
Lactic Starter Dose Shapes S. aureus and STEC O26:H11 Growth, and Bacterial Community Patterns in Raw Milk Uncooked Pressed Cheeses

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Experimental Design
2.3. Cheese-Making and Sampling
2.4. Physico-Chemical Analyses
2.5. Microbiological Analyses
2.6. Amplicon Sequencing
2.7. Statistical Analyses
3. Results
3.1. Quality of Raw Milks
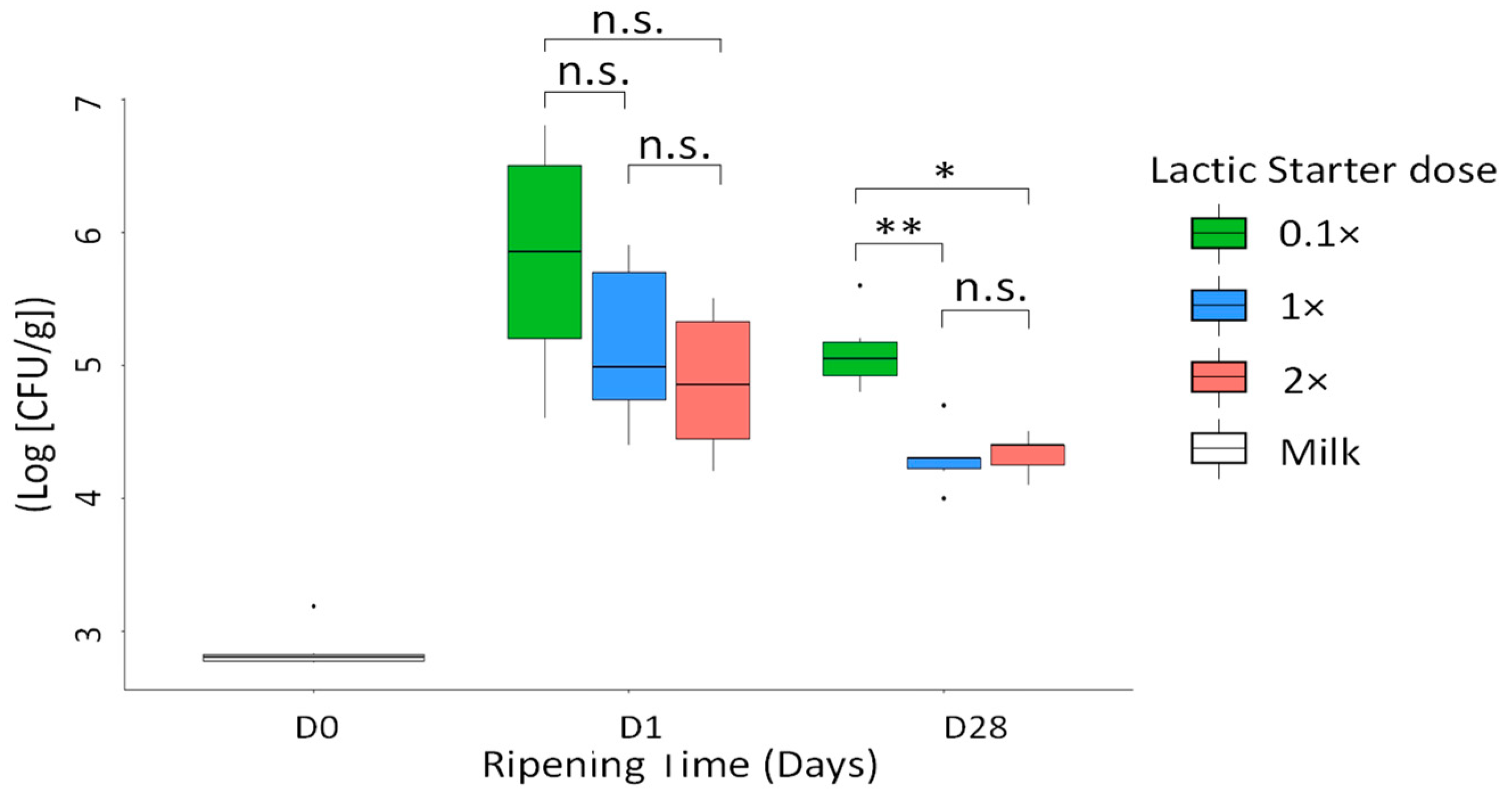
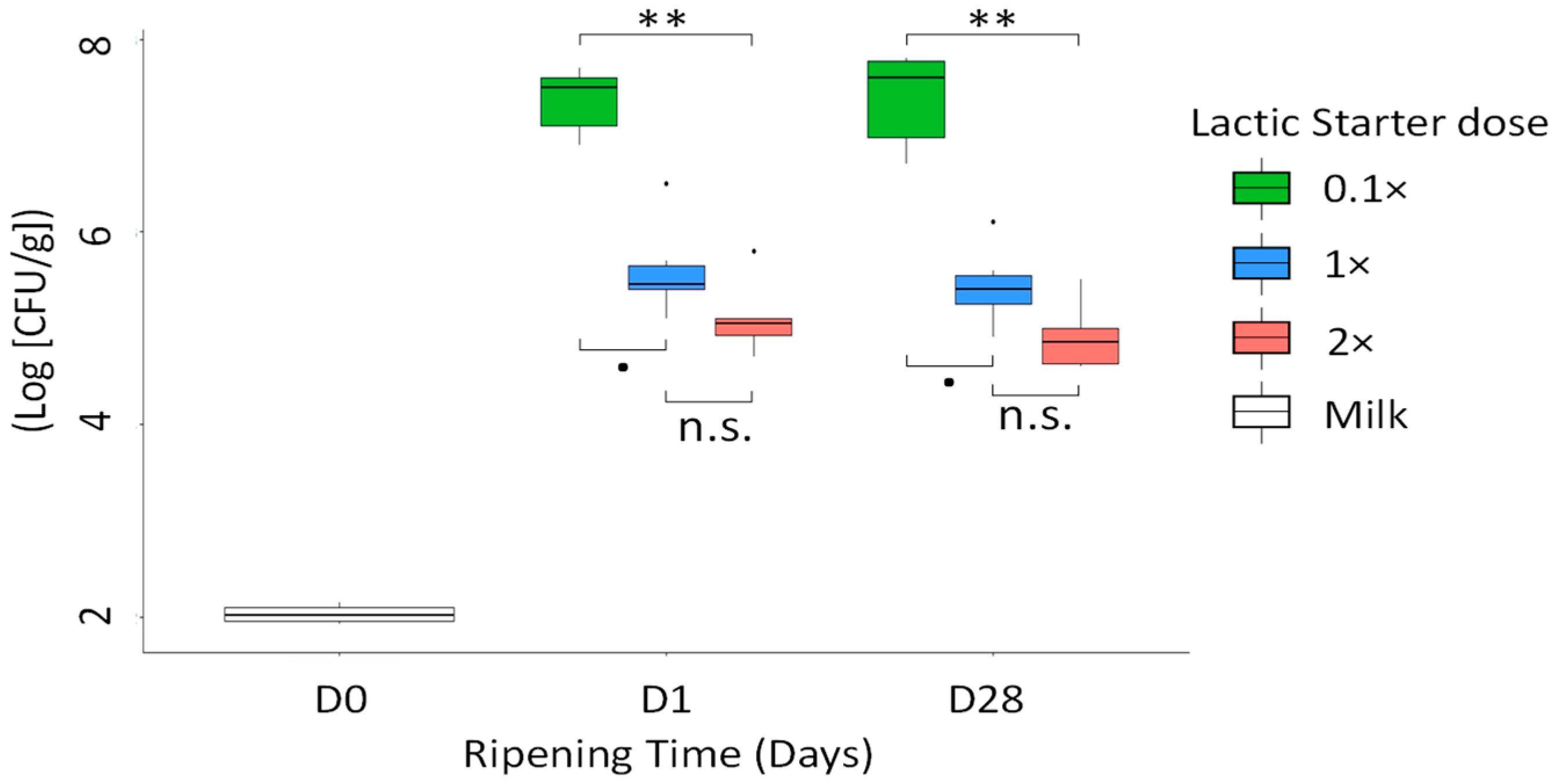
3.2. Levels of Pathogens
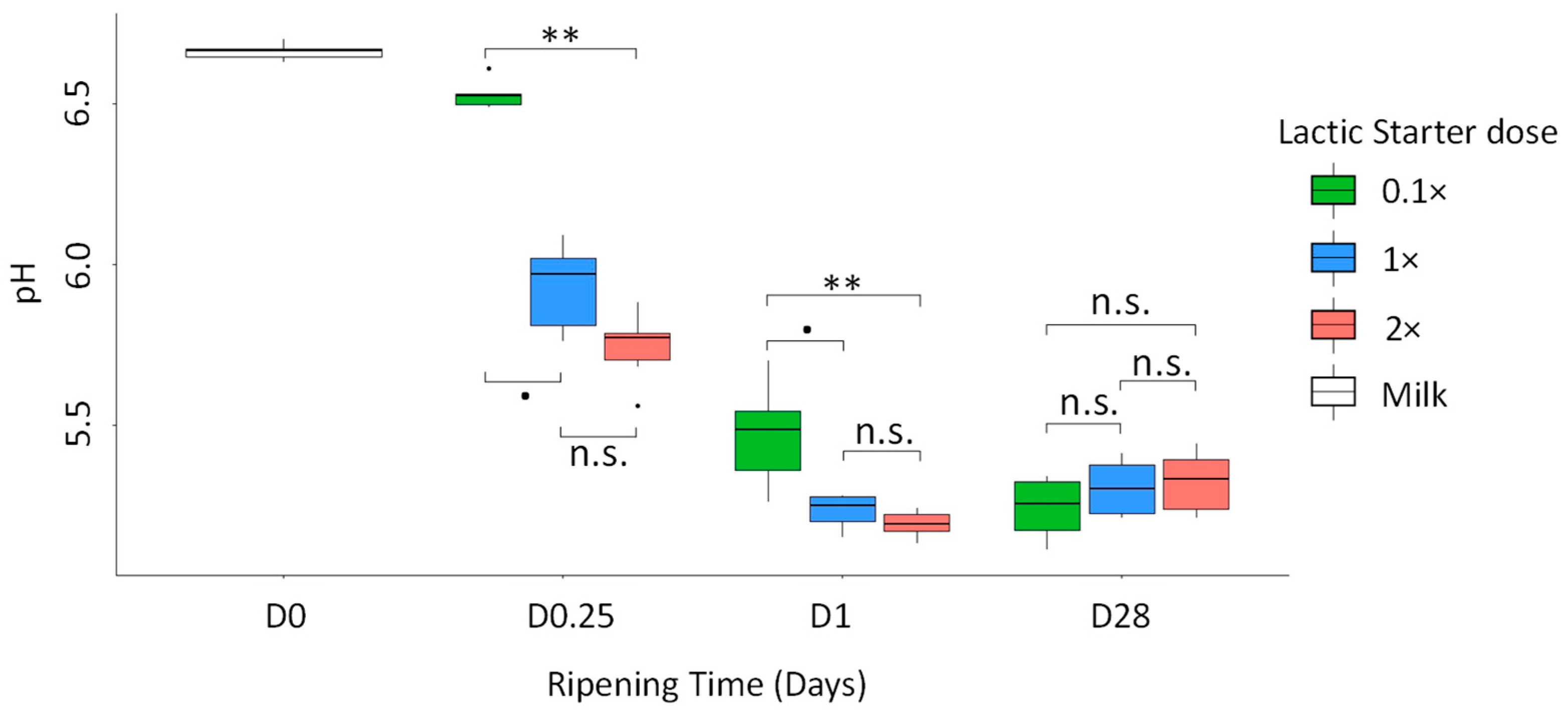
3.3. Physico-Chemical Parameters
3.4. Microbial Counts
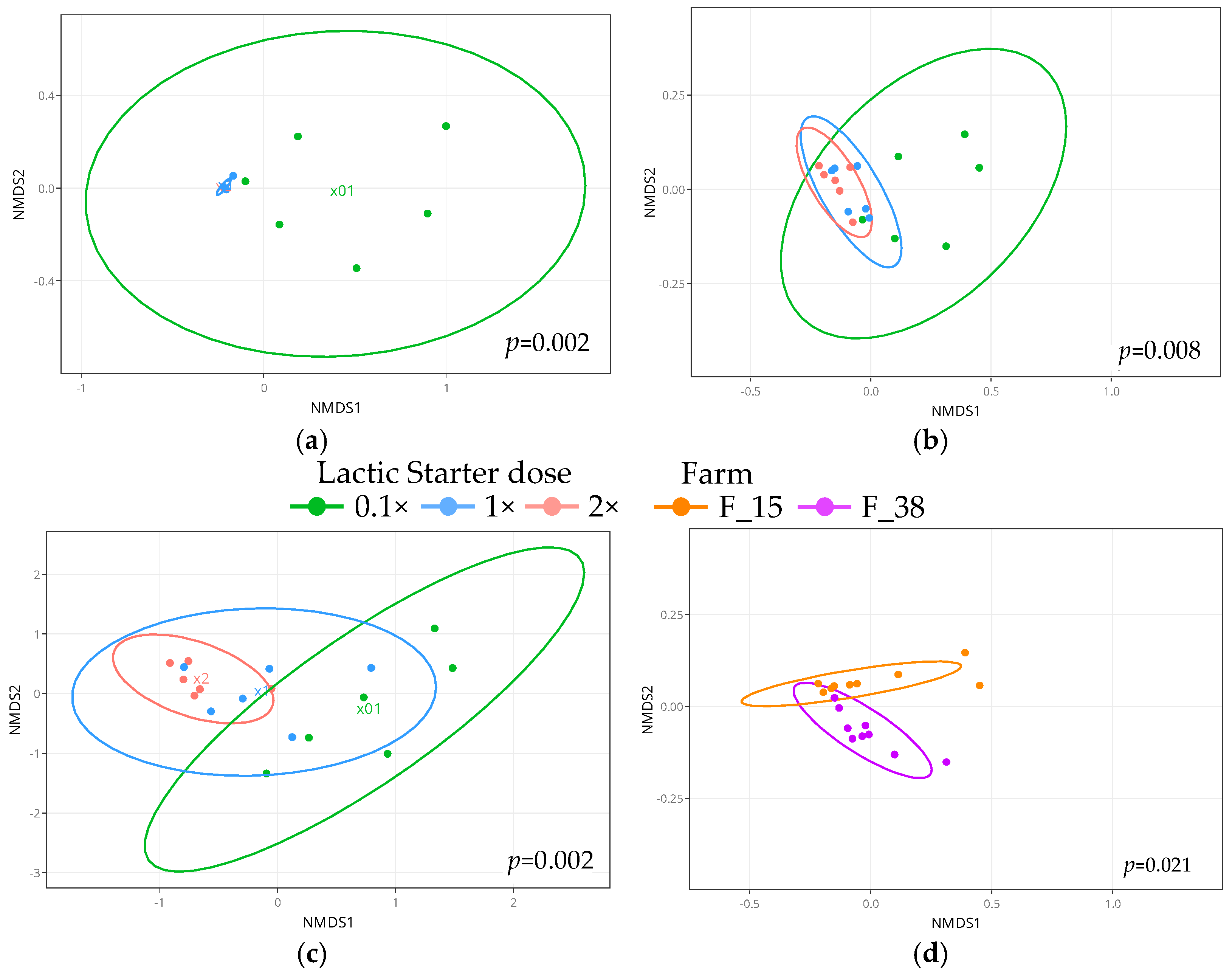
3.5. Bacterial Diversity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Settanni, L.; Moschetti, G. New trends in technology and identity of traditional dairy and fermented meat production processes: Preservation of typicality and hygiene. Trends Food Sci. Technol. 2014, 37, 51–58. [Google Scholar] [CrossRef]
- Zindy, P.; Halawany-Darson, R.; Hauwuy, A. Consumers’ perception and attitude towards regional “terroir” products: Focus on “terroir” cheeses. INRA Prod. Anim. 2017, 12, 229–240. [Google Scholar]
- Blaya, J.; Barzideh, Z.; LaPointe, G. Symposium review: Interaction of starter cultures and nonstarter lactic acid bacteria in the cheese environment. J. Dairy Sci. 2018, 101, 3611–3629. [Google Scholar] [CrossRef] [PubMed]
- Callon, C.; Berdagué, J.L.; Dufour, E.; Montel, M.C. The effect of raw milk microbial flora on the sensory characteristics of salers-type cheeses. J. Dairy Sci. 2005, 88, 3840–3850. [Google Scholar] [CrossRef] [Green Version]
- Gobbetti, M.; Di Cagno, R.; Calasso, M.; Neviani, E.; Fox, P.F.; De Angelis, M. Drivers that establish and assembly the lactic acid bacteria biota in cheeses. Trends Food Sci. Technol. 2018, 78, 244–254. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Mancini, L.; Fox, P.F. Pros and cons for using Non-Starter Lactic Acid Bacteria (NSLAB) as secondary/adjunct starters for cheese ripening. Trends Food Sci. Technol. 2015, 45, 167–178. [Google Scholar] [CrossRef]
- Kongo, J.M. Lactic acid bacteria as starter-cultures for cheese processing: Past, present and future developments. In Lactic Acid Bacteria—R & D for Food, Health and Livestock Purposes; InTech Open Limited: London, UK, 2013. [Google Scholar] [CrossRef] [Green Version]
- Frétin, M.; Martin, B.; Rifa, E.; Isabelle, V.-M.; Pomiès, D.; Ferlay, A.; Montel, M.-C.; Delbès, C. Bacterial community assembly from cow teat skin to ripened cheeses is influenced by grazing systems. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montel, M.-C.; Buchin, S.; Mallet, A.; Delbes-Paus, C.; Vuitton, D.A.; Desmasures, N.; Berthier, F. Traditional cheeses: Rich and diverse microbiota with associated benefits. Int. J. Food Microbiol. 2014, 177, 136–154. [Google Scholar] [CrossRef]
- Johnson, M.E. A 100-year review: Cheese production and quality. J. Dairy Sci. 2017, 100, 9952–9965. [Google Scholar] [CrossRef]
- O’Sullivan, D.J.; Giblin, L.; McSweeney, P.L.H.; Sheehan, J.J.; Cotter, P.D. Nucleic acid-based approaches to investigate microbial-related cheese quality defects. Front. Microbiol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Donnelly, C. Review of Controls for Pathogen Risks in Scottish Artisan Cheeses Made from Unpasteurised Milk; Food Standards Scotland: Aberdeen, UK, 2018.
- Fournet, N.; Laurent, E.; Jones, G.; Jourdan Da Silva, N.; Tourdjman, M.; Chereau, F.; Nisavanh, A.; De Valk, H. Surveillance of Collective Food Toxi-Infections. Data from the Mandatory Declaration 2018; Santé Publique France: Paris, France, 2020.
- Bonanno, L.; Delubac, B.; Michel, V.; Auvray, F. Influence of stress factors related to cheese-making process and to STEC detection procedure on the induction of stx phages from STEC O26:H11. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Delbes, C.; Alomar, J.; Chougui, N.; Martin, J.-F.; Montel, M.-C. Staphylococcus aureus Growth and enterotoxin production during the manufacture of uncooked, semihard cheese from cows’ raw milk. J. Food Prot. 2006, 69, 2161–2167. [Google Scholar] [CrossRef] [PubMed]
- Miszczycha, S.D.; Perrin, F.; Ganet, S.; Jamet, E.; Tenenhaus-Aziza, F.; Montel, M.-C.; Thevenot-Sergentet, D. Behavior of different shiga toxin-producing Escherichia coli serotypes in various experimentally contaminated raw-milk cheeses. Appl. Environ. Microbiol. 2013, 79, 150–158. [Google Scholar] [CrossRef] [Green Version]
- Parente, E.; Guidone, A.; Matera, A.; De Filippis, F.; Mauriello, G.; Ricciardi, A. Microbial community dynamics in thermophilic undefined milk starter cultures. Int. J. Food Microbiol. 2016, 217, 59–67. [Google Scholar] [CrossRef]
- Lucchini, R.; Cardazzo, B.; Carraro, L.; Negrinotti, M.; Balzan, S.; Novelli, E.; Fasolato, L.; Fasoli, F.; Farina, G. Contribution of natural milk culture to microbiota, safety and hygiene of raw milk cheese produced in alpine malga. Ital. J. Food Saf. 2018, 7. [Google Scholar] [CrossRef]
- International Standard Organisation (ISO). ISO 6888-2:1999(en) Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Coagulase-Positive Staphylococci (Staphylococcus aureus and Other Species)—Part 2: Technique Using Rabbit Plasma Fibrinogen Agar Medium; CEN Comite Europeen de Normalisation: Brussels, Belgium, 1999; Volume 7. [Google Scholar]
- International Standard Organisation (ISO). ISO/TS 13136:2012(en) Microbiology of Food and Animal Feed—Real-Time Polymerase Chain Reaction (PCR)-Based Method for the Detection of Food-Borne Pathogens—Horizontal Method for the Detection of Shiga Toxin-Producing Escherichia coli (STEC) and the Determination of O157, O111, O26, O103 and O145 Serogroups; CEN Comite Europeen de Normalisation: Brussels, Belgium, 2012; Volume 22. [Google Scholar]
- Mitra, S. Multiple data analyses and statistical approaches for analyzing data from metagenomic studies and clinical trials. In Evolutionary Genomics: Statistical and Computational Methods; Methods in Molecular Biology; Anisimova, M., Ed.; Springer: New York, NY, USA, 2019; pp. 605–634. ISBN 978-1-4939-9074-0. [Google Scholar]
- Quigley, L.; O’Sullivan, O.; Beresford, T.P.; Ross, R.P.; Fitzgerald, G.F.; Cotter, P.D. High-Throughput sequencing for detection of subpopulations of bacteria not previously associated with artisanal cheeses. Appl. Environ. Microbiol. 2012, 78, 5717–5723. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, D.J.; Cotter, P.D.; O’Sullivan, O.; Giblin, L.; McSweeney, P.L.H.; Sheehan, J.J. Temporal and spatial differences in microbial composition during the manufacture of a continental-type cheese. Appl. Environ. Microbiol. 2015, 81, 2525–2533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carafa, I.; Stocco, G.; Franceschi, P.; Summer, A.; Tuohy, K.M.; Bittante, G.; Franciosi, E. Evaluation of autochthonous lactic acid bacteria as starter and non-starter cultures for the production of traditional mountain cheese. Food Res. Int. 2019, 115, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Feutry, F.; Oneca, M.; Berthier, F.; Torre, P. Biodiversity and growth dynamics of lactic acid bacteria in artisanal PDO ossau-iraty cheeses made from raw ewe’s milk with different starters. Food Microbiol. 2012, 29, 33–42. [Google Scholar] [CrossRef]
- Settanni, L.; Di Grigoli, A.; Tornambé, G.; Bellina, V.; Francesca, N.; Moschetti, G.; Bonanno, A. Persistence of wild Streptococcus thermophilus strains on wooden vat and during the manufacture of a traditional caciocavallo type cheese. Int. J. Food Microbiol. 2012, 155, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Kamimura, B.A.; Cabral, L.; Noronha, M.F.; Baptista, R.C.; Nascimento, H.M.; Sant’Ana, A.S. Amplicon sequencing reveals the bacterial diversity in milk, dairy premises and Serra Da Canastra artisanal cheeses produced by three different farms. Food Microbiol. 2020, 89, 103453. [Google Scholar] [CrossRef]
- Choi, J.; In Lee, S.; Rackerby, B.; Frojen, R.; Goddik, L.; Ha, S.-D.; Park, S.H. Assessment of overall microbial community shift during cheddar cheese production from raw milk to aging. Appl. Microbiol. Biotechnol. 2020, 104, 6249–6260. [Google Scholar] [CrossRef]
- Fernandez-García, E.; Tomillo, J.; Nuñez, M. Formation of biogenic amines in raw milk Hispánico cheese manufactured with proteinases and different levels of starter culture. J. Food Prot. 2000, 63, 1551–1555. [Google Scholar] [CrossRef] [PubMed]
- Khosrowshahi, A.; Madadlou, A.; Ebrahim Zadeh Mousavi, M.; Emam-Djomeh, Z. Monitoring the chemical and textural changes during ripening of Iranian white cheese made with different concentrations of starter. J. Dairy Sci. 2006, 89, 3318–3325. [Google Scholar] [CrossRef]
- Özer, E.; Kesenkaş, H. The effect of using different starter culture combinations on ripening parameters, microbiological and sensory properties of Mihaliç cheese. J. Food Sci. Technol. 2019, 56, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- De Buyser, M.L.; Lombard, B.; Schulten, S.M.; In’t Veld, P.H.; Scotter, S.L.; Rollier, P.; Lahellec, C. Validation of EN ISO standard methods 6888 Part 1 and Part 2: 1999—Enumeration of coagulase-positive staphylococci in foods. Int. J. Food Microbiol. 2003, 83, 185–194. [Google Scholar] [CrossRef]
- International Standard Organisation (ISO). ISO 19020:2017(en) Microbiology of the Food Chain—Horizontal Method for the Immunoenzymatic Detection of Staphylococcal Enterotoxins in Foodstuffs; CEN Comite Europeen de Normalisation: Brussels, Belgium, 2017; Volume 21. [Google Scholar]
- Frétin, M.; Chassard, C.; Delbès, C.; Lavigne, R.; Rifa, E.; Theil, S.; Fernandez, B.; Laforce, P.; Callon, C. Robustness and efficacy of an inhibitory consortium against E. coli O26:H11 in raw milk cheeses. Food Control 2020, 115, 107282. [Google Scholar] [CrossRef]
- Millet, L.; Saubusse, M.; Didienne, R.; Tessier, L.; Montel, M.C. Control of Listeria monocytogenes in raw-milk cheeses. Int. J. Food Microbiol. 2006, 108, 105–114. [Google Scholar] [CrossRef] [PubMed]
- International Standard Organisation (ISO). ISO 5534:2004 (en) (IDF 4:2004) Cheese and Processed Cheese–Determination of the Total Solids Content (Reference Method); CEN Comite Europeen de Normalisation: Brussels, Belgium, 2004; Volume 7. [Google Scholar]
- Frétin, M.; Ferlay, A.; Verdier-Metz, I.; Fournier, F.; Montel, M.-C.; Farruggia, A.; Delbès, C.; Martin, B. The effects of low-input grazing systems and milk pasteurisation on the chemical composition, microbial communities, and sensory properties of uncooked pressed cheeses. Int. Dairy J. 2017, 64, 56–67. [Google Scholar] [CrossRef]
- Noël, Y.; Ardö, Y.; Pocher, S.; Hunter, A.; Lavanchy, P.; Luginbuhl, W.; Bars, D.L.; Polychroniadou, A.; Pellegrino, L. Characterisation of protected denomination of origin cheeses: Relationships between sensory texture and instrumental data. Lait 1998, 78, 569–588. [Google Scholar] [CrossRef]
- Terzaghi, B.E.; Sandine, W.E. Improved medium for lactic Streptococci and their bacteriophages. Appl. Microbiol. 1975, 29, 807–813. [Google Scholar] [CrossRef]
- Man, J.C.D.; Rogosa, M.; Sharpe, M.E. A medium for the cultivation of lactobacilli. J. Appl. Bacteriol. 1960, 23, 130–135. [Google Scholar] [CrossRef]
- Denis, C.; Gueguen, M.; Henry, E.; Levert, D. New media for the numeration of cheese surface bacteria. Lait 2001, 81, 365–379. [Google Scholar] [CrossRef]
- Mossel, D.A.; Kleynen-Semmeling, A.M.; Vincentie, H.M.; Beerens, H.; Catsaras, M. Oxytetracycline-Glucose-Yeast extract agar for selective enumeration of moulds and yeasts in foods and clinical material. J. Appl. Bacteriol. 1970, 33, 454–457. [Google Scholar] [CrossRef] [PubMed]
- Duthoit, F.; Godon, J.-J.; Montel, M.-C. Bacterial community dynamics during production of registered designation of origin salers cheese as evaluated by 16S RRNA gene single-strand conformation polymorphism analysis. Appl. Environ. Microbiol. 2003, 69, 3840–3848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poirier, S.; Rué, O.; Peguilhan, R.; Coeuret, G.; Zagorec, M.; Champomier-Vergès, M.-C.; Loux, V.; Chaillou, S. Deciphering intra-species bacterial diversity of meat and seafood spoilage microbiota using GyrB amplicon sequencing: A comparative analysis with 16S RDNA V3-V4 amplicon sequencing. PLoS ONE 2018, 13, e0204629. [Google Scholar] [CrossRef]
- Theil, S.; Rifa, E. Amplicon workflow for microbial community analysis. F1000Research 2021, 10, 7. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution sample inference from illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Delbès-Paus, C.; Miszczycha, S.; Ganet, S.; Helinck, S.; Veisseire, P.; Pochet, S.; Thévenot, D.; Montel, M.-C. Behavior of Escherichia coli O26:H11 in the Presence of Hafnia alvei in a model cheese ecosystem. Int. J. Food Microbiol. 2013, 160, 212–218. [Google Scholar] [CrossRef]
- Callon, C.; Arliguie, C.; Montel, M.-C. Control of shigatoxin-producing Escherichia coli in cheese by dairy bacterial strains. Food Microbiol. 2016, 53, 63–70. [Google Scholar] [CrossRef]
- Medved’ová, A.; Koňuchová, M.; Kvočiková, K.; Hatalová, I.; Valík, L. Effect of lactic acid bacteria addition on the microbiological safety of pasta-filata types of cheeses. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- European Commission. Commission of the European Communities commission regulation (EC) No 2073/2005 of 15 November 2005 on microbiological criteria for foodstuffs. Off. J. Eur. Union 2005, L338, 1–26. [Google Scholar]
- Alomar, J.; Lebert, A.; Montel, M.C. Effect of temperature and PH on growth of Staphylococcus aureus in co-culture with Lactococcus garvieae. Curr. Microbiol. 2008, 56, 408–412. [Google Scholar] [CrossRef] [PubMed]
- Delpech, P.; Bornes, S.; Alaterre, E.; Bonnet, M.; Gagne, G.; Montel, M.-C.; Delbès, C. Staphylococcus aureus transcriptomic response to inhibition by H2O2-producing Lactococcus garvieae. Food Microbiol. 2015, 51, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Al-Nabulsi, A.A.; Osaili, T.M.; AbuNaser, R.A.; Olaimat, A.N.; Ayyash, M.; Al-Holy, M.A.; Kadora, K.M.; Holley, R.A. Factors affecting the viability of Staphylococcus aureus and production of enterotoxin during processing and storage of white-brined cheese. J. Dairy Sci. 2020, 103, 6869–6881. [Google Scholar] [CrossRef]
- Evivie, S.E.; Ogwu, M.C.; Abdelazez, A.; Bian, X.; Liu, F.; Li, B.; Huo, G. Suppressive effects of Streptococcus thermophilus KLDS 3.1003 on some foodborne pathogens revealed through in vitro, in vivo and genomic insights. Food Funct. 2020, 11, 6573–6587. [Google Scholar] [CrossRef]
- Silvetti, T.; Capra, E.; Morandi, S.; Cremonesi, P.; Decimo, M.; Gavazzi, F.; Giannico, R.; De Noni, I.; Brasca, M. Microbial population profile during ripening of Protected Designation of Origin (PDO) silter cheese, produced with and without autochthonous starter culture. LWT 2017, 84, 821–831. [Google Scholar] [CrossRef]
- Marilley, L.; Casey, M.G. Flavours of cheese products: Metabolic pathways, analytical tools and identification of producing strains. Int. J. Food Microbiol. 2004, 90, 139–159. [Google Scholar] [CrossRef]
- Bukvicki, D.; Siroli, L.; D’Alessandro, M.; Cosentino, S.; Fliss, I.; Said, L.B.; Hassan, H.; Lanciotti, R.; Patrignani, F. Unravelling the potential of Lactococcus lactis strains to be used in cheesemaking production as biocontrol agents. Foods 2020, 9, 1815. [Google Scholar] [CrossRef]
- Anast, J.M.; Dzieciol, M.; Schultz, D.L.; Wagner, M.; Mann, E.; Schmitz-Esser, S. Brevibacterium from Austrian hard cheese harbor a putative histamine catabolism pathway and a plasmid for adaptation to the cheese environment. Sci. Rep. 2019, 9, 6164. [Google Scholar] [CrossRef]
- Bockelmann, W.; Willems, K.P.; Neve, H.; Heller, K.H. Cultures for the ripening of smear cheeses. Int. Dairy J. 2005, 15, 719–732. [Google Scholar] [CrossRef]
- Boyaval, P.; Desmazeaud, M.J. Current state of research on Brevibacterium linens. Lait 1983, 63, 187–216. [Google Scholar] [CrossRef] [Green Version]
- Famelart, M.H.; Graet, Y.L.; Michel, F.; Richoux, R.; Riaublanc, A. Evaluation of the methods of measurement for functional properties of Emmental cheeses from the west of France. Lait 2002, 82, 225–245. [Google Scholar] [CrossRef]
- Saubusse, M.; Millet, L.; Delbès, C.; Callon, C.; Montel, M.C. Application of single strand conformation polymorphism—PCR method for distinguishing cheese bacterial communities that inhibit Listeria monocytogenes. Int. J. Food Microbiol. 2007, 116, 126–135. [Google Scholar] [CrossRef]
- Gay, M.; Amgar, A. Factors moderating Listeria monocytogenes growth in raw milk and in soft cheese made from raw milk. Lait 2005, 85, 153–170. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piqueras, J.; Chassard, C.; Callon, C.; Rifa, E.; Theil, S.; Lebecque, A.; Delbès, C. Lactic Starter Dose Shapes S. aureus and STEC O26:H11 Growth, and Bacterial Community Patterns in Raw Milk Uncooked Pressed Cheeses. Microorganisms 2021, 9, 1081. https://doi.org/10.3390/microorganisms9051081
Piqueras J, Chassard C, Callon C, Rifa E, Theil S, Lebecque A, Delbès C. Lactic Starter Dose Shapes S. aureus and STEC O26:H11 Growth, and Bacterial Community Patterns in Raw Milk Uncooked Pressed Cheeses. Microorganisms. 2021; 9(5):1081. https://doi.org/10.3390/microorganisms9051081
Chicago/Turabian StylePiqueras, Justine, Christophe Chassard, Cécile Callon, Etienne Rifa, Sébastien Theil, Annick Lebecque, and Céline Delbès. 2021. "Lactic Starter Dose Shapes S. aureus and STEC O26:H11 Growth, and Bacterial Community Patterns in Raw Milk Uncooked Pressed Cheeses" Microorganisms 9, no. 5: 1081. https://doi.org/10.3390/microorganisms9051081
APA StylePiqueras, J., Chassard, C., Callon, C., Rifa, E., Theil, S., Lebecque, A., & Delbès, C. (2021). Lactic Starter Dose Shapes S. aureus and STEC O26:H11 Growth, and Bacterial Community Patterns in Raw Milk Uncooked Pressed Cheeses. Microorganisms, 9(5), 1081. https://doi.org/10.3390/microorganisms9051081