
In Vivo Confirmation of the Antimicrobial Effect of Probiotic Candidates against Gardnerella vaginalis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation
2.2. Antimicrobial Effect of the LAB Metabolites against the Growth of GV
2.3. BV-Induced Mice Model and Administration of the LAB
2.4. RNA Extraction and RT-PCR
2.5. Histopathological Analysis
2.6. Adhesion
2.7. Antibiotic Susceptibility
2.8. Assessment of Enzyme Production
2.9. Analysis of the Lactic Acid Level Using the HPLC-UV Method
2.10. Hemolysis Activity
2.11. Bile Salt Hydrolase Activity
2.12. Statistical Analysis
3. Results
3.1. Antimicrobial Effect of the LAB Metabolites against the Growth of GV
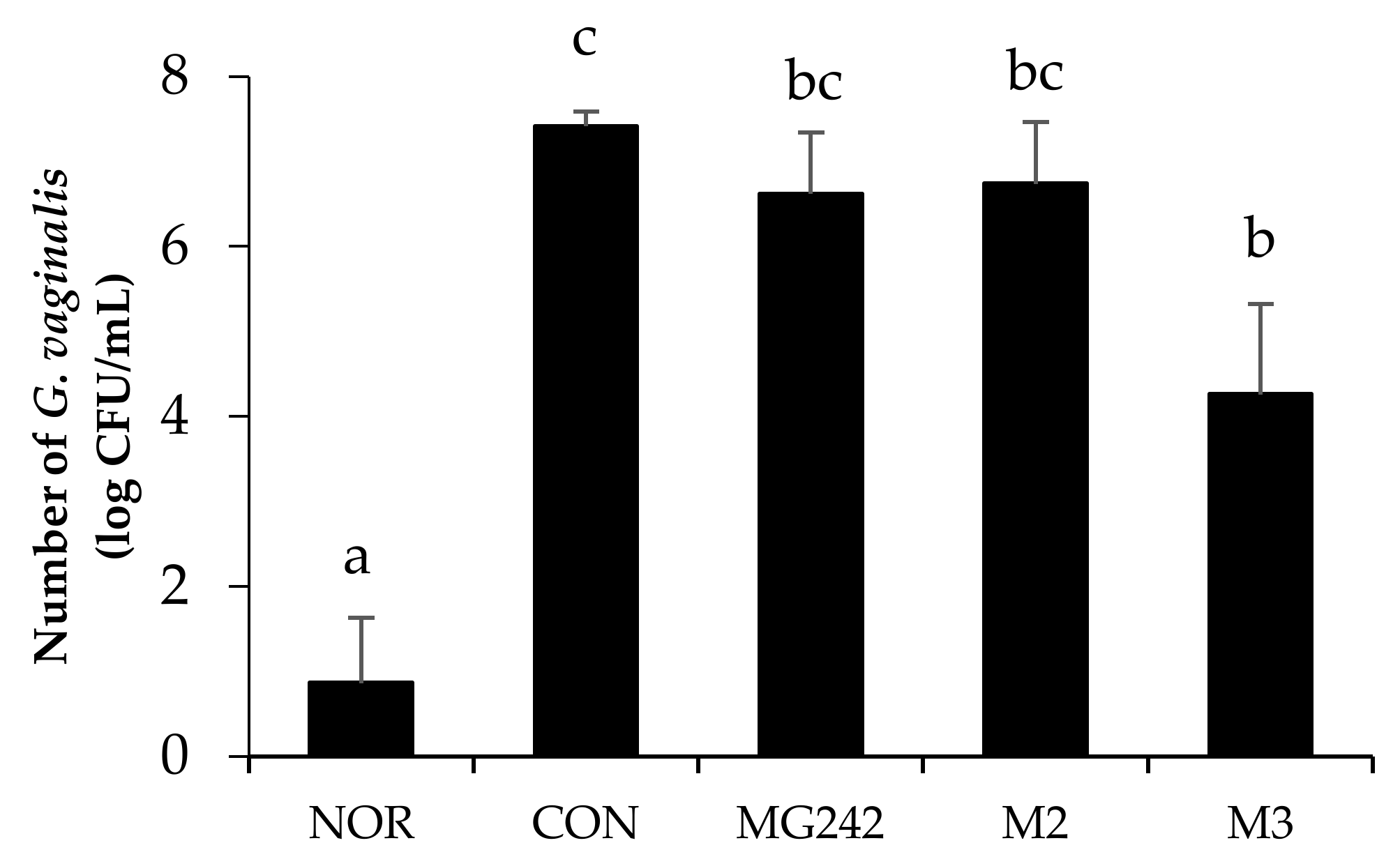
3.2. In Vivo Antimicrobial Effect of the LAB on BV-Induced Mice
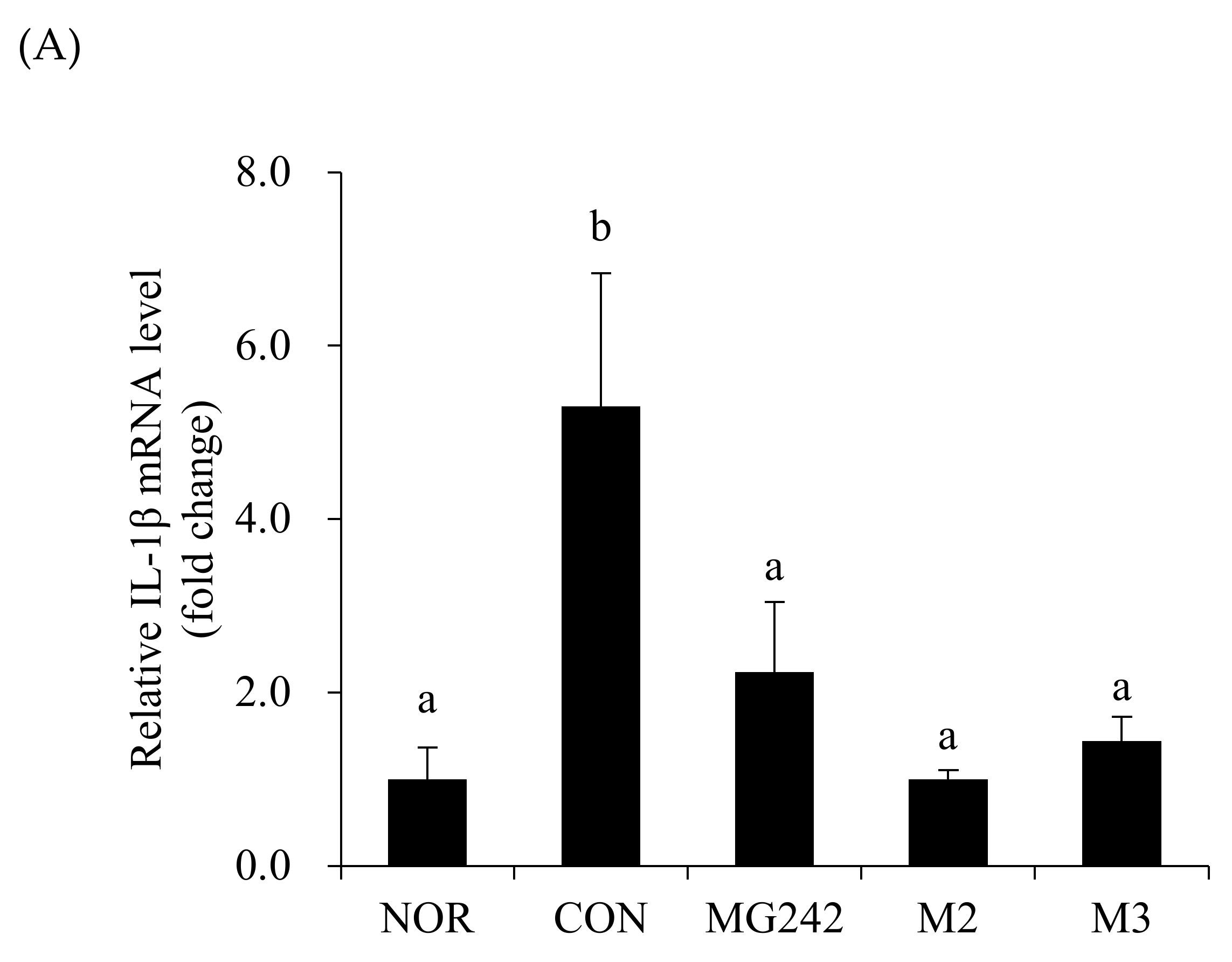
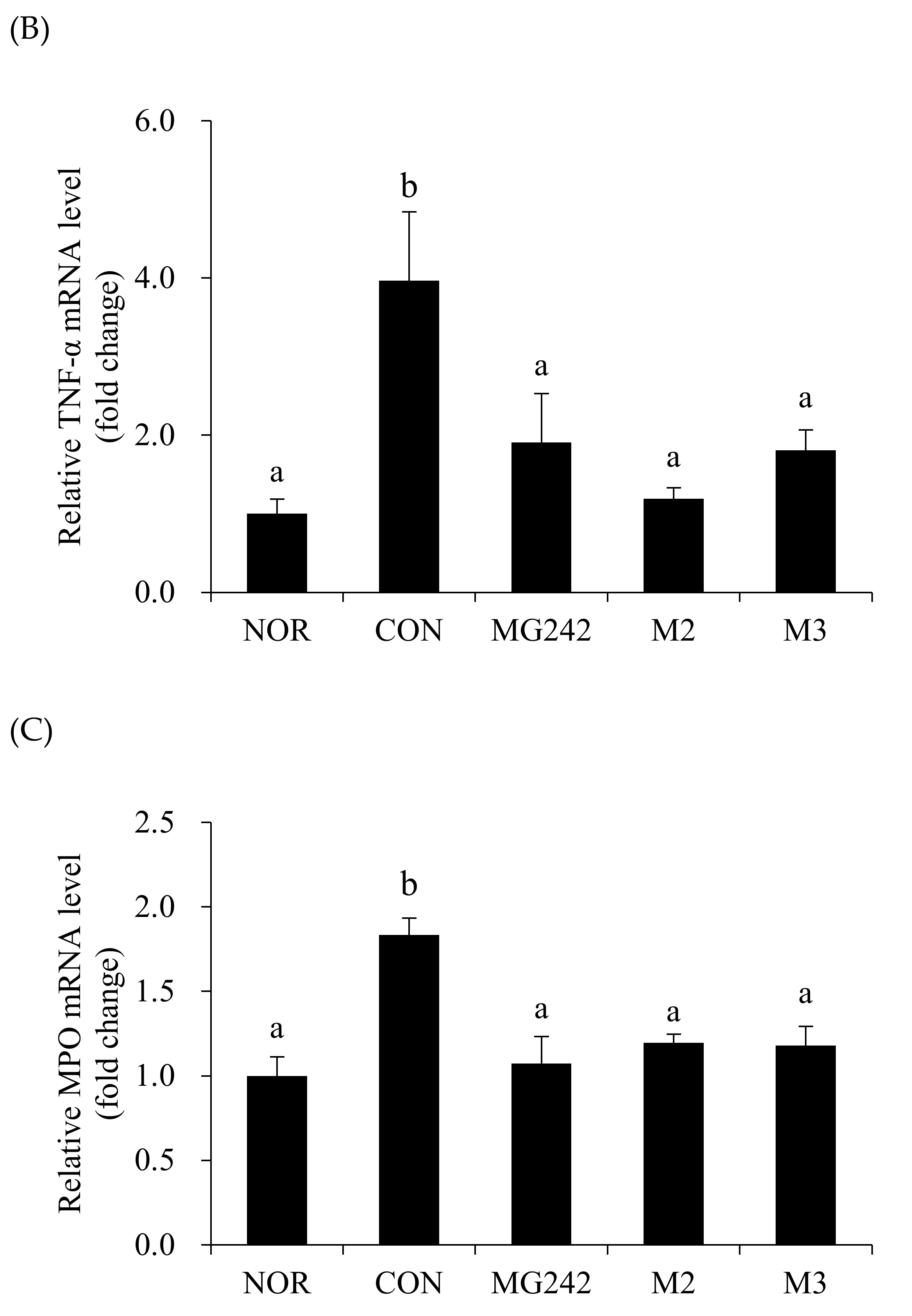
3.3. Pro-Inflammatory Biomarkers of the Vaginal Tissues in BV-Induced Mice
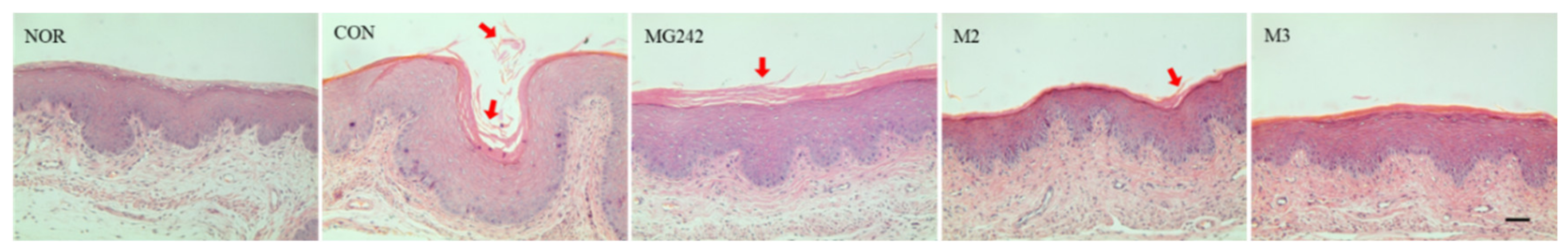
3.4. Histopathological Analysis of BV-Induced Mice
3.5. Fulfillment of the LAB
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Machado, A.; Castro, J.; Cereija, T.; Almeida, C.; Cerca, N.J.P. Diagnosis of bacterial vaginosis by a new multiplex peptide nucleic acid fluorescence in situ hybridization method. PeerJ 2015, 3, e780. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, F.; Fernández-Blázquez, A.; García, B.J. Vaginosis. Vaginal microbiota. Enferm. Infecc. Microbiol. Clin. 2019, 37, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108, 4680–4687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van de Wijgert, J.H.H.M.; Jespers, V. The global health impact of vaginal dysbiosis. Res. Microbiol. 2017, 168, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.-S.; Ehlers, M.M.; Lombaard, H.; Redelinghuys, M.J.; Kock, M.M. Etiology of bacterial vaginosis and polymicrobial biofilm formation. Crit. Rev. Microbiol. 2017, 43, 651–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, D.; Castro, J.; Palmeira-de-Oliveira, A.; Martinez-de-Oliveira, J.; Cerca, N. Bacterial vaginosis biofilms: Challenges to current therapies and emerging solutions. Front. Microbiol. 2016, 6, 1528. [Google Scholar] [CrossRef] [Green Version]
- Hay, S.P.P.; Daniels, D. UK National Guideline for the Management of Bacterial Vaginosis 2012 UK: Clinical Effectiveness Group British Association for Sexual Health and HIV; NHS Evidence: London, UK; Manchester, UK, 2012. [Google Scholar]
- Anukam, K.C.; Osazuwa, E.; Osemene, G.I.; Ehigiagbe, F.; Bruce, A.W.; Reid, G. Clinical study comparing probiotic Lactobacillus GR-1 and RC-14 with metronidazole vaginal gel to treat symptomatic bacterial vaginosis. Microbes Infect. 2006, 8, 2772–2776. [Google Scholar] [CrossRef]
- Fethers, K.A.; Fairley, C.K.; Hocking, J.S.; Gurrin, L.C.; Bradshaw, C.S. Sexual risk factors and bacterial vaginosis: A systematic review and meta-analysis. Clin. Infect. Dis. 2008, 47, 1426–1435. [Google Scholar] [CrossRef]
- Oduyebo, O.O.; Anorlu, R.I.; Ogunsola, F.T. The effects of antimicrobial therapy on bacterial vaginosis in non-pregnant women. Cochrane Database Syst. Rev. 2009, 3. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collado, M.C.; Isolauri, E.; Salminen, S.; Sanz, Y. The impact of probiotic on gut health. Curr. Drug Metab. 2009, 10, 68–78. [Google Scholar] [CrossRef]
- Reid, G.; Bruce, A.W.; Fraser, N.; Heinemann, C.; Owen, J.; Henning, B. Oral probiotics can resolve urogenital infections. FEMS Immunol. Med. Microbiol. 2001, 30, 49–52. [Google Scholar] [CrossRef] [PubMed]
- Boris, S.; Barbés, C. Role played by lactobacilli in controlling the population of vaginal pathogens. Microbes Infect. 2000, 2, 543–546. [Google Scholar] [CrossRef]
- Jang, S.E.; Jeong, J.J.; Choi, S.Y.; Kim, H.; Han, M.J.; Kim, D.H. Lactobacillus rhamnosus HN001 and Lactobacillus acidophilus La-14 attenuate Gardnerella vaginalis-infected bacterial vaginosis in mice. Nutrients 2017, 9, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.E.; Kim, J.K.; Han, S.K.; Jang, S.E.; Han, M.J.; Kim, D.H. Lactobacillus plantarum NK3 and Bifidobacterium longum NK49 Alleviate Bacterial Vaginosis and Osteoporosis in Mice by Suppressing NF-κ B-Linked TNF-α Expression. J. Med. Food 2019, 22, 1022–1031. [Google Scholar] [CrossRef]
- Kang, C.H.; Han, S.H.; Kim, Y.; Paek, N.S.; So, J.S. In vitro probiotic properties of Lactobacillus salivarius MG242 isolated from human vagina. Probiotics Antimicrob. Proteins 2018, 10, 343–349. [Google Scholar] [CrossRef]
- Kang, C.H.; Kim, Y.; Han, S.H.; Kim, J.S.; Paek, N.S.; So, J.S. In vitro probiotic properties of vaginal Lactobacillus fermentum MG901 and Lactobacillus plantarum MG989 against Candida albicans. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 228, 232–237. [Google Scholar] [CrossRef]
- Paek, N.S.; Lee, Y.Y.; Han, S.H.; Kang, C.H.; So, J.S. Characterization and inhibitory activity of Lactobacillus plantarum MG989 and Lactobacillus fermentum MG901 isolated from vaginal microbiota of Korean women against Gardnerella vaginalis and Candida albicans. Korean Soc. Biotechnol. Bioeng. J. 2016, 31, 40–45. [Google Scholar]
- So, J.S.; Paek, N.S. Composition Comprising Lactobacillus fermentum MG901 or Lactobacillus plantarum MG989. KR Patent 10-2016-0026155, 4 March 2016. [Google Scholar]
- Nguyen, T.H.; Kim, Y.; Kim, J.-S.; Jeong, Y.; Park, H.M.; Kim, J.W.; Kim, J.E.; Kim, H.; Paek, N.S.; Kang, C.H. Evaluating the cryoprotective encapsulation of the lactic acid bacteria in simulated gastrointestinal conditions. Biotechnol. Bioprocess Eng. 2020, 25, 287–292. [Google Scholar] [CrossRef]
- Joo, H.M.; Hyun, Y.J.; Myoung, K.S.; Ahn, Y.T.; Lee, J.H.; Huh, C.S.; Han, M.J.; Kim, D.H. Lactobacillus johnsonii HY7042 ameliorates Gardnerella vaginalis-induced vaginosis by killing Gardnerella vaginalis and inhibiting NF-κB activation. Int. Immunopharmacol. 2011, 11, 1758–1765. [Google Scholar] [CrossRef]
- Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.D.L.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; EFSA Panel on Additives Products or Substances used in Animal Feed (FEEDAP); et al. Guidance on the characterisation of microorganisms used as feed additives or as production organisms. EFSA J. 2018, 16, e05206. [Google Scholar] [PubMed]
- Shehata, M.; El Sohaimy, S.; El-Sahn, M.A.; Youssef, M. Screening of isolated potential probiotic lactic acid bacteria for cholesterol lowering property and bile salt hydrolase activity. Ann. Agric. Sci. 2016, 61, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Dashnyam, P.; Mudududdla, R.; Hsieh, T.J.; Lin, T.C.; Lin, H.Y.; Chen, P.Y.; Hsu, C.Y.; Lin, C.H. β-Glucuronidases of opportunistic bacteria are the major contributors to xenobiotic-induced toxicity in the gut. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Z.; Luna, Y.; Yu, P.; Fan, H. Lactobacilli inactivate Chlamydia trachomatis through lactic acid but not H2O2. PLoS ONE 2014, 9, e107758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Hanlon, D.E.; Moench, T.R.; Cone, R.A. In vaginal fluid, bacteria associated with bacterial vaginosis can be suppressed with lactic acid but not hydrogen peroxide. BMC Infect. Dis. 2011, 11, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Tachedjian, G.; Aldunate, M.; Bradshaw, C.S.; Cone, R.A. The role of lactic acid production by probiotic Lactobacillus species in vaginal health. Res. J. Microbiol. 2017, 168, 782–792. [Google Scholar] [CrossRef] [PubMed]
- Hemalatha, R.; Ramalaxmi, B.A.; Swetha, E.; Balakrishna, N.; Mastromarino, P. Evaluation of vaginal pH for detection of bacterial vaginosis. Indian J. Med. Res. 2013, 138, 354. [Google Scholar]
- Aroutcheva, A.; Gariti, D.; Simon, M.; Shott, S.; Faro, J.; Simoes, J.A.; Gurguis, A.; Faro, S. Defense factors of vaginal lactobacilli. Am. J. Obstet. Gynecol. 2001, 185, 375–379. [Google Scholar] [CrossRef]
- Boskey, E.; Cone, R.; Whaley, K.; Moench, T. Origins of vaginal acidity: High D/L lactate ratio is consistent with bacteria being the primary source. Hum. Reprod. 2001, 16, 809–1813. [Google Scholar] [CrossRef]
- Giraud, E.; Lelong, B.; Raimbault, M. Influence of pH and initial lactate concentration on the growth of Lactobacillus plantarum. Appl. Microbiol. Biotechnol. 1991, 36, 96–99. [Google Scholar] [CrossRef]
- Witkin, S.S.; Mendes-Soares, H.; Linhares, I.M.; Jayaram, A.; Ledger, W.J.; Forney, L.J. Influence of vaginal bacteria and D-and L-lactic acid isomers on vaginal extracellular matrix metalloproteinase inducer: Implications for protection against upper genital tract infections. MBio 2013, 4, e00413–e00460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borgdorff, H.; Tsivtsivadze, E.; Verhelst, R.; Marzorati, M.; Jurriaans, S.; Ndayisaba, G.F.; Schuren, F.H.; van de Wijgert, J.H. Lactobacillus-dominated cervicovaginal microbiota associated with reduced HIV/STI prevalence and genital HIV viral load in African women. ISME J. 2014, 8, 1781–1793. [Google Scholar] [CrossRef] [PubMed]
- Kindinger, L.M.; Bennett, P.R.; Lee, Y.S.; Marchesi, J.R.; Smith, A.; Cacciatore, S.; Holmes, E.; Nicholson, J.K.; Teoh, T.G.; MacIntyre, D.A. The interaction between vaginal microbiota, cervical length, and vaginal progesterone treatment for preterm birth risk. Microbiome 2017, 5, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Dronkers, T.; Ouwehand, A.C.; Rijkers, G. Global analysis of clinical trials with probiotics. Heliyon 2020, 6, e04467. [Google Scholar] [CrossRef]
- García-Cayuela, T.; Korany, A.M.; Bustos, I.; de Cadiñanos, L.P.G.; Requena, T.; Peláez, C.; Martínez-Cuesta, M.C. Adhesion abilities of dairy Lactobacillus plantarum strains showing an aggregation phenotype. Food Res. Int. 2014, 57, 44–50. [Google Scholar] [CrossRef]
- Morelli, L. In vitro assessment of probiotic bacteria: From survival to functionality. Int. Dairy J. 2007, 17, 1278–1283. [Google Scholar] [CrossRef]
- Ross, R.; Desmond, C.; Fitzgerald, G.; Stanton, C. Overcoming the technological hurdles in the development of probiotic foods. J. Appl. Microbiol. 2005, 98, 1410–1417. [Google Scholar] [CrossRef]
- Kang, C.H.; Kim, Y.G.; Han, S.H.; Jeong, Y.; Paek, N.S. Antibacterial activity and probiotic properties of lactic acid bacteria from Korean Intestine Origin. Korean Soc. Biotechnol. Bioeng. J. 2017, 32, 153–159. [Google Scholar]
- Patterson, J.L.; Stull-Lane, A.; Girerd, P.H.; Jefferson, K.K. Analysis of adherence, biofilm formation and cytotoxicity suggests a greater virulence potential of Gardnerella vaginalis relative to other bacterial-vaginosis-associated anaerobes. Microbiology 2010, 156, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swidsinski, A.; Mendling, W.; Loening-Baucke, V.; Swidsinski, S.; Dörffel, Y.; Scholze, J.; Lochs, H.; Verstraelen, H. An adherent Gardnerella vaginalis biofilm persists on the vaginal epithelium after standard therapy with oral metronidazole. Am. J. Obstet. Gynecol. 2008, 198, e1–e97. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, A.; Mendling, W.; Loening-Baucke, V.; Ladhoff, A.; Swidsinski, S.; Hale, L.P.; Lochs, H. Adherent biofilms in bacterial vaginosis. Obstet. Gynecol. 2005, 106, 1013–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabbatini, S.; Monari, C.; Ballet, N.; Decherf, A.C.; Bozza, S.; Camilloni, B.; Perito, S.; Vecchiarelli, A. Anti-Biofilm Properties of Saccharomyces cerevisiae CNCM I-3856 and Lacticaseibacillus rhamnosus ATCC 53103 Probiotics against G. vaginalis. Microorganisms 2020, 8, 1294. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, V.; Spano, G. Horizontal gene transfer in the gut: Is it a risk? Food Res. Int. 2009, 42, 1501–1502. [Google Scholar] [CrossRef]
- Charteris, W.P.; Kelly, P.M.; Morelli, L.; Collins, J.K. Gradient diffusion antibiotic susceptibility testing of potentially probiotic lactobacilli. J. Food Prot. 2001, 64, 2007–2014. [Google Scholar] [CrossRef]
- Imperial, I.C.; Ibana, J.A. Addressing the antibiotic resistance problem with probiotics: Reducing the risk of its double-edged sword effect. Front. Microbiol. 2016, 7, 1983. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Exposure Time (log CFU/mL) | |||
|---|---|---|---|---|
| T0 | T12 | T24 | T36 | |
| Control (mBHI with 10% MRS broth) | 6.93 ± 0.014 | 7.69 ± 0.004 a | 8.07 ± 0.015 a | 6.56 ± 0.021 a |
| Lig. salivarius MG242 | 7.28 ± 0.013 d (61.72%) | 7.90 ± 0.028 c (33.05%) | 6.17 ± 0.012 c (59.78%) | |
| M2 (MG901 + MG989) | 7.39 ± 0.014 c (49.90%) | 7.97 ± 0.028 b (21.19%) | 6.29 ± 0.009 b (16.10%) | |
| M3 (MG242 + MG901 + MG989) | 7.54 ± 0.010 b (30.51%) | 7.94 ± 0.003 bc (26.69%) | 6.12 ± 0.043 c (64.16%) | |
| Antibiotics (μL/mL) | Lig.salivarius MG242 | Lim. fermentum MG901 | Lac. plantarum MG989 | |||
|---|---|---|---|---|---|---|
| MIC | EFSA | MIC | EFSA | MIC | EFSA | |
| Ampicillin | 0.094 | 4 | 0.038 | 2 | 0.75 | 2 |
| Gentamycin | 2 | 16 | 0.125 | 16 | 0.094 | 16 |
| Kanamycin | 64 | 64 | 2 | 64 | 24 | 64 |
| Streptomycin | 24 | 64 | 1.5 | 64 | n.r. | n.r. |
| Tetracycline | 0.75 | 8 | 6 | 8 | 32 | 32 |
| Chloramphenicol | 4 | 4 | 12 | 4 | 12 | 8 |
| Erythromycin | 0.047 | 1 | 0.023 | 1 | 0.19 | 1 |
| Clindamycin | 0.064 | 4 | <0.016 | 4 | 0.094 | 4 |
| Enzyme | Lig. Salivarius MG242 | Lim. Fermentum MG901 | Lac. Plantarum MG989 |
|---|---|---|---|
| Alkaline phosphatase | 1 | 0 | 1 |
| Esterase (C4) | 1 | 3 | 1 |
| Esterase lipase (C8) | 1 | 2 | 1 |
| Leucine arylamidase | 4 | 3 | 4 |
| Valine arylamidase | 1 | 1 | 3 |
| Cystine arylamidase | 1 | 1 | 1 |
| Acid phosphatase | 3 | 1 | 2 |
| Naphthol-AS-BI-phosphohydrolase | 3 | 2 | 2 |
| α-Galactosidase | 1 | 4 | 0 |
| β-Galactosidase | 0 | 5 | 5 |
| α-Glucosidase | 0 | 3 | 3 |
| β-Glucosidase | 0 | 0 | 4 |
| N-Acetyl-β-glucosaminidase | 0 | 0 | 5 |
| Strains | Lactic Acid Content (g/L) | Isomer Ratio (%) | |||
|---|---|---|---|---|---|
| D (-) | L (+) | Total (D + L) | D (-) | L (+) | |
| Lig. salivarius MG242 | 1.1 | 16.4 | 17.4 | 6.3 | 94.3 |
| Lim. fermentum MG901 | 4.8 | 7.0 | 11.8 | 40.7 | 59.3 |
| Lac. plantarum MG989 | 0.9 | 10.9 | 11.8 | 7.6 | 92.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Kim, Y.; Kang, C.-H. In Vivo Confirmation of the Antimicrobial Effect of Probiotic Candidates against Gardnerella vaginalis. Microorganisms 2021, 9, 1690. https://doi.org/10.3390/microorganisms9081690
Kim H, Kim Y, Kang C-H. In Vivo Confirmation of the Antimicrobial Effect of Probiotic Candidates against Gardnerella vaginalis. Microorganisms. 2021; 9(8):1690. https://doi.org/10.3390/microorganisms9081690
Chicago/Turabian StyleKim, Hyemin, YongGyeong Kim, and Chang-Ho Kang. 2021. "In Vivo Confirmation of the Antimicrobial Effect of Probiotic Candidates against Gardnerella vaginalis" Microorganisms 9, no. 8: 1690. https://doi.org/10.3390/microorganisms9081690
APA StyleKim, H., Kim, Y., & Kang, C. -H. (2021). In Vivo Confirmation of the Antimicrobial Effect of Probiotic Candidates against Gardnerella vaginalis. Microorganisms, 9(8), 1690. https://doi.org/10.3390/microorganisms9081690