KY610393 | KY610461 | KX271254 |
| Rosellinia limonispora | Coniolariella limoniispora | CBS 382.86 | - | - | KF719199 | KF719211 | - |
| Rosellinia mammiformis | Rosellinia mammiformis | CBS 445.89 | - | - | KF719200 | KF719212 | - |
| Rosellinia merrillii | Rosellinia merrillii | HAST 89112601 [48] | - | - | GU300071 | - | GQ470229 |
| Rosellinia necatrix | Dematophora necatrix | CBS 349.36 [56] | - | - | AY909001 | KF719204 | KY624310 |
| W97 | - | - | DF977487 | DF977487 | DF977466 | ||
| Rosellinia nectrioides | Rosellinia nectrioides | CBS 449.89 [4] | - | - | MN984622 | MN984628 | - |
| Rosellinia romana | Rosellinia glabra | PAD S00033: fungarium Saccardo, holotype | MW626900 | MW626908 | - | - | - |
| Rosellinia somala | Helicogermslita celastri | PAD S00034: fungarium Saccardo, isolectotype | MW626901 | MW626909 | - | - | - |
| Xylaria bambusicola | Xylaria bambusicola | WPS 205, holotype [48,50] | - | - | EF026123 | - | AY951762 |
| Xylaria hypoxylon | Xylaria hypoxylon | CBS 122620 [6,59] | - | - | KY204024 | KY610495 | KX271279 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forin, N.; Vizzini, A.; Fainelli, F.; Ercole, E.; Baldan, B. Taxonomic Re-Examination of Nine Rosellinia Types (Ascomycota, Xylariales) Stored in the Saccardo Mycological Collection. Microorganisms 2021, 9, 666. https://doi.org/10.3390/microorganisms9030666
Forin N, Vizzini A, Fainelli F, Ercole E, Baldan B. Taxonomic Re-Examination of Nine Rosellinia Types (Ascomycota, Xylariales) Stored in the Saccardo Mycological Collection. Microorganisms. 2021; 9(3):666. https://doi.org/10.3390/microorganisms9030666
Chicago/Turabian StyleForin, Niccolò, Alfredo Vizzini, Federico Fainelli, Enrico Ercole, and Barbara Baldan. 2021. "Taxonomic Re-Examination of Nine Rosellinia Types (Ascomycota, Xylariales) Stored in the Saccardo Mycological Collection" Microorganisms 9, no. 3: 666. https://doi.org/10.3390/microorganisms9030666
APA StyleForin, N., Vizzini, A., Fainelli, F., Ercole, E., & Baldan, B. (2021). Taxonomic Re-Examination of Nine Rosellinia Types (Ascomycota, Xylariales) Stored in the Saccardo Mycological Collection. Microorganisms, 9(3), 666. https://doi.org/10.3390/microorganisms9030666