
Characterization of Partially Purified Bacteriocins Produced by Enterococcus faecium Strains Isolated from Soybean Paste Active Against Listeria spp. and Vancomycin-Resistant Enterococci

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Putative LAB Strains and Screening for Bacteriocin Production
2.2. Differentiation and Identification of Bacterial Isolates
2.3. Evaluation of Nature, Integrity and Stability of the Bioactive Molecule
2.4. Bacteriocin Production, Growth Kinetics and Fermentation
2.5. Effect of Varying Incubation Temperature on Bacteriocin Production of Studied E. faecium Strains
2.6. Screening for Associated Bacteriocin Genes
2.7. Bacterial Growth Inhibition (Cell Lysis)
2.8. Assessment of the Range of Activity of Bacteriocins Produced by E. faecium Strains
2.9. Quantification of Adhered Bacteriocin to the Producer Cells
2.10. Adsorption of Bacteriocin to the Test Organisms
2.11. Bacteriocin Partial Purification
2.12. Molecular Mass Estimation of Bacteriocins Using Tricine-SDS-PAGE
2.13. Evaluation of Bacterial Growth Inhibition of Test Organisms Using Semi-Purified Bacteriocin
2.14. Evaluation of Bacterial Cell Lysis of Non-Metabolically Active Test Organisms Using Semi-Purified Bacteriocins
2.15. Evaluation of Viable Cell Count Reduction Using Semi-Purified Bacteriocins
3. Results
3.1. Isolation and Identification of Putative LAB Strains and Screening for Bacteriocin Production
3.2. Evaluation of Nature, Integrity and Stability of the Bioactive Molecule
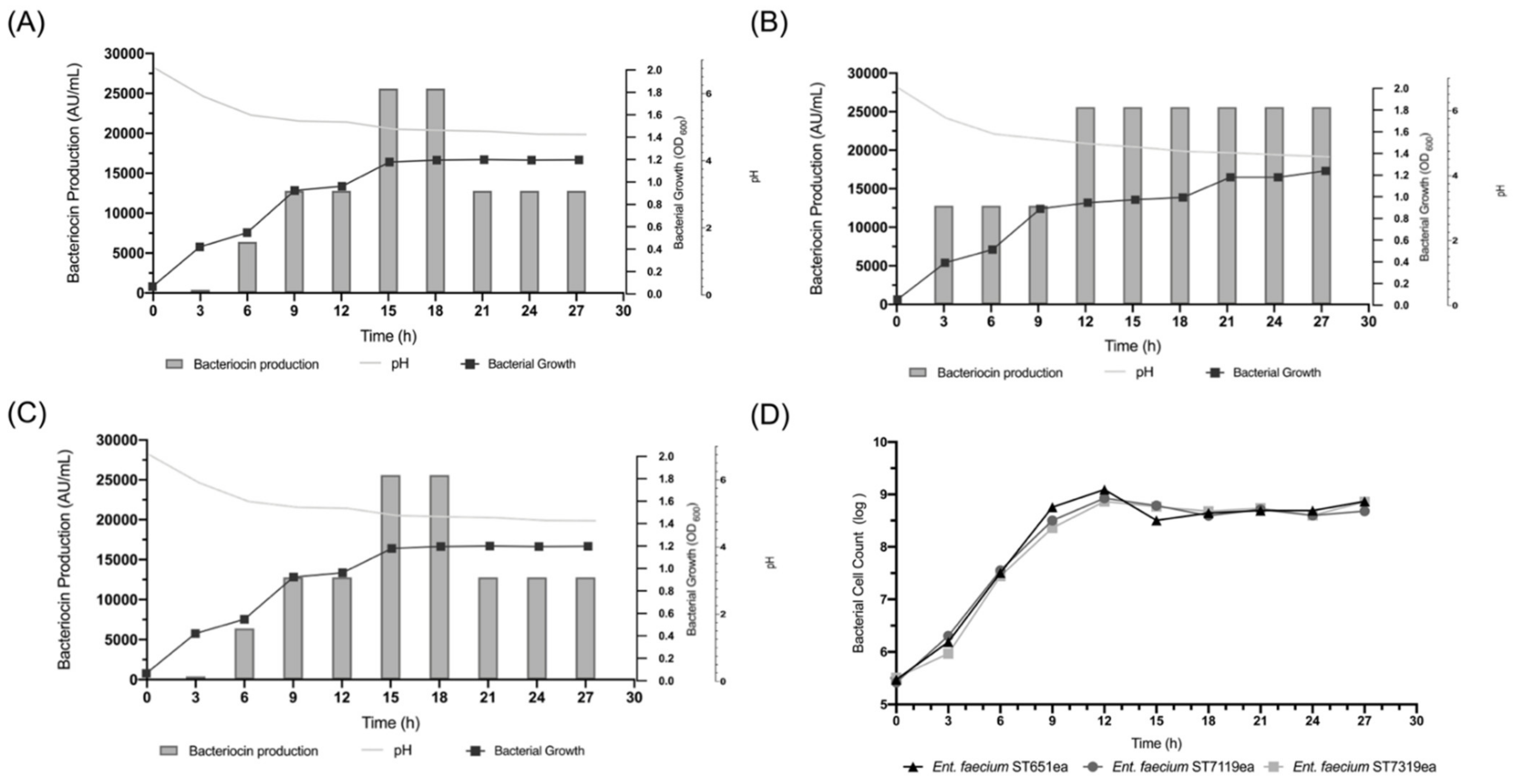
3.3. Bacteriocin Production, Growth Kinetics and Fermentation
3.4. Effect of Varying Incubation Temperature on Bacteriocin Production of E. faecium ST651ea, ST7119ea and ST7319ea
3.5. Screening for Bacteriocin Associated Genes
3.6. Bacterial Growth Inhibition (Cell Lysis) and Spectrum of Activity
3.7. Quantification of Adhered Bacteriocin on the Producer’s Cell Surface
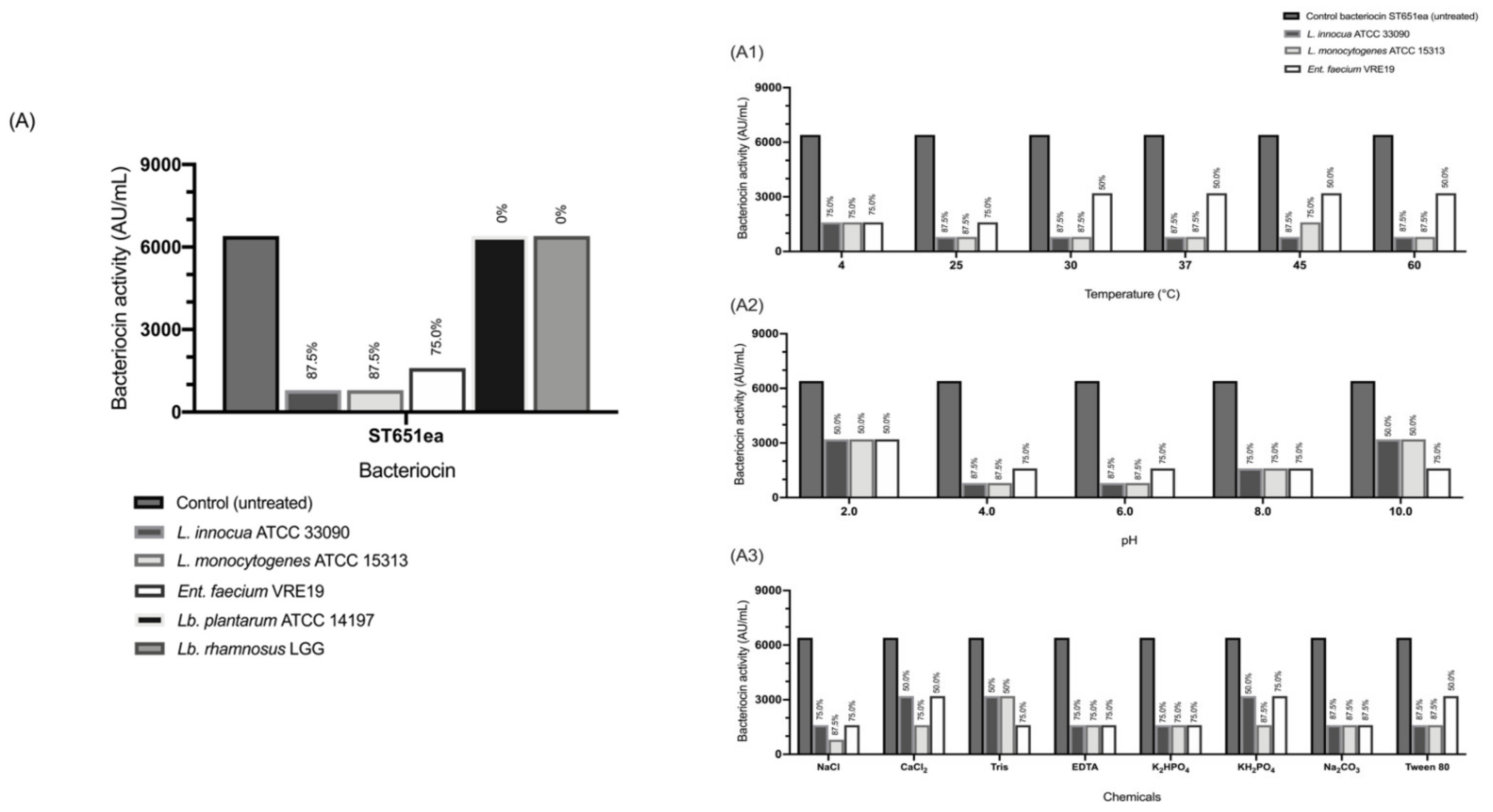
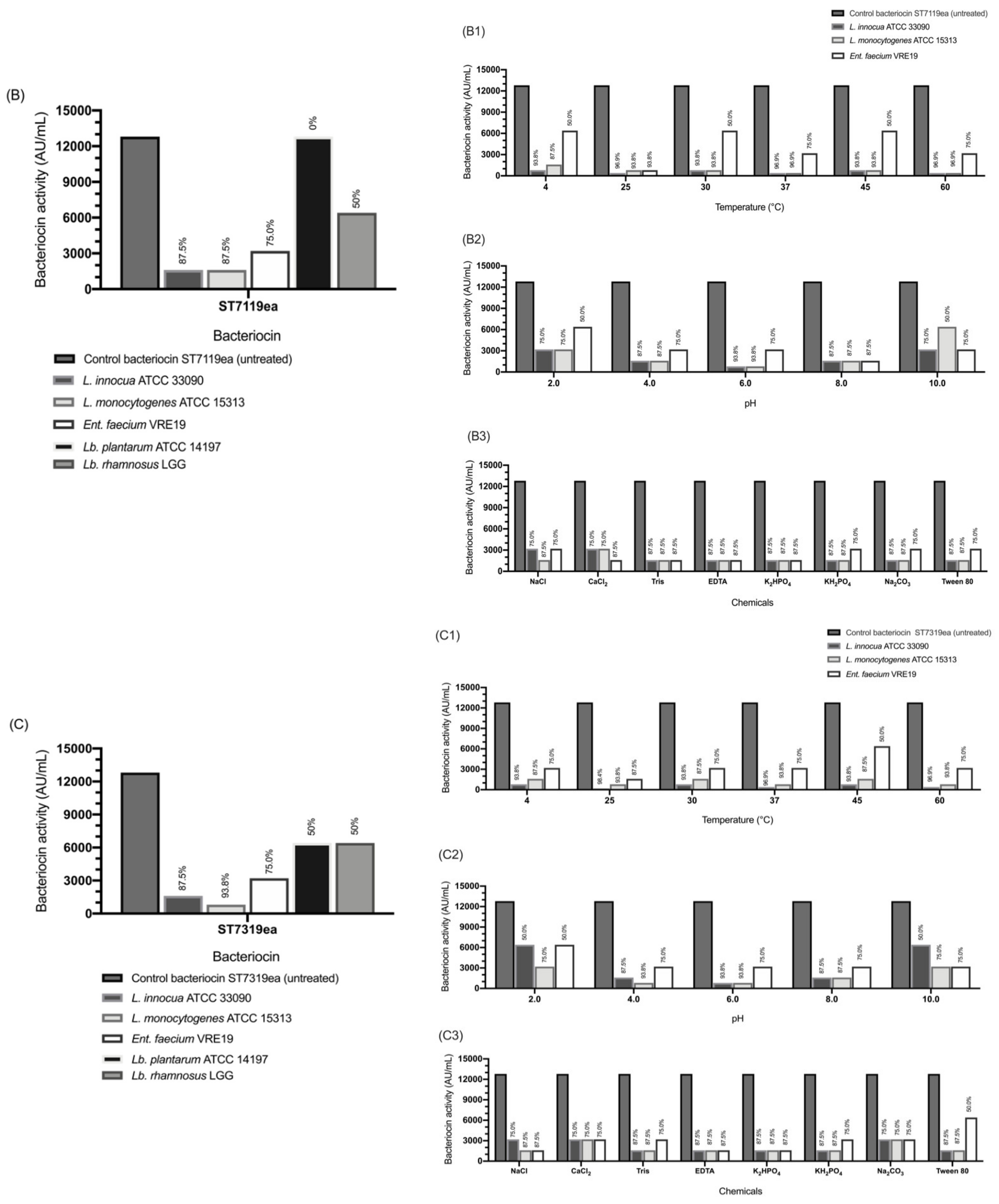
3.8. Adsorption of Bacteriocin to the Target Cells
3.9. Factors Affecting the Adsorption of Bacteriocins to Sensitive Test Organisms
3.10. Bacteriocin Partial Purification
3.11. Molecular Size Estimation of Bacteriocins by Tricine-SDS-PAGE
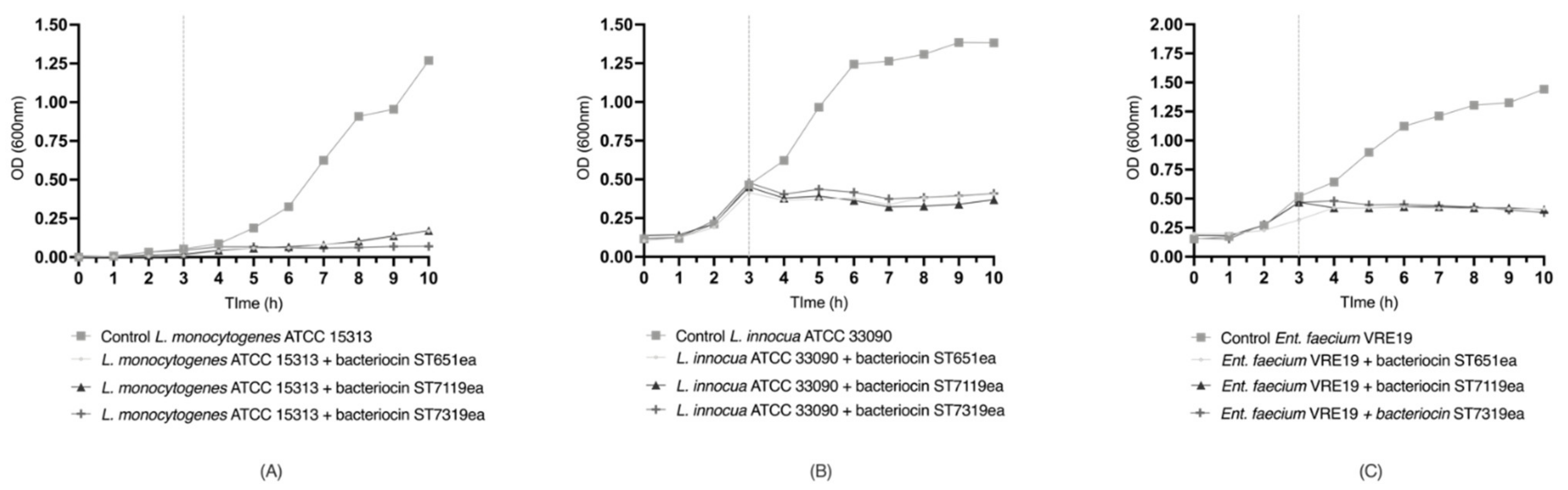
3.12. Evaluation of Bacterial Growth Inhibition of Test Organisms Using Semi-Purified Bacteriocins
3.13. Evaluation of Bacterial Cell Lysis in Non-Metabolically Active Test Organisms Using Semi-Purified Bacteriocins
3.14. Quantification of Viable Cells of Test Organisms after Treatment of Semi-Purified Bacteriocins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Foulquié Moreno, M.R.; Sarantinopoulos, P.; Tsakalidou, E.; De Vuyst, L. The role and application of enterococci in food and health. Int. J. Food Microbiol. 2006, 106, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Hayaloglu, A.A. Comparisons of different single-strain starter cultures for their effects on ripening and grading of Beyaz cheese. Int. J. Food Sci. Technol. 2007, 42, 930–938. [Google Scholar] [CrossRef]
- Litopoulou-Tzanetaki, E.; Tzanetakis, N.; Vafopoulou-Mastrojiannaki, A. Effect of the type of lactic starter on microbiologicalchemical and sensory characteristics of Feta cheese. Food Microbiol. 1993, 10, 31–41. [Google Scholar] [CrossRef]
- Franz, C.M.A.P.; Stiles, M.E.; Schleifer, K.H.; Holzapfel, W.H. Enterococci in foods—A conundrum for food safety. Int. J. Food Microbiol. 2003, 88, 105–122. [Google Scholar] [CrossRef]
- Franz, C.M.A.P.; Specht, I.; Haberer, P.; Holzapfel, W.H. Bile Salt Hydrolase Activity of Enterococci Isolated from Food: Screening and Quantitative Determination. J. Food Prot. 2001, 64, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Moellering, R.C. Emergence of Enterococcus as a Significant Pathogen. Clin. Infect. Dis. 1992, 14, 1173–1176. [Google Scholar] [CrossRef] [PubMed]
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The Genus Enterococcus: Between Probiotic Potential and Safety Concerns—An Update. Front. Microbiol. 2018, 9, 1791. [Google Scholar] [CrossRef] [PubMed]
- Ouwehand, A.; Vesterlund, S. Chapter 11: Antimicrobial Components from Lactic Acid Bacteria: Classification and Physiology. In Lactic Acid Bacteria: Microbial and Functional Aspects, 3rd ed.; Revised and Expanded. Food Science and Technology; Salminen, S., von Wright, A., Ouwehand, A., Eds.; CRC Press, Taylor and Francis Group: Oxford, UK, 2004; pp. 390–410. [Google Scholar]
- De Vuyst, L.D.; Vandamme, E. Chapter 1: Lactic Acid Bacteria and Bacteriocins: Their Practical Importance. In Bacteriocins of Lactic Acid Bacteria: Microbiology, Genetics and Applications; Vuyst, L.D., Vandamme, E., Eds.; Laboratory of Industrial Microbiology and Biocatalysis, University of Ghent: Ghent, Belgium, 1994; ISBN 978-l-4615-2668-l. [Google Scholar]
- Klaenhammer, T. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol. Rev. 1993, 12, 39–85. [Google Scholar] [CrossRef]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Ness, I.F.; Diep, D.B.; Ike, Y. Enterococcal Bacteriocins and Antimicrobial Proteins that Contribute to Niche Control. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M.S., Clewell, D.B., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. Available online: http://www.ncbi.nlm.nih.gov/books/NBK190428/ (accessed on 17 November 2020).
- Hickey, R.M.; Twomey, D.P.; Ross, R.P.; Hill, C. Production of enterolysin A by a raw milk enterococcal isolate exhibiting multiple virulence factors. Microbiology 2003, 149 Pt 3, 655–664. [Google Scholar] [CrossRef] [Green Version]
- Nilsen, T.; Nes, I.F.; Holo, H. Enterolysin A, a cell wall-degrading bacteriocin from Enterococcus faecalis LMG 2333. Appl. Environ. Microbiol. 2003, 69, 2975–2984. [Google Scholar] [CrossRef] [Green Version]
- Tomita, H.; Kamei, E.; Ike, Y. Cloning and Genetic Analyses of the Bacteriocin 41 Determinant Encoded on the Enterococcus faecalis Pheromone-Responsive Conjugative Plasmid pYI14: A Novel Bacteriocin Complemented by Two Extracellular Components (Lysin and Activator). J. Bacteriol. 2008, 190, 2075–2085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giraffa, G.; Carminati, D. Control of Listeria monocytogenes in the rind of Taleggio, a surface-smear cheese, by a bacteriocin from Enterococcus faecium 7C5. Sci. Aliment. 1997, 17, 383–391. [Google Scholar]
- Liu, X.; Vederas, J.C.; Whittal, R.M.; Zheng, J.; Stiles, M.E.; Carlson, D.; Franz, C.M.A.P.; McMullen, L.M.; van Belkum, M.J. Identification of an N-Terminal Formylated, Two-Peptide Bacteriocin from Enterococcus faecalis 710C. J. Agric. Food Chem. 2011, 59, 5602–5608. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, E.; Bednarczyk, A.; Schaeffer, C.; Cai, Y.; Marchioni, E.; Van Dorsselaer, A.; Ennahar, S. Production of Enterocins L50A, L50B, and IT, a New Enterocin, by Enterococcus faecium IT62, a Strain Isolated from Italian Ryegrass in Japan. Antimicrob. Agents Chemother. 2008, 52, 1917–1923. [Google Scholar] [CrossRef] [Green Version]
- Achemchem, F.; Abrini, J.; Martínez-Bueno, M.; Valdivia, E.; Maqueda, M. Control of Listeria monocytogenes in goat’s milk and goat’s jben by the bacteriocinogenic Enterococcus faecium F58 strain. J. Food Prot. 2006, 69, 2370–2376. [Google Scholar] [CrossRef] [PubMed]
- Giraffa, G. Enterococcal bacteriocins: Their potential as anti-Listeria factors in dairy technology. Food Microbiol. 1995, 12, 291–299. [Google Scholar] [CrossRef]
- Aymerich, T.; Garriga, M.; Ylla, J.; Vallier, J.; Monfort, J.M.; Hugas, M. Application of Enterocins as Biopreservatives against Listeria innocua in Meat Products. J. Food Prot. 2000, 63, 721–726. [Google Scholar] [CrossRef]
- Muñoz, A.; Maqueda, M.; Gálvez, A.; Martínez-Bueno, M.; Rodríguez, A.; Valdivia, E. Biocontrol of psychrotrophic enterotoxigenic Bacillus cereus in a nonfat hard cheese by an enterococcal strain-producing enterocin AS-48. J. Food Prot. 2004, 67, 1517–1521. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Hagi, A.; Nii, T.; Tsubotani, Y.; Nakata, H.; Iwata, K. Novel antiseptic compound OPB-2045G shows potent bactericidal activity against methicillin-resistant Staphylococcus aureus and vancomycin-resistant Enterococcus both in vitro and in vivo: A pilot study in animals. J. Med. Microbiol. 2015, 64, 32–36. [Google Scholar] [CrossRef]
- Basanta, A.; Sánchez, J.; Gómez-Sala, B.; Herranz, C.; Hernández, P.E.; Cintas, L.M. Antimicrobial activity of Enterococcus faecium L50, a strain producing enterocins L50 (L50A and L50B), P and Q, against beer-spoilage lactic acid bacteria in broth, wort (hopped and unhopped), and alcoholic and non-alcoholic lager beers. Int. J. Food Microbiol. 2008, 125, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Al Atya, A.K.; Belguesmia, Y.; Chataigne, G.; Ravallec, R.; Vachée, A.; Szunerits, S.; Boukherroub, R.; Drider, D. Anti-MRSA Activities of Enterocins DD28 and DD93 and Evidences on Their Role in the Inhibition of Biofilm Formation. Front. Microbiol. 2016, 7, 817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, T.; Tomita, H.; Ike, Y. Bac 32, a Novel Bacteriocin Widely Disseminated among Clinical Isolates of Enterococcus faecium. Antimicrob. Agents Chemother. 2006, 50, 1202–1212. [Google Scholar] [CrossRef] [Green Version]
- De Vos, P.; Garrity, G.M.; Jones, D.; Kreig, N.R.; Ludwig, W.; Rainey, F.A.; Schleifer, K.-H.; Whitman, W.B. Bergey’s Manual of Systematic Bacteriology: Volume 3. The Firmicutes; Springer: Heidelberg, Germany, 2009. [Google Scholar] [CrossRef]
- Todorov, S.D.; Franco, B.D.G.M.; Tagg, J.R. Bacteriocins of Gram-positive bacteria having activity spectra extending beyond closely related species. Benef. Microbes 2019, 10, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, K.M.O.; de Matos, C.R.; Salles, H.O.; de Melo Franco, B.D.G.; Arellano, K.; Holzapfel, W.H.; Todorov, S.D. Exploring Beneficial/Virulence Properties of Two Dairy-Related Strains of Streptococcus infantarius subsp. infantarius. Probiotics Antimicrob. Proteins 2020, 12, 1524–1541. [Google Scholar] [CrossRef]
- de Moraes, G.M.D.; de Abreu, L.R.; do Egito, A.S.; Salles, H.O.; da Silva, L.M.F.; Nero, L.A.; Todorov, S.D.; dos Santos, K.M.O. Functional properties of Lactobacillus mucosae strains isolated from Brazilian goat milk. Probiotics Antimicrob. Proteins 2016, 9, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.S.; Todorov, S.D.; Ivanova, I.V.; Belguesmia, Y.; Choiset, Y.; Rabesona, H.; Chobert, J.-M.; Haertlé, T.; Franco, B.D.G.M. Characterization of a two-peptide plantaricin produced by Lactobacillus plantarum MBSa4 isolated from Brazilian salami. Food Control 2016, 60, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Aymerich, T.; Holo, H.; Håvarstein, L.S.; Hugas, M.; Garriga, M.; Nes, I.F. Biochemical and genetic characterization of enterocin A from Enterococcus faecium, a new anti-listerial bacteriocin in the pediocin family of bacteriocins. Appl. Environ. Microbiol. 1996, 62, 1676–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- du Toit, M.; Franz, C.M.; Dicks, L.M.; Holzapfel, W.H. Preliminary characterization of bacteriocins produced by Enterococcus faecium and Enterococcus faecalis isolated from pig faeces. J. Appl. Microbiol. 2000, 88, 482–494. [Google Scholar] [CrossRef] [PubMed]
- Cintas, L.M.; Casaus, P.; Håvarstein, L.S.; Hernández, P.E.; Nes, I.F. Biochemical and genetic characterization of enterocin P, a novel sec-dependent bacteriocin from Enterococcus faecium P13 with a broad antimicrobial spectrum. Appl. Environ. Microbiol. 1997, 63, 4321–4330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez, J.; Bourque, D.; Criado, R.; Choi, Y.J.; Cintas, L.M.; Hernández, P.E.; Míguez, C.B. Heterologous extracellular production of enterocin P from Enterococcus faecium P13 in the methylotrophic bacterium Methylobacterium extorquens. FEMS Microbiol. Lett. 2005, 248, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Kruger, M.F.; de Barbosa, M.S.; Miranda, A.; Landgraf, M.; Destro, M.T.; Todorov, S.D.; Franco, B.D.G.M. Isolation of bacteriocinogenic strain of Lactococcus lactis subsp. lactis from rocket salad (Eruca sativa Mill.) and evidence of production of a variant of nisin with modification in the leader-peptide. Food Control 2013, 33, 467–476. [Google Scholar] [CrossRef]
- Todorov, S.D.; Ho, P.; Vaz-Velho, M.; Dicks, L.M.T. Characterization of bacteriocins produced by two strains of Lactobacillus plantarum isolated from Beloura and Chouriço, traditional pork products from Portugal. Meat Sci. 2010, 84, 334–343. [Google Scholar] [CrossRef]
- Yang, R.; Johnson, M.C.; Ray, B. Novel Method to Extract Large Amounts of Bacteriocins from Lactic Acid Bacteriat. Appl. Environ. Microbiol. 1992, 58, 5. [Google Scholar] [CrossRef] [Green Version]
- Biscola, V.; Todorov, S.D.; Capuano, V.S.C.; Abriouel, H.; Gálvez, A.; Franco, B.D.G.M. Isolation and characterization of a nisin-like bacteriocin produced by a Lactococcus lactis strain isolated from charqui, a Brazilian fermented, salted and dried meat product. Meat Sci. 2013, 93, 607–613. [Google Scholar] [CrossRef]
- Todorov, S.; Onno, B.; Sorokine, O.; Chobert, J.M.; Ivanova, I.; Dousset, X. Detection and characterization of a novel antibacterial substance produced by Lactobacillus plantarum ST 31 isolated from sourdough. Int. J. Food Microbiol. 1999, 48, 167–177. [Google Scholar] [CrossRef]
- Schägger, H.; von Jagow, G. Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef]
- Powell, J.E.; Witthuhn, R.C.; Todorov, S.D.; Dicks, L.M.T. Characterization of bacteriocin ST8KF produced by a kefir isolate Lactobacillus plantarum ST8KF. Int. Dairy J. 2007, 17, 190–198. [Google Scholar] [CrossRef]
- Todorov, S.D.; Dicks, L.M.T. Bacteriocin production by Pediococcus pentosaceus isolated from marula (Scerocarya birrea). Int. J. Food Microbiol. 2009, 132, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Kjos, M.; Nes, I.F.; Diep, D.B. Mechanisms of Resistance to Bacteriocins Targeting the Mannose Phosphotransferase System. Appl. Environ. Microbiol. 2011, 77, 3335–3342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gravesen, A.; Jydegaard Axelsen, A.-M.; Mendes da Silva, J.; Hansen, T.B.; Knøchel, S. Frequency of Bacteriocin Resistance Development and Associated Fitness Costs in Listeria monocytogenes. Appl. Environ. Microbiol. 2002, 68, 756–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gravesen, A.; Ramnath, M.; Rechinger, K.B.; Andersen, N.; Jänsch, L.; Héchard, Y.; Hastings, J.W.; Knøchel, S. High-level resistance to class IIa bacteriocins is associated with one general mechanism in Listeria monocytogenes. Microbiology 2002, 148, 2361–2369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macwana, S.; Muriana, P.M. Spontaneous bacteriocin resistance in Listeria monocytogenes as a susceptibility screen for identifying different mechanisms of resistance and modes of action by bacteriocins of lactic acid bacteria. J. Microbiol. Methods 2012, 88, 7–13. [Google Scholar] [CrossRef]
- Pot, B.; Ludwig, W.; Kersters, K.; Schleifer, K.H. Chapter 2: Taxonomy of Lactic Acid Bacteria. In Bacteriocins of Lactic Acid Bacteria: Microbiology, Genetics and Applications; Vuyst, L.D., Vandamme, E., Eds.; Laboratory of Industrial Microbiology and Biocatalysis, University of Ghent: Ghent, Belgium, 1994; ISBN 978-l-4615-2668-l. [Google Scholar]
- Franz, C.M.A.P.; Van Belkum, M.J.; Holzapfel, W.H.; Abriouel, H.; Gálvez, A. Diversity of enterococcal bacteriocins and their grouping in a new classification scheme. FEMS Microbiol. Rev. 2007, 31, 293–310. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.; Jeong, D.-W.; Lee, J.-H. Safety and biotechnological properties of Enterococcus faecalis and Enterococcus faecium isolates from Meju. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 813–820. [Google Scholar] [CrossRef]
- Onda, T.; Yanagida, F.; Uchimura, T.; Tsuji, M.; Ogino, S.; Shinohara, T.; Yokotsuka, K. Widespread distribution of the bacteriocin-producing lactic acid cocci in miso-paste products. J. Appl. Microbiol. 2002, 92, 695–705. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, V.Q.; Camargo, A.C.; Todorov, S.D.; Nero, L.A. Potential Control of Listeria monocytogenes by Bacteriocinogenic Enterococcus hirae ST57ACC and Pediococcus pentosaceus ST65ACC Strains Isolated from Artisanal Cheese. Probiot. Antimicrob. Proteins 2019, 11, 696–704. [Google Scholar] [CrossRef]
- Gálvez, A.; Valdivia, E.; Abriouel, H.; Camafeita, E.; Mendez, E.; Martínez-Bueno, M.; Maqueda, M. Isolation and characterization of enterocin EJ97, a bacteriocin produced by Enterococcus faecalis EJ97. Arch. Microbiol. 1998, 171, 59–65. [Google Scholar] [CrossRef]
- Lauková, A.; Chrastinová, L.; Pogány Simonová, M.; Strompfová, V.; Plachá, I.; Čobanová, K.; Formelová, Z.; Chrenková, M.; Ondruška, L. Enterococcus faecium AL 41: Its Enterocin M and Their Beneficial Use in Rabbits Husbandry. Probiot. Antimicrob. Proteins 2012, 4, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Mills, S.; Ross, R.P.; Hill, C. Bacteriocins and bacteriophage; a narrow-minded approach to food and gut microbiology. FEMS Microbiol. Rev. 2017, 41 (Suppl. S1), S129–S153. [Google Scholar] [CrossRef]
- Todorov, S.D.; Dicks, L.M.T. Pediocin ST18, an anti-listerial bacteriocin produced by Pediococcus pentosaceus ST18 isolated from boza, a traditional cereal beverage from Bulgaria. Process Biochem. 2005, 40, 365–370. [Google Scholar] [CrossRef]
- Dillon, V.M. Natural Anti-microbial Systems|Preservative Effects during Storage. In Encyclopedia of Food Microbiology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 941–947. [Google Scholar] [CrossRef]
- Kawamoto, S.; Shima, J.; Sato, R.; Eguchi, T.; Ohmomo, S.; Shibato, J.; Horikoshi, N.; Takeshita, K.; Sameshima, T. Biochemical and Genetic Characterization of Mundticin KS, an Antilisterial Peptide Produced by Enterococcus mundtii NFRI 7393. Appl. Environ. Microbiol. 2002, 68, 3830–3840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saavedra, L.; Minahk, C.; de Ruiz Holgado, A.P.; Sesma, F. Enhancement of the Enterocin CRL35 Activity by a Synthetic Peptide Derived from the NH2-Terminal Sequence. Antimicrob. Agents Chemother. 2004, 48, 2778–2781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todorov, S.D.; Rachman, C.; Fourrier, A.; Dicks, L.M.T.; van Reenen, C.A.; Prévost, H.; Dousset, X. Characterization of a bacteriocin produced by Lactobacillus sakei R1333 isolated from smoked salmon. Anaerobe 2011, 17, 23–31. [Google Scholar] [CrossRef]
- Kang, J.H.; Lee, M.S. Characterization of a bacteriocin produced by Enterococcus faecium GM-1 isolated from an infant. J. Appl. Microbiol. 2005, 98, 1169–1176. [Google Scholar] [CrossRef]
- Birri, D.J.; Brede, D.A.; Tessema, G.T.; Nes, I.F. Bacteriocin Production, Antibiotic Susceptibility and Prevalence of Haemolytic and Gelatinase Activity in Faecal Lactic Acid Bacteria Isolated from Healthy Ethiopian Infants. Microb. Ecol. 2013, 65, 504–516. [Google Scholar] [CrossRef] [PubMed]
- De Kwaadsteniet, M.; Fraser, T.; Van Reenen, C.A.; Dicks, L.M.T. Bacteriocin T8, a Novel Class IIa sec-Dependent Bacteriocin Produced by Enterococcus faecium T8, Isolated from Vaginal Secretions of Children Infected with Human Immunodeficiency Virus. Appl. Environ. Microbiol. 2006, 72, 4761–4766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flannagan, S.E.; Clewell, D.B.; Sedgley, C.M. A “retrocidal” plasmid in Enterococcus faecalis: Passage and protection. Plasmid 2008, 59, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, J.; Diep, D.B.; Herranz, C.; Nes, I.F.; Cintas, L.M.; Hernández, P.E. Amino acid and nucleotide sequence, adjacent genes, and heterologous expression of hiracin JM79, a sec-dependent bacteriocin produced by Enterococcus hirae DCH5, isolated from Mallard ducks (Anas platyrhynchos). FEMS Microbiol. Lett. 2007, 270, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Todorov, S.D.; Dicks, L.M.T. Optimization of bacteriocin ST311LD production by Enterococcus faecium ST311LD, isolated from spoiled black olives. J. Microbiol. 2005, 43, 370–374, ISSN 1225-8873. [Google Scholar] [PubMed]
- Gardini, F.; Martuscelli, M.; Caruso, M.C.; Galgano, F.; Crudele, M.A.; Favati, F.; Guerzoni, M.E.; Suzzi, G. Effects of pH, temperature and NaCl concentration on the growth kinetics, proteolytic activity and biogenic amine production of Enterococcus faecalis. Int. J. Food Microbiol. 2001, 64, 105–117. [Google Scholar] [CrossRef]
- Yang, E.; Fan, L.; Yan, J.; Jiang, Y.; Doucette, C.; Fillmore, S.; Walker, B. Influence of culture media, pH and temperature on growth and bacteriocin production of bacteriocinogenic lactic acid bacteria. AMB Express 2018, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todorov, S.D.; Wachsman, M.; Tomé, E.; Dousset, X.; Destro, M.T.; Dicks, L.M.T.; de Melo Franco, B.D.G.; Vaz-Velho, M.; Drider, D. Characterization of an antiviral pediocin-like bacteriocin produced by Enterococcus faecium. Food Microbiol. 2010, 27, 869–879. [Google Scholar] [CrossRef] [PubMed]
- O’Keeffe, T.; Hill, C.; Ross, R.P. Characterization and Heterologous Expression of the Genes Encoding Enterocin a Production, Immunity, and Regulation in Enterococcus faecium DPC1146. Appl. Environ. Microbiol. 1999, 65, 1506–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.G.; Becattini, S.; Moody, T.U.; Shliaha, P.V.; Littmann, E.R.; Seok, R.; Gjonbalaj, M.; Eaton, V.; Fontana, E.; Amoretti, L.; et al. Microbiota-derived lantibiotic restores resistance against vancomycin-resistant Enterococcus. Nature 2019, 572, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Melander, R.J.; Zurawski, D.V.; Melander, C. Narrow-spectrum antibacterial agents. Med. Chem. Commun. 2018, 9, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Furtado, D.N.; Favaro, L.; Nero, L.A.; Franco, B.D.G.M.; Todorov, S.D. Nisin Production by Enterococcus hirae DF105Mi Isolated from Brazilian Goat Milk. Probiot. Antimicrob. Proteins 2019, 11, 1391–1402. [Google Scholar] [CrossRef] [PubMed]
- Yıldırım, Z.; Avs¸ar, Y.K.; Yıldırım, M. Factors affecting the adsorption of buchnericin LB, a bacteriocin produced by Lactocobacillus buchneri. Microbiol. Res. 2002, 157, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Todorov, S.D. Bacteriocin production by Lactobacillus plantarum AMA-K isolated from Amasi, a Zimbabwean fermented milk product and study of the adsorption of bacteriocin AMA-K to Listeria sp. Braz. J. Microbiol. 2008, 39, 178–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iseppi, R.; Stefani, S.; de Niederhausern, S.; Bondi, M.; Sabia, C.; Messi, P. Characterization of Anti-Listeria monocytogenes Properties of two Bacteriocin-Producing Enterococcus mundtii Isolated from Fresh Fish and Seafood. Curr. Microbiol. 2019, 76, 1010–1019. [Google Scholar] [CrossRef] [PubMed]
- Todorov, S.D.; Dicks, L.M.T. Evaluation of lactic acid bacteria from kefir, molasses and olive brine as possible probiotics based on physiological properties. Ann. Microbiol. 2008, 58, 661–670. [Google Scholar] [CrossRef]
- Cintas, L.M.; Casaus, P.; Herranz, C.; Håvarstein, L.S.; Holo, H.; Ndez, P.E.H.; Nes, I.F. Biochemical and Genetic Evidence that Enterococcus faecium L50 Produces Enterocins L50A and L50B, the sec-Dependent Enterocin P, and a Novel Bacteriocin Secreted without an N-Terminal Extension Termed Enterocin Q. J. Bacteriol. 2000, 182, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joerger, M.C.; Klaenhammer, T.R. Characterization and purification of helveticin J and evidence for a chromosomally determined bacteriocin produced by Lactobacillus helveticus 481. J. Bacteriol. 1986, 167, 439–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vuyst, L.; Callewaert, R.; Crabbe, K. Primary metabolite kinetics of bacteriocin biosynthesis by Lactobacillus amylovorus and evidence for stimulation of bacteriocin production under unfavourable growth conditions. Microbiology 1996, 142, 817–827. [Google Scholar] [CrossRef] [Green Version]
- Meincken, M.; Todorov, S.T. Study the effect of bacteriocins on target cell by atomic force microscopy, a new method for visualisation of the mode of action. Trakia J. Sci. 2009, 7, 28–32. [Google Scholar]
- Ike, Y.; Clewell, D.B. Genetic analysis of the pAD1 pheromone response in Streptococcus faecalis, using transposon Tn917 as an insertional mutagen. J. Bacteriol. 1984, 158, 777–783. [Google Scholar] [CrossRef] [Green Version]
- Ennahar, S.; Sashihara, T.; Sonomoto, K.; Ishizaki, A. Class IIa bacteriocins: Biosynthesis, structure and activity. FEMS Microbiol. Rev. 2000, 24, 85–106. [Google Scholar] [CrossRef]
- De Vuyst, L.; Foulquié Moreno, M.R.; Revets, H. Screening for enterocins and detection of hemolysin and vancomycin resistance in enterococci of different origins. Int. J. Food Microbiol. 2003, 84, 299–318. [Google Scholar] [CrossRef]
- Deegan, L.H.; Cotter, P.D.; Hill, C.; Ross, P. Bacteriocins: Biological tools for bio-preservation and shelf-life extension. Int. Dairy J. 2006, 16, 1058–1071. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteriocin | Primer | Oligonucleotide Sequence (5′–3′) | Product Size (bp) | References |
|---|---|---|---|---|
| Enterocin A | entA-F | GAG ATT TAT CTC CAT AAT CT | 452 | [32] |
| entA-R | GTA CCA CTC ATA GTG GAA | |||
| Enterocin B | entB-F | GAA AAT GAT CAC AGA ATG CCT A | 159 | [33] |
| entB-R | GTT GCA TTT AGA GTA TAC ATT TG | |||
| Enterocin L50B | entL50B-F | ATG GGA GCA ATC GCA AAA TTA | 135 | [34] |
| entL50B-R | TAG CCA TTT TTC AAT TTG ATC | |||
| Enterocin P | entP-F | ATG AGA AAA AAA TTA TTT AGT TT | 216 | [35] |
| entP-R | TTA ATG TCC CAT ACC TGC CAA ACC | |||
| Nisin | Nis-F | ATG AGT ACA AAA GAT TTCAAC TT | 203 | [36] |
| Nis-R | TTA TTT GCT TAC GTG AAC GC | |||
| Pediocin | PedPro-F | CAA GAT CGT TAA CCA GTT T | 1238 | [37] |
| Ped1041-R | CCG TTG TTC CCA TAG TCT AA |
| Strain ID | Identified Bacteriocin Genes | Protein Sequences | |
|---|---|---|---|
| E. faecium ST651ea | entB | Reconstructed sequences | ENDHRMPNELNRPNNLSKGGAKCGAAIAGGLFGIP KGPLA WAAGLANVYS KCN |
| GenBank: ADR70740.1 | WSKCGAAIAGGLFGIPKPLAWAAGLANVYSKCK | ||
| entP | Reconstructed sequences | MRKKLFSLTL IGKFGLVVTN FGTKVDAATR SYDNGIYCNNS KCWVNWGEAKE NIAGIVISGW ASGLAGMGH | |
| GenBank: ERK34332.1 | MRKKLFSLTL IGKFGLVVTN FGTKVDAATR SYGNGVYCNNS KCWVNWGEAKENIAGIVIS GWASGLAGIGH | ||
| E. faecium ST7119ea | entA | Reconstructed sequences | SKDPKYSDI LEVLQKVYLK LEKQKYELDP GPLINRLVN- - - -TAYTN KIRFTEYQEELIRNLSEIGRT AGINGLYRA DYG |
| GenBank: AP8904224.1 | MKKNAKQIVH ELYNDISISK DPKYSDILEV LQKVYLKLEKQ KYELDPGPLI NRLVNYLYFT AYTNKIRFTE YQEELIRNLSE | ||
| entB | Reconstructed sequences | ENDHRMPNELN RPNNLSKGGAKCGAAIAGGLFGI PKGPLAWAAGLANVYSKCN | |
| GenBank: ADR70740.1 | WSKCGAAIAGGLFGI PKGPLAWAAGLANVYSKCK | ||
| E. faecium ST7319ea | entA | Reconstructed sequences | GSAK MKKNAKQIVHELYNDISISKDPKYSDILEVL QKVYLKLEKQKYELDPGPLINRLVNYLYFT-YTN KIRFTEYQEELIRNLSEIGRTAGINGLYRADYGE |
| GenBank: AP8904224.1 | MKKNAKQIVHELYNDISISKDPKYSDILEVL QKVYLKLEKQKYELDPGPLINRLVNYLYFTAY TNKIRFTEYQEELIRNLSE | ||
| entB | Reconstructed sequences | ENDHRMPNELNRPNNLSKGGAKCGAAIAG GLFGIPKGPLALVAGLANVYSKCN | |
| GenBank: ADR70740.1 | WSKCGAAIAGGLFGIPKGPLAWAAGLANVYSKCK | ||
| Test Organism | Number of Strains with Positive Inhibition/Total Number of Strains | ||
|---|---|---|---|
| E. faecium ST651ea | E. faecium ST7119ea | E. faecium ST7319ea | |
| Enterococcus spp. | |||
| E. aviumc | 0/3 | 0/3 | 0/3 |
| E. duransc | 0/1 | 0/1 | 0/1 |
| E. gallinarumc | 0/1 | 0/1 | 0/1 |
| E. faeciumc | 10/12 | 12/12 | 11/12 |
| E. faecalisc | 2/2 | 2/2 | 2/2 |
| E. thailandicusc | 1/1 | 1/1 | 1/1 |
| Vancomycin-resistant Enterococcus spp. | |||
| E. lactisd | 1/1 | 1/1 | 0/1 |
| E. faeciumd | 9/11 | 7/11 | 5/11 |
| VRE clinical isolates d | 15/19 | 18/19 | 19/19 |
| Lactobacillus spp. | |||
| Lb. brevisc | 0/3 | 0/3 | 0/3 |
| Lb. coryniformisc | 0/1 | 0/1 | 0/1 |
| Lb. curvatusc | 0/2 | 0/2 | 0/2 |
| Lb. fermentumc | 0/2 | 0/2 | 0/2 |
| Lb. paracaseic | 1/5 | 1/5 | 1/5 |
| Lb. plantarumac | 0/9 | 0/9 | 0/9 |
| Lb. reuteric | 0/2 | 0/2 | 0/2 |
| Lb. rhamnosusc | 0/5 | 0/5 | 0/5 |
| Lb. rhamnosus LGG a | 0/1 | 0/1 | 0/1 |
| Lb. sakeic | 1/5 | 1/5 | 1/5 |
| Lb. salivariusc | 0/3 | 0/3 | 0/3 |
| Lactococcus spp. | |||
| Lc. lactisc | 0/2 | 0/2 | 0/2 |
| Leuconostoc spp. | |||
| Leuc. citreumc | 0/1 | 0/1 | 0/1 |
| Leuc. mesenteroidesce | 0/7 | 0/7 | 0/7 |
| Listeria spp. | |||
| L. monocytogenesa | 1/1 | 1/1 | 1/1 |
| L. innocuaa | 1/1 | 1/1 | 1/1 |
| Pediococcus spp. | |||
| P. acidilacticice | 0/4 | 0/4 | 0/4 |
| P. pentosaceusc | 1/3 | 0/3 | 0/3 |
| Staphylococcus spp. | |||
| S. arlettaeb | 0/1 | 0/1 | 0/1 |
| S. aureusb | 0/1 | 0/1 | 0/1 |
| S. auricularisb | 0/1 | 0/1 | 0/1 |
| S. capitis subsp. capitis b | 0/1 | 0/1 | 0/1 |
| S. carnosus subsp. carnosus b | 0/1 | 0/1 | 0/1 |
| S. cohnii subsp. cohnii b | 0/1 | 0/1 | 0/1 |
| S. delphinib | 0/1 | 0/1 | 0/1 |
| S. epidermidisb | 0/2 | 0/2 | 0/2 |
| S. haemolyticusb | 0/1 | 0/1 | 0/1 |
| S. hominisb | 0/1 | 0/1 | 0/1 |
| S. aureus MRSA ad | 0/6 | 0/6 | 0/6 |
| S. lentusb | 0/1 | 0/1 | 0/1 |
| S. succinusb | 0/1 | 0/1 | 0/1 |
| S. warnerib | 0/1 | 0/1 | 0/1 |
| Streptococcus spp. | |||
| Str. gordoniib | 0/1 | 0/1 | 0/1 |
| Str. mitisb | 0/1 | 0/1 | 0/1 |
| Str. mutansb | 0/1 | 0/1 | 0/1 |
| Str. sanguinisb | 0/1 | 0/1 | 0/1 |
| Str. salivarius subsp. Salivarius b | 0/1 | 0/1 | 0/1 |
| Str. sanguinisb | 0/1 | 0/1 | 0/1 |
| Str. thermophilusb | 0/1 | 0/1 | 0/1 |
| Weisella | |||
| W. cibariac | 0/1 | 0/1 | 0/1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fugaban, J.I.I.; Vazquez Bucheli, J.E.; Holzapfel, W.H.; Todorov, S.D. Characterization of Partially Purified Bacteriocins Produced by Enterococcus faecium Strains Isolated from Soybean Paste Active Against Listeria spp. and Vancomycin-Resistant Enterococci. Microorganisms 2021, 9, 1085. https://doi.org/10.3390/microorganisms9051085
Fugaban JII, Vazquez Bucheli JE, Holzapfel WH, Todorov SD. Characterization of Partially Purified Bacteriocins Produced by Enterococcus faecium Strains Isolated from Soybean Paste Active Against Listeria spp. and Vancomycin-Resistant Enterococci. Microorganisms. 2021; 9(5):1085. https://doi.org/10.3390/microorganisms9051085
Chicago/Turabian StyleFugaban, Joanna Ivy Irorita, Jorge Enrique Vazquez Bucheli, Wilhelm Heinrich Holzapfel, and Svetoslav Dimitrov Todorov. 2021. "Characterization of Partially Purified Bacteriocins Produced by Enterococcus faecium Strains Isolated from Soybean Paste Active Against Listeria spp. and Vancomycin-Resistant Enterococci" Microorganisms 9, no. 5: 1085. https://doi.org/10.3390/microorganisms9051085
APA StyleFugaban, J. I. I., Vazquez Bucheli, J. E., Holzapfel, W. H., & Todorov, S. D. (2021). Characterization of Partially Purified Bacteriocins Produced by Enterococcus faecium Strains Isolated from Soybean Paste Active Against Listeria spp. and Vancomycin-Resistant Enterococci. Microorganisms, 9(5), 1085. https://doi.org/10.3390/microorganisms9051085