
Rehydration before Application Improves Functional Properties of Lyophilized Lactiplantibacillus plantarum HAC03

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Fermentation and Freeze-Drying Process
2.3. Food Additives and Rehydration Conditions
2.4. Simulated Gastro-Intestinal Passage Resistance Assay
2.5. Bacterial Adhesion to Mucin
2.6. Bacterial Zeta Potential
2.7. Product Formulation
2.8. Determination of Bacterial Viability by Flow Cytometry
2.9. Adhesion to Intestinal Epithelial Cell Line
2.10. Immune Stimulation In Vitro
2.11. Bacterial Hydrophobicity
2.12. Statistical Analysis
3. Results
3.1. Influence of Single Food Additives on Lp. plantarum HAC03 Viability after GIT Stress, Mucin Adhesion and Zeta Potential
3.2. Effect of the Formulation Mixtures on Lp. plantarum HAC03 Viability after SSDP
3.3. Determination of Lp. plantarum HAC03 Viability by Flow Cytometry and Influence of the B-Active Formulation
3.4. Impact of B-Active Formulation on Lp. plantarum HAC03 Zeta Potential and Hydrophobicity
3.5. Influence of B-Active Formulation on the Adhesion of Lp. plantarum HAC03 to Intestinal Epithelial Cells and Mucin
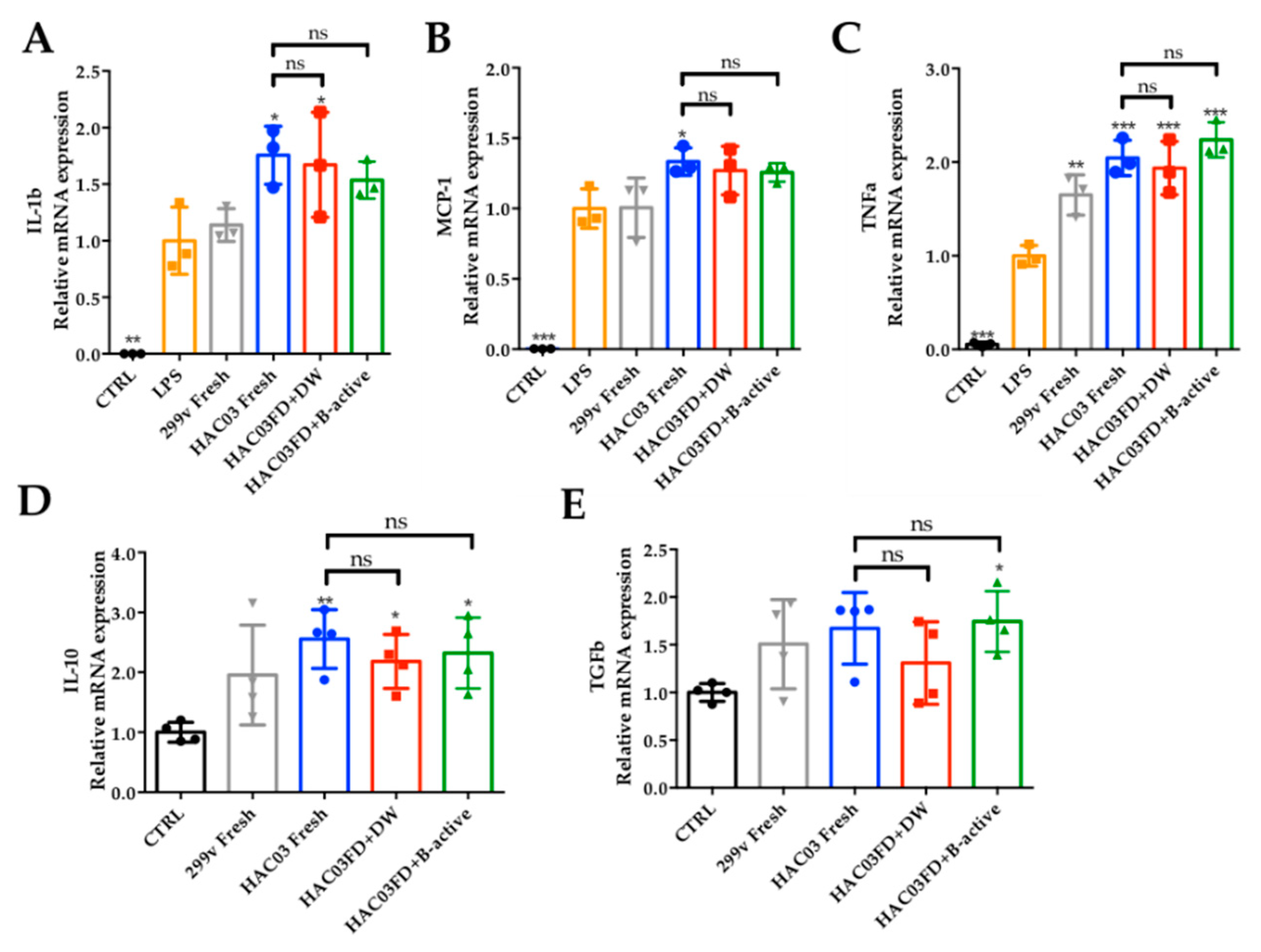
3.6. Effect of B-Active Formulation on the Immunomodulatory Potential of Lp. plantarum HAC03
3.7. Influence of B-Active Formulation on the Viability of Different Potential Probiotic Strains
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Van Bokhorst-van de Veen, H.; Lee, I.; Marco, M.; Wels, M.; Bron, P.A.; Kleerebezem, M. Modulation of Lactobacillus plantarum Gastrointestinal Robustness by Fermentation Conditions Enables Identification of Bacterial Robustness Markers. PLoS ONE 2012, 7, e39053. [Google Scholar] [CrossRef] [Green Version]
- Senz, M.; Keil, C.; Schmacht, M.; Palinski, S.; Cämmerer, B.; Hageböck, M. Influence of Media Heat Sterilization Process on Growth Performance of Representative Strains of the Genus Lactobacillus. Fermentation 2019, 5, 20. [Google Scholar] [CrossRef] [Green Version]
- Lodato, P.; Huergo, M.S.; Buera, M.P. Viability and thermal stability of a strain of Saccharomyces cerevisiae freeze-dried in different sugar and polymer matrices. J. Microbiol. Biotechnol. 1999, 52, 215–220. [Google Scholar] [CrossRef]
- Zhao, G.; Zhang, G. Effect of protective agents, freezing temperature, rehydration media on viability of malolactic bacteria subjected to freeze-drying. J. Appl. Microbiol. 2005, 99, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Morgan, C.A.; Herman, N.; White, P.A.; Vesey, G. Preservation of micro-organisms by drying; a review. J. Microbiol. Meth. 2006, 66, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Schoug, A.; Olsson, J.; Carlfors, J.; Schnürer, J.; Hakansson, S. Freeze-drying of Lactobacillus coryniformis Si3—effects of sucrose concentration, cell density and freezing rate on cell survival and thermophysical properties. Cryobiology 2006, 53, 119–127. [Google Scholar] [CrossRef]
- Yeo, S.; Shin, H.S.; Lee, H.W.; Hong, D.; Park, H.; Holzapfel, W.; Huh, C.S. Determination of optimized growth medium and cryoprotective additives to enhance the growth and survival of Lactobacillus salivarius. J. Microbiol. Biotechnol. 2018, 28, 718–731. [Google Scholar] [CrossRef] [PubMed]
- Edward, V.A.; Huch, M.; Dortu, M.; Thonart, P.; Egounlety, M.; Van Zyl, P.J. Biomass production and small-scale testing of freeze-dried lactic acid bacteria starter strains for cassava fermentations. Food Control. 2011, 22, 389–395. [Google Scholar] [CrossRef]
- Panoff, J.M.; Thammavongs, B.; Guéguen, M.; Boutibonnes, P. Cold stress responses in mesophilic bacteria. Cryobiology 1998, 36, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Galdeano, C.M.; Perdigón, G. Role of viability of probiotic strains in their persistence in the gut and in mucosal immune stimulation. J. Appl. Microbiol. 2004, 97, 673–681. [Google Scholar] [CrossRef]
- Pelletier, X.; Laure-Boussuge, S.; Donazzolo, Y. Hydrogen excretion upon ingestion of dairy products in lactose-intolerant male subjects: Importance of the live flora. Eur. J. Clin. Nutr. 2001, 55, 509–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iaconelli, C.; Lemetais, G.; Kechaou, N.; Chain, F.; Bermúdez-Humarán, L.G.; Langella, P.; Gervais, P.; Beney, L. Drying process strongly affects probiotics viability and functionalities. J. Biotechnol. 2015, 214, 17–26. [Google Scholar] [CrossRef]
- Kiekens, S.; Vandenheuvel, D.; Broeckx, G.; Claes, I.; Allonsius, C.; Boeck, I.D.; Thys, S.; Timmermans, J.P.; Kiekens, F.; Lebeer, S. Impact of spray-drying on the pili of Lactobacillus rhamnosus GG. Microb. Biotechnol. 2019, 12, 849–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, V.; Burkhardt, D.H.; Rouskin, S.; Li, G.-W.; Weissman, J.S.; Gross, C.A. A Stress Response that Monitors and Regulates mRNA Structure Is Central to Cold Shock Adaptation. Mol. Cell 2018, 70, 274–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosnea, L.A.; Kourkoutas, Y.; Albantaki, N.; Tzia, C.; Koutinas, A.A.; Kanellaki, M. Functionality of freeze-dried L. casei cells immobilized on wheat grains. LWT Food Sci. Technol. 2009, 42, 1696–1702. [Google Scholar] [CrossRef]
- Dimitrellou, D.; Kandylis, P.; Kourkoutas, Y. Effect of cooling rate, freeze-drying, and storage on survival of free and immobilized Lactobacillus casei ATCC 393. LWT Food Sci. Technol. 2016, 69, 468–473. [Google Scholar] [CrossRef]
- Chen, H.; Lin, C.; Chen, M. The Effects of Freeze Drying and Rehydration on Survival of Microorganisms in Kefir. Asian-Austral. J. Anim. Sci. 2005, 19, 126–130. [Google Scholar]
- Leach, R.H.; Scott, W.J. The Influence of Rehydration on the Viability of Dried Micro-Organisms. J. Gen. Microbiol. 1959, 21, 295–307. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, J.H.; Enright, J.J. Effect of low moisture levels on the thermostability of active dry yeast. Food Tech. 1957, 11, 859. [Google Scholar]
- Nagashima, A.I.; Pansiera, P.E.; Baracat, M.M.; Gómez, R.J. Development of effervescent products, in powder and tablet form, supplemented with probiotics Lactobacillus acidophilus and Saccharomyces boulardii. Food Sci. Technol. 2013, 33, 605–611. [Google Scholar] [CrossRef] [Green Version]
- Font de Valdez, G.; de Girori, G.S.; de Ruiz Holgado, A.P.; Oliver, G. Rehydration conditions and viability of freeze-dried lactic acid bacteria. Cryobiology 1985, 22, 574–577. [Google Scholar] [CrossRef]
- Schillinger, U.; Guigas, C.; Holzapfel, W.H. In vitro adherence and other functional properties of lactobacilli used in probiotic yoghurt-like products. Int. Dairy J. 2005, 15, 1289–1297. [Google Scholar] [CrossRef]
- FAO/WHO. Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria. Report of a Joint FAO/WHO Expert Consultation on Evaluation of Health and Nutritional Properties of Probiotics in Food including Powder Milk with Live Lactic Acid Bacteria. Available online: http://www.fao.org/3/a-a0512e.pdf (accessed on 6 January 2020).
- Haberer, P.; Du Toit, M.; Dicks, L.M.T.; Ahrens, F.; Holzapfel, W.H. Effect of potentially probiotic lactobacilli on faecal enzyme activity in minipigs on a high-fat, high-cholesterol diet—A preliminary in vivo trial. Int. J. Food Microbiol. 2002, 87, 287–291. [Google Scholar] [CrossRef]
- Bengoa, A.A.; Zavala, L.; Carasi, P.; Trejo, S.A.; Bronsoms, S.; de los Ángeles Serradell, M.; Abraham, A.G. Simulated gastrointestinal conditions increase adhesion ability of Lactobacillus paracasei strains isolated from kefir to Caco-2 cells and mucin. Food Res. Int. 2018, 103, 462–467. [Google Scholar] [CrossRef]
- BD Biosciences. Bacterial Detection and Live/Dead Discrimination by Flow Cytometry. Available online: https://www.bdbiosciences.com/documents/Bacterial_Detection_Live_Dead.pdf (accessed on 20 January 2020).
- Bertuzzi, A.; D’Agnano, I.; Gandolfi, A.; Graziano, A.; Starace, G.; Ubezio, P. Study of propidium iodide binding to DNA in intact cells by flow cytometry. Cell Biophys. 1990, 17, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Sophatha, B.; Piwat, S.; Teanpaisan, R. Adhesion, anti-adhesion and aggregation properties relating to surface charges of selected Lactobacillus strains: Study in Caco-2 and H357 cells. Arch. Microbiol. 2020, 202, 1349–1357. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gonzalez, N.; Prete, R.; Battista, N.; Corsetti, A. Adhesion Properties of Food-Associated Lactobacillus plantarum Strains on Human Intestinal Epithelial Cells and Modulation of IL-8 Release. Front. Microbiol. 2018, 9, 2392. [Google Scholar] [CrossRef]
- Buntin, N.; de Vos, W.M.; Hongpattarakere, T. Variation of mucin adhesion, cell surface characteristics, and molecular mechanisms among Lactobacillus plantarum isolated from different habitats. Appl. Microbiol. Biotechnol. 2017, 101, 7663–7674. [Google Scholar] [CrossRef]
- Pelletier, C.; Bouley, C.; Cayuela, C.; Bouttier, S.; Bourlioux, P.; Bellon-Fontaine, M.N. Cell surface characteristics of Lactobacillus casei subsp. casei, Lactobacillus paracasei subsp. paracasei, and Lactobacillus rhamnosus strains. Appl. Environ. Microbiol. 1997, 63, 1725–1731. [Google Scholar] [CrossRef] [Green Version]
- Neuhaus, F.C.; Baddiley, J. A Continuum of Anionic Charge: Structures and Functions of d-Alanyl-Teichoic Acids inGram-Positive Bacteria. Microbiol. Mol. Biol. Rev. 2003, 67, 686–723. [Google Scholar] [CrossRef] [Green Version]
- Cowan, M.M.; Van der Mei, H.C.; Stokroos, I.; Busscher, H.J. Heterogeneity of Surfaces of Subgingival Bacteria as Detected by Zeta Potential Measurements. J. Dent. Res. 1992, 71, 1803–1806. [Google Scholar] [CrossRef]
- De Wouters, T.; Jans, C.; Niederberger, T.; Fischer, P.; Rühs, P.A. Adhesion Potential of Intestinal Microbes Predicted by Physico-Chemical Characterization Methods. PLoS ONE 2015, 10, e0136437. [Google Scholar]
- Park, S.; Ji, Y.; Park, H.; Lee, K.; Park, H.; Beck, B.R.; Shin, H.; Holzapfel, W.H. Evaluation of functional properties of lactobacilli isolated from Korean white kimchi. Food Control 2016, 69, 5–12. [Google Scholar] [CrossRef]
- Molin, G. Probiotic in foods not containing milk or milk constituents, with special reference to Lactobacillus plantarum 299v. Am. J. Clin. Nutr. 2001, 73, 380–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FDA. Substances Added to Food. 2020. Available online: https://www.accessdata.fda.gov/scripts/cder/daf/index.cfm (accessed on 21 April 2020).
- Ji, Y.; Kim, H.; Park, H.; Lee, J.; Lee, H.; Shin, H.; Holzapfel, W.H. Functionality and safety of lactic bacterial strains from Korean kimchi. Food Control 2013, 31, 467–473. [Google Scholar] [CrossRef]
- Laparra, J.; Sanz, Y. Comparison of in vitro models to study bacterial adhesion to the intestinal epithelium. Lett. Appl. Microbiol. 2009, 49, 695–701. [Google Scholar] [CrossRef] [Green Version]
- R&D Systems. Flow Cytometry Protocol for Analysis of Cell Viability using Propidium Iodide. 2016. Available online: https://www.rndsystems.com/resources/protocols/flow-cytometry-protocol-analysis-cell-viability-using-propidium-iodide (accessed on 15 January 2020).
- Botes, M.; Loos, B.; Van Reenen, C.A.; Dicks, L.M. Adhesion of the probiotic strains Enterococcus mundtii ST4SA and Lactobacillus plantarum 423 to Caco-2 cells under conditions simulating the intestinal tract, and in the presence of antibiotics and anti-inflammatory medicaments. Arch. Microbiol. 2008, 190, 573–584. [Google Scholar] [CrossRef]
- Khalil, A.; Omran, H.; Alsheikh, F. Balance of pro- and anti-inflammatory cytokines in livers of high fat diet rats exposed to fractionated gamma irradiation. BMC Res. Notes 2018, 11, 741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hul, M.; Karnik, K.; Canene-Adams, K.; de Souza, M.; van den Abbeele, P.; Mrzarti, M.; Delzenne, N.M.; Everard, A.; Cani, P.D. Comparison of the effects of soluble corn fiber and fructooligosaccharides on metabolism, inflammation, and gut microbiome of high-fat diet-fed mice. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E779–E791. [Google Scholar] [CrossRef]
- Zhu, J.; Yang, F.; Sang, L.; Zhai, J.; Zhang, X.; Yue, D.; Sun, X. IL-33 Aggravates DSS-Induced Acute Colitis in Mouse Colon Lamina Propria by Enhancing Th2 Cell Responses. Mediators Inflamm. 2015, 913041. [Google Scholar] [CrossRef] [Green Version]
- Xie, S.; Macedo, P.; Hew, M.; Nassenstein, C.; Lee, K.Y.; Chung, K.F. Expression of transforming growth factor-beta (TGF-beta) in chronic idiopathic cough. Respir. Res. 2009, 10, 40. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Marneros, A.G. Macrophages are essential for the early wound healing response and the formation of a fibrovascular scar. Am. J. Pathol. 2013, 182, 2407–2417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arellano-Ayala, K.; Ascencio-Valle, F.J.; Gutiérrez-González, P.; Estrada-Girón, Y.; Torres-Vitela, M.R.; Macías-Rodríguez, M.E. Hydrophobic and adhesive patterns of lactic acid bacteria and their antagonism against foodborne pathogens on tomato surface (Solanum lycopersicum L.). J. Appl. Microbiol. 2020, 129, 876–891. [Google Scholar] [CrossRef] [PubMed]
- Dommels, Y.E.; Kemperman, R.A.; Zebregs, Y.E.; Draaisma, R.B.; Jol, A.; Wolvers, D.A.; Albers, R. Survival of Lactobacillus reuteri DSM 17938 and Lactobacillus rhamnosus GG in the Human Gastrointestinal Tract with Daily Consumption of a Low-Fat Probiotic Spread. J. Appl. Environ. Microbiol. 2009, 75, 6198–6204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theunissen, J.J.H.; Stolz, E.; Michel, M.F. The effects of medium and rate of freezing on the survival of chlamydias after lyophilization. J. Appl. Bacteriol. 1993, 75, 473–477. [Google Scholar] [CrossRef]
- Corcoran, B.M.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Survival of probiotic lactobacilli in acidic environments is enhanced in the presence of metabolizable sugars. Appl. Environ. Microbiol. 2005, 71, 3060–3067. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.S.; Silva, J.; Ho, P.; Teixeira, P.; Malcata, F.X.; Gibbs, P. Relevant factors for the preparation of freeze-dried lactic acid bacteria. Int. Dairy J. 2004, 14, 835–847. [Google Scholar] [CrossRef]
- Watson, D.; Sleator, R.D.; Hill, C.; Gahan, C.G. Enhancing bile tolerance improves survival and persistence of Bifidobacterium and Lactococcus in the murine gastrointestinal tract. BMC Microbiol. 2008, 8, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du Toit, E.; Vesterlund, S.; Gueimonde, M.; Salminen, S. Assessment of the effect of stress-tolerance acquisition on some basic characteristics of specificprobiotics. Int. J. Food Microbiol. 2013, 165, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [Green Version]
- Soni, K.A.; Balasubramanian, A.K.; Beskok, A.; Pillai, S.D. Zeta Potential of Selected Bacteria in Drinking Water When Dead, Starved, or Exposed to Minimal and Rich Culture Media. Curr. Microbiol. 2007, 56, 93–97. [Google Scholar] [CrossRef]
- Ng, W.; Ting, Y. Zeta potential of bacterial cells: Effect of wash buffers. Peer J. Prepr. 2017, 5, e110v7. [Google Scholar]
- Costa, E.; Usall, J.; Teixido, N.; Garcia, N.; Vinas, I. Effect of protective agents, rehydration media and initial cell concentration on viability of Pantoea agglomerans strain CPA-2 subjected to freeze-drying. J. Appl. Microbiol. 2000, 89, 793–800. [Google Scholar] [CrossRef]
- Sinha, R.N.; Shukla, A.K.; Lal, M.; Ranganathan, B. Rehydration of Freeze-Dried Cultures of Lactic Streptococci. J. Food Sci. 1982, 47, 668–669. [Google Scholar] [CrossRef]
- Fu, N.; Chen, X.D. Towards a maximal cell survival in convective thermal drying processes. Food Res. Int. 2011, 44, 1127–1149. [Google Scholar] [CrossRef]
- Golowczyc, M.A.; Silva, J.; Teixeira, P.; De Antoni, G.L.; Abraham, A.G. Cellular injuries of spray-dried Lactobacillus spp. isolated from kefir and their impact on probiotic properties. Int. J. Food Microbiol. 2011, 144, 556–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guergoletto, K.B.; Tsuruda, A.Y.; Hirooka, E.Y.; Martins, E.P.; Souza, J.C.; Sivieri, K.; Garcia, S. Dried Probiotics for Use in Functional Food Applications; INTECH Open Access Publisher: London, UK, 2012. [Google Scholar]
- Wisselink, H.W.; Weusthuis, R.A.; Eggink, G.; Hugenholtz, J.; Grobben, G.J. Mannitol production by lactic acid bacteria: A review. Int. Dairy J. 2002, 12, 151–161. [Google Scholar] [CrossRef]
- Carvalho, A.S.; Silva, J.; Ho, P.; Teixeira, P.; Malcata, F.X.; Gibbs, P. Protective effect of sorbitol and monosodium glutamate during storage of freeze- dried lactic acid bacteria. Le Lait INRA Ed. 2003, 83, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Iraporda, C.; Rubel, I.A.; Manrique, G.D.; Abraham, A.G. Influence of inulin rich carbohydrates from Jerusalem artichoke (Helianthus tuberosus L.) tubers on probiotic properties of Lactobacillus strains. LWT Food Sci. Technol. 2019, 101, 738–746. [Google Scholar] [CrossRef]
- Ehrmann, M.A.; Kurzak, P.; Bauer, J.; Vogel, R.F. Characterization of lactobacilli towards their use as probiotic adjuncts in poultry. J. Appl. Microbiol. 2002, 92, 966–975. [Google Scholar] [CrossRef]
- Wang, X.; Liu, L.; Zhou, X.; Huo, Y.; Gao, J.; Gu, H. Casein phosphopeptide combined with fluoride enhances the inhibitory effect on initial adhesion of Streptococcus mutans to the saliva-coated hydroxyapatite disc. BMC Oral Health 2020, 20, 169. [Google Scholar] [CrossRef] [PubMed]
- Santivarangkna, C.; Higl, B.; Foerst, P. Protection mechanisms of sugars during different stages of preparation process of dried lactic acid starter cultures. Food Microbiol. 2008, 25, 429–441. [Google Scholar] [CrossRef] [PubMed]
- García-Cayuela, T.; Korany, A.M.; Bustos, I.P.; Gómez de Cadiñanos, L.; Requena, T.; Peláez, C.; Martínez-Cuesta, M.C. Adhesion abilities of dairy Lactobacillus plantarum strains showing an aggregation phenotype. Food Res. Int. 2014, 57, 44–50. [Google Scholar] [CrossRef]
- Lebeer, S.; Claes, I.; Tytgat, H.L.; Verhoeven, T.L.; Marien, E.; von Ossowski, I.; Reunanen, J.; Palva, A.; Vos, W.M.; Keersmaecker, S.C.; et al. Functional analysis of Lactobacillus rhamnosus GG pili in relation to adhesion and immunomodulatory interactions with intestinal epithelial cells. Appl. Environ. Microbiol. 2012, 78, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Golowczyc, M.A.; Mobili, P.; Abraham, A.G.; Garrote, G.L.; De Antoni, G.L. Protective action of Lactobacillus kefir carrying S-layer against Salmonella enterica serovar Enteritidis. Int. J. Food Microbiol. 2007, 118, 264–273. [Google Scholar] [CrossRef]
- Gheziel, C.; Russo, P.; Arena, M.P. Evaluating the Probiotic Potential of Lactobacillus plantarum Strains from Algerian Infant Feces: Towards the Design of Probiotic Starter Cultures Tailored for Developing Countries. Probiotics Antimicro. 2019, 11, 113–123. [Google Scholar] [CrossRef]
- Konstantinov, S.R.; Smidt, H.; de Vos, W.M.; Bruijns, S.C.M.; Singh, S.K.; Valence, F.; Molle, D.; Lortal, S.; Altermann, E.; Klaenhammer, T.R.; et al. S layer protein a of Lactobacillus acidophilus NCFM regulates immature dendritic cell and T-cell functions. Proc. Natl. Acad. Sci. USA 2008, 105, 19474–19479. [Google Scholar] [CrossRef] [Green Version]
- Vargas Garcia, C. The Role of Pili on Lactobacillus rhamnosus GG in the Interaction with Macrophages and Other Immune Cells. Ph.D. Thesis, KU Leuven, Flanders, Belgium, 2015. [Google Scholar]
- Båth, K. Factors Important for Persistence of Lactobacillus reuteri in the Gastrointestinal Tract: A Study of Extracellular Proteins, Stress Response and Survival of Mutants in a Model System. Doctoral Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2007. [Google Scholar]
- West, A.H.; Stock, A.M. Histidine kinases and response regulator proteins in two-component signaling systems. Trends Biochem. Sci. 2001, 26, 369–376. [Google Scholar] [CrossRef]
- Yeo, S.; Park, H.; Ji, Y.; Park, S.; Yang, J.; Lee, J.; Holzapfel, W. Influence of gastrointestinal stress on autoinducer-2 activity of two Lactobacillus species. FEMS Microbiol. Ecol. 2015, 91, fiv065. [Google Scholar] [CrossRef] [Green Version]
- Park, H.; Lee, K.; Yeo, S.; Shin, H.; Holzapfel, W.H. Autoinducer-2 Quorum Sensing Influences Viability of Escherichia coli O157:H7 under Osmotic and In Vitro Gastrointestinal Stress Conditions. Front. Microbiol. 2017, 8, 1077. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Log CFU/mL | Log CFU/mL | Survival (%) | Log CFU/mL | Survival (%) | ||
|---|---|---|---|---|---|---|
| Fresh cells Lp. plantarum HAC03 | 8.47 ± 0.01 | 8.47 ± 0.07 | 99.83 | 7.90 ± 0.12 | 27.24 *** | |
| Freeze-dried cells HAC03+DW | 8.48 ± 0.04 | 7.57 ± 0.09 | 12.24 | 7.17 ± 0.11 | 4.92 | |
| A | Sucrose | 8.46 ± 0.12 | 8.46 ± 0.18 | 104.77 | 7.94 ± 0.18 | 31.06 *** |
| Malic acid | ||||||
| Sorbitol | ||||||
| Soy peptone | ||||||
| Riboflavin | ||||||
| B | Sucrose | 8.51 ± 0.07 | 8.44 ± 0.06 | 84.77 | 8.07 ± 0.05 | 36.35 *** |
| Malic acid | ||||||
| Sorbitol | ||||||
| Soy peptone | ||||||
| C | Sucrose | 8.42 ± 0.03 | 8.28 ± 0.06 | 71.50 | 7.91 ± 0.01 | 31.06 *** |
| Sorbitol | ||||||
| Soy peptone | ||||||
| D | Soy peptone | 8.66 ± 0.05 | 8.27 ± 0.02 | 40.82 | 6.10 ± 0.04 | 0.28 |
| Tryptophan | ||||||
| Sorbitol | ||||||
| Sodium bicarbonate | ||||||
| E | Soy peptone | 8.51 ± 0.23 | 8.57 ± 0.06 | 120.89 | 7.17 ± 0.01 | 4.93 |
| Tryptophan | ||||||
| Glucose | ||||||
| Sodium bicarbonate | ||||||
| Treatment | Initial Counts | After 1 h Acid Stress | After 2 h Bile Stress | Mucin Adhesion (%) | Zeta Potential (mV) | |||
|---|---|---|---|---|---|---|---|---|
| Log CFU/mL | Log CFU/mL | Survival (%) | Log CFU/mL | Survival (%) | ||||
| Fresh cells of Lp. plantarum HACO3 | 8.38 ± 0.13 | 8.07 ± 0.05 | 50.30 | 7.77 ± 0.13 | 24.51 *** | 4.38 ± 0.38 *** | −17.97 ± 1.12 *** | |
| Freeze-dried cells of HAC03 + DW | 7.99 ± 0.12 | 6.71 ± 0.17 | 5.40 | 6.27 ± 0.28 | 2.10 | 1.55 ± 0.36 | 21.03 ± 1.46 | |
| 1 | Arabinose | 8.09 ± 0.04 | 6.55 ± 0.02 | 2.91 | 5.90 ± 0.13 | 0.66 ** | 0.84 ± 0.00 * | 24.73 ± 1.20 * |
| 2 | Xylose | 7.95 ± 0.03 | 6.74 ± 0.32 | 7.12 | 6.20 ± 0.37 | 2.15 | 1.57 ± 0.05 | 23.13 ± 1.24 |
| 3 | Rhamnose | 8.00 ± 0.01 | 6.62 ± 0.09 | 4.28 | 6.14 ± 0.12 | 1.44 | 0.76 ± 0.03 * | 23.13 ± 2.11 |
| 4 | Mannose | 8.02 ± 0.01 | 7.80 ± 0.02 | 60.21 | 7.14 ± 0.07 | 13.48 *** | 18.80 ± 0.68 *** | 24.90 ± 1.64 * |
| 5 | Fructose | 8.07 ± 0.12 | 7.66 ± 0.03 | 39.24 | 7.20 ± 0.25 | 13.76 *** | 5.79 ± 0.21 *** | 24.77 ± 2.33 |
| 6 | Mannitol | 8.02 ± 0.01 | 6.91 ± 0.18 | 8.02 | 6.47 ± 0.07 | 2.80 | 0.99 ± 0.02 | 22.70 ± 1.11 |
| 7 | Sucrose | 7.95 ± 0.03 | 7.63 ± 0.06 | 48.25 | 7.07 ± 0.28 | 14.52 *** | 1.01 ± 0.07 | 21.60 ± 1.28 |
| 8 | Sorbitol | 8.00 ± 0.05 | 7.54 ± 0.09 | 34.75 | 7.22 ± 0.03 | 16.69 *** | 1.45 ± 0.01 | 23.17 ± 1.12 |
| 9 | Glucose | 7.93 ± 0.04 | 7.60 ± 0.10 | 48.07 | 7.30 ± 0.02 | 23.50 *** | 6.50 ± 0.37 *** | 23.10 ± 1.67 |
| 10 | Maltose | 8.19 ± 0.16 | 7.72 ± 0.04 | 35.75 | 7.15 ± 0.16 | 9.39 *** | 9.74 ± 0.14 *** | 21.03 ± 1.16 |
| 11 | Trehalose | 7.94 ± 0.04 | 6.52 ± 0.24 | 3.95 | 6.13 ± 0.11 | 1.54 | 0.69 ± 0.04 * | 22.87 ± 1.40 |
| 12 | Alginic acid | 8.00 ± 0.00 | 4.00 ± 0.00 | 0.01 | 3.49 ± 0.00 | 0.003 ** | nd | −4.86 ± 1.24 *** |
| 13 | Starch | 8.10 ± 0.10 | 6.47 ± 0.13 | 2.51 | 6.16 ± 0.04 | 1.18 | 0.26 ± 0.02 ** | 2.27 ± 0.89 *** |
| 14 | Gelatin | 8.10 ± 0.01 | 6.89 ± 0.13 | 6.22 | 6.68 ± 0.37 | 4.48 | 2.01 ± 0.94 | 23.40 ± 3.46 |
| 15 | Albumin | 8.11 ± 0.18 | 6.93 ± 0.05 | 7.11 | 6.85 ± 0.17 | 6.46 * | 0.83 ± 0.07 * | 28.73 ± 2.20 ** |
| 16 | Pepsin | 8.04 ± 0.04 | 7.16 ± 0.09 | 13.29 | 6.89 ± 0.14 | 7.30 ** | 2.45 ± 0.14 * | −11.57 ± 1.14 *** |
| 17 | Soy Peptone | 8.23 ± 0.04 | 7.53 ± 0.38 | 24.86 | 7.31 ± 0.33 | 14.43 ** | 1.42 ± 0.16 | −2.66 ± 0.43 *** |
| 18 | Yeast extract | 8.11 ± 0.06 | 7.36 ± 0.72 | 29.65 | 7.11 ± 0.75 | 17.27 * | 3.23 ± 0.10 *** | −8.58 ± 0.39 *** |
| 19 | Arginine | 7.44 ± 0.79 | 4.00 ± 0.00 | 0.07 | 3.61 ± 0.17 | 0.04 ** | nd | 0.74 ± 0.10 *** |
| 20 | Tryptophan | 8.12 ± 0.02 | 6.42 ± 0.08 | 1.99 | 6.14 ± 0.04 | 1.05 | 1.69 ± 0.19 | 26.63 ± 0.25 ** |
| 21 | Phenyl alanine | 8.00 ± 0.02 | 6.53 ± 0.05 | 3.41 | 6.15 ± 0.13 | 1.44 | 0.14 ± 0.001 *** | 3.44 ± 6.10 ** |
| 22 | Ornithine | 8.10 ± 0.02 | 6.47 ± 0.38 | 2.78 | 6.01 ± 0.16 | 0.84 * | 1.44 ± 0.15 | 0.09 ± 0.15 *** |
| 23 | Glutamic acid | 8.03 ± 0.03 | 6.87 ± 0.91 | 26.93 | 4.19 ± 1.09 | 0.10 ** | nd | 0.26 ± 0.20 *** |
| 24 | Proline | 8.14 ± 0.07 | 6.91 ± 0.04 | 5.91 | 6.45 ± 0.22 | 2.24 | 1.32 ± 0.40 | 14.60 ± 1.21 ** |
| 25 | Lysine | 8.01 ± 0.24 | 6.05 ± 0.37 | 1.16 | 5.72 ± 0.41 | 0.54 | 0.97 ± 0.09 | 0.00 ± 0.16 *** |
| 26 | Serine | 7.99 ± 0.20 | 6.58 ± 0.18 | 3.92 | 6.26 ± 0.13 | 1.87 | 1.85 ± 0.11 | 8.08 ± 0.96 *** |
| 27 | Threonine | 7.87 ± 0.03 | 6.52 ± 0.13 | 4.62 | 6.15 ± 0.10 | 1.97 | 0.91 ± 0.10 * | 10.68 ± 2.14 ** |
| 28 | Aspartic Acid | 7.84 ± 0.05 | 7.06 ± 0.69 | 29.95 | 5.80 ± 0.78 | 1.86 | nd | 17.10 ± 0.82 * |
| 29 | Tyrosine | 7.68 ± 0.30 | 6.08 ± 0.57 | 3.12 | 5.84 ± 0.61 | 1.82 | 6.89 ± 0.16 *** | 4.70 ± 1.35 *** |
| 30 | Histidine | 7.70 ± 0.12 | 6.27 ± 0.01 | 3.86 | 5.87 ± 0.05 | 1.57 | 2.86 ± 0.60 ** | −0.20 ± 0.67 *** |
| 31 | Sodium phosphate | 7.87 ± 0.00 | 6.29 ± 0.04 | 2.66 | 6.01 ± 0.09 | 1.39 | 0.96 ± 0.04 | −5.53 ± 0.34 *** |
| 32 | Sodium L-tartrate | 8.03 ± 0.19 | 4.00 ± 0.00 | 0.01 | 3.49 ± 0.00 | 0.003 * | 0.72 ± 0.05 * | −4.26 ± 1.19 *** |
| 33 | Sodium bicarbonate | 8.08 ± 0.02 | 6.56 ± 0.02 | 3.06 | 6.04 ± 0.15 | 0.96 | 0.83 ± 0.04 * | 2.69 ± 0.12 *** |
| 34 | Malic acid | 8.00 ± 0.00 | 4.00 ± 0.00 | 0.01 | 3.49 ± 0.00 | 0.003 ** | nd | 21.57 ± 1.89 |
| 35 | Pyruvic acid | 8.00 ± 0.00 | 4.00 ± 0.00 | 0.01 | 3.49 ± 0.00 | 0.003 ** | nd | 2.54 ± 0.04 *** |
| 36 | Betaine | 7.90 ± 0.04 | 6.57 ± 0.05 | 4.75 | 6.19 ± 0.06 | 1.95 | 0.67 ± 0.03 * | 10.05 ± 1.34 *** |
| 37 | Taurine | 7.87 ± 0.19 | 6.50 ± 0.17 | 4.35 | 6.25 ± 0.13 | 2.46 | 0.48 ± 0.01 ** | 14.73 ± 0.46 ** |
| 38 | Riboflavin | 7.96 ± 0.22 | 6.84 ± 0.16 | 7.70 | 6.22 ± 0.20 | 1.85 | 0.81 ± 0.14 * | −0.20 ± 1.43 *** |
| 39 | Thiamine | 7.77 ± 0.32 | 6.41 ± 0.34 | 4.46 | 5.47 ± 0.33 | 0.50 | 0.63 ± 0.06 ** | 3.47 ± 0.40 *** |
| 40 | Ascorbic acid | 6.49 ± 0.03 | 4.08 ± 0.11 | 0.40 | 3.49 ± 0.00 | 0.10 ** | nd | 21.07 ± 1.99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arellano-Ayala, K.; Lim, J.; Yeo, S.; Bucheli, J.E.V.; Todorov, S.D.; Ji, Y.; Holzapfel, W.H. Rehydration before Application Improves Functional Properties of Lyophilized Lactiplantibacillus plantarum HAC03. Microorganisms 2021, 9, 1013. https://doi.org/10.3390/microorganisms9051013
Arellano-Ayala K, Lim J, Yeo S, Bucheli JEV, Todorov SD, Ji Y, Holzapfel WH. Rehydration before Application Improves Functional Properties of Lyophilized Lactiplantibacillus plantarum HAC03. Microorganisms. 2021; 9(5):1013. https://doi.org/10.3390/microorganisms9051013
Chicago/Turabian StyleArellano-Ayala, Karina, Juhwan Lim, Subin Yeo, Jorge Enrique Vazquez Bucheli, Svetoslav Dimitrov Todorov, Yosep Ji, and Wilhelm Heinrich Holzapfel. 2021. "Rehydration before Application Improves Functional Properties of Lyophilized Lactiplantibacillus plantarum HAC03" Microorganisms 9, no. 5: 1013. https://doi.org/10.3390/microorganisms9051013
APA StyleArellano-Ayala, K., Lim, J., Yeo, S., Bucheli, J. E. V., Todorov, S. D., Ji, Y., & Holzapfel, W. H. (2021). Rehydration before Application Improves Functional Properties of Lyophilized Lactiplantibacillus plantarum HAC03. Microorganisms, 9(5), 1013. https://doi.org/10.3390/microorganisms9051013