
A Markerless Gene Deletion System in Streptococcus suis by Using the Copper-Inducible Vibrio parahaemolyticus YoeB Toxin as a Counterselectable Marker



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, Primers, and Culture Conditions
2.2. Preparationof Synthetic Peptide and Natural Transformation Experiment
2.3. Construction of a S. suis Strain Expressing the Copper-Inducible YoeBVp Toxin
2.4. RNA Extraction
2.5. RT-qPCR Analysis
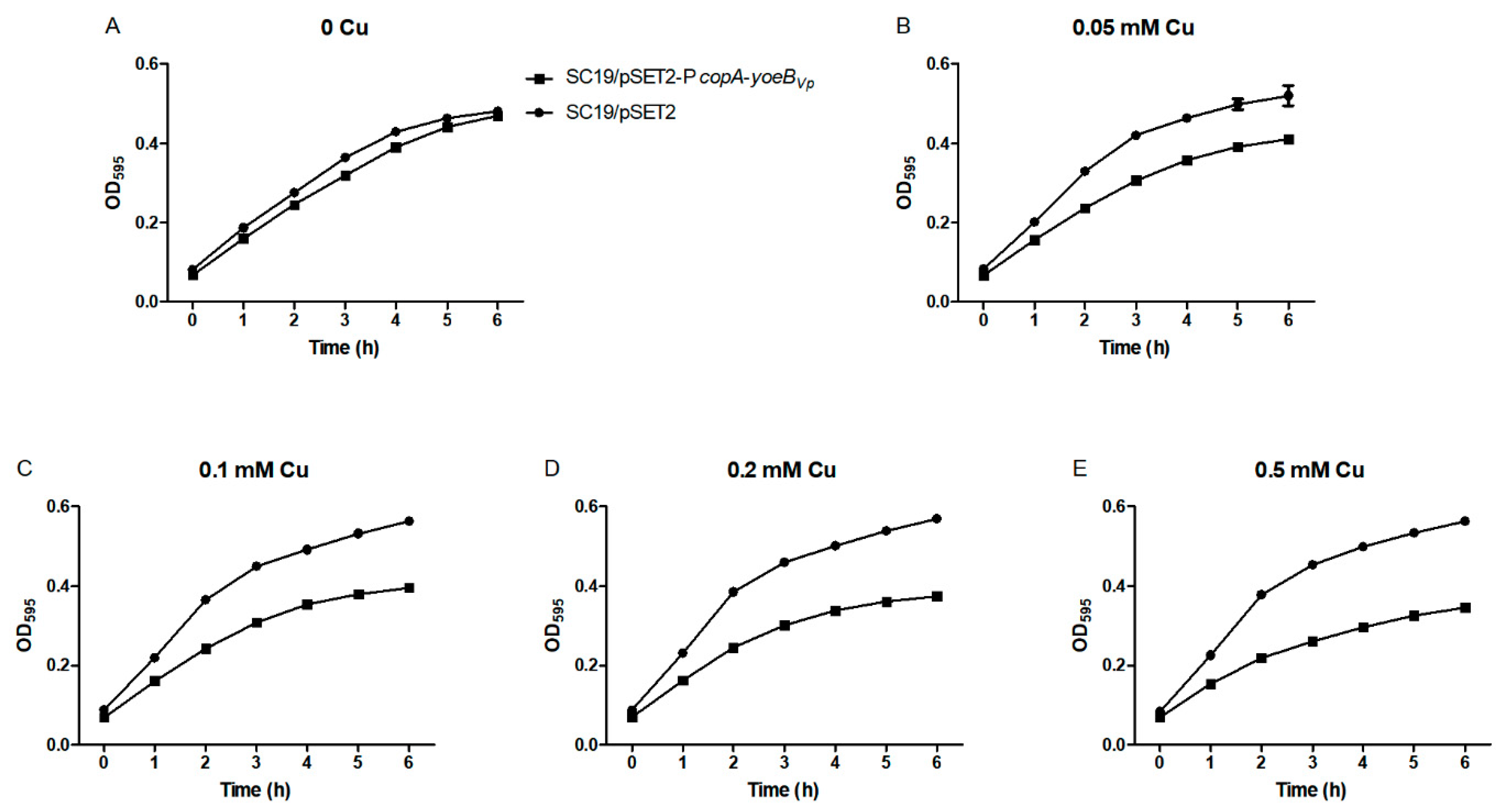
2.6. Growth Curves Analyses
2.7. Spot Dilution Assays
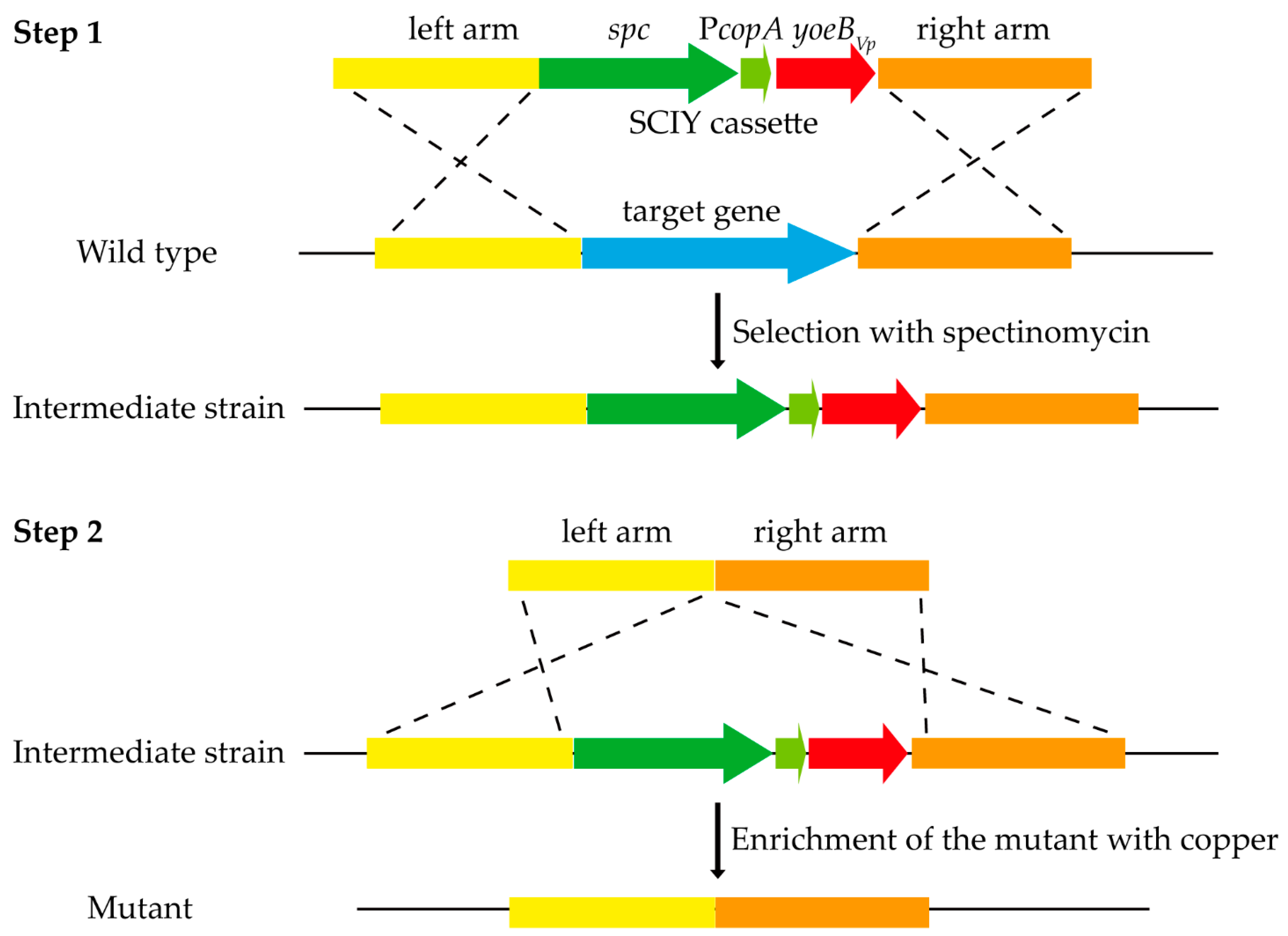
2.8. Construction of the SCIY Positive-Negative Selectable Cassette
2.9. Construction of Markerless Gene Deletion Mutants Using the SCIY Cassette
3. Results
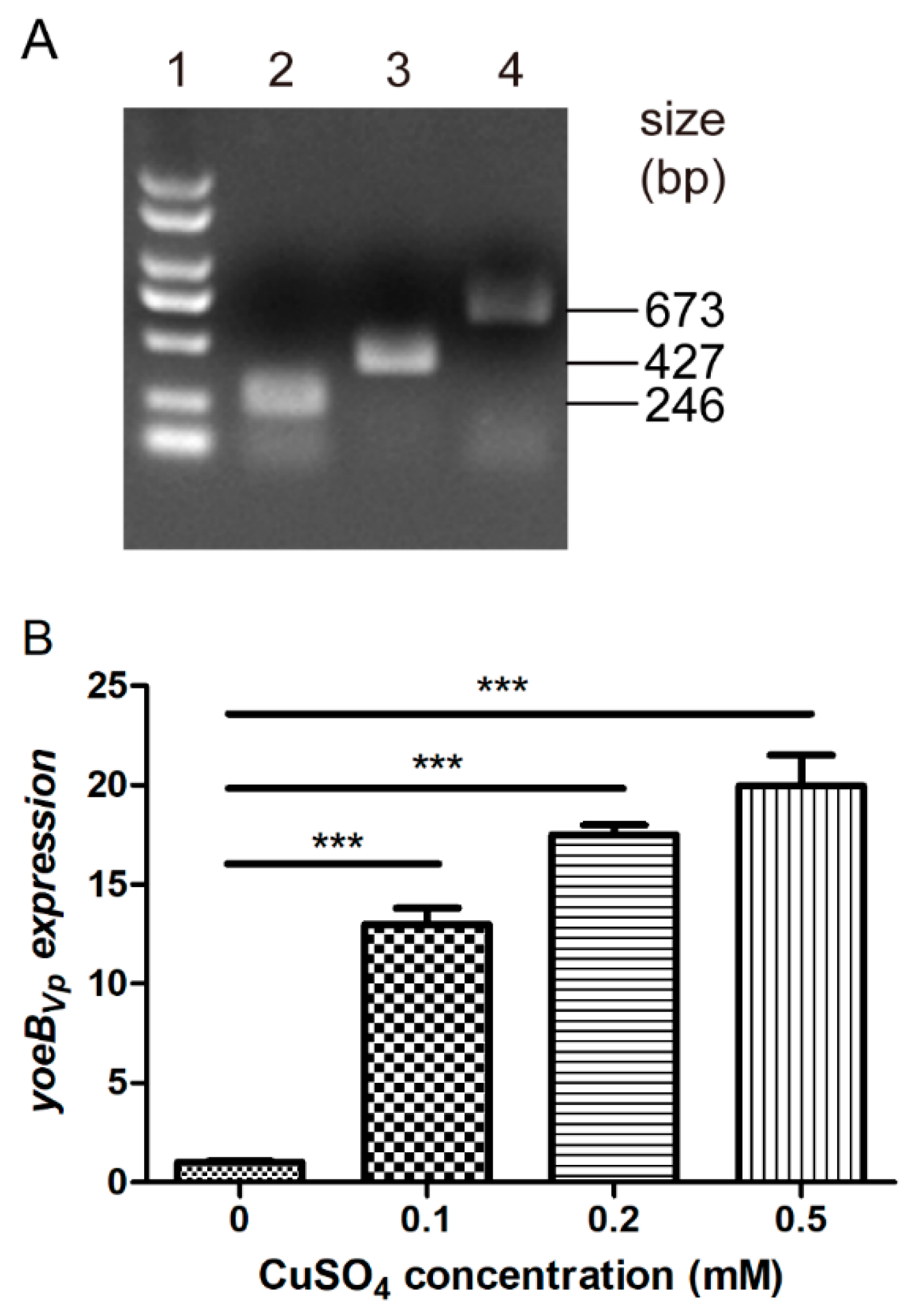
3.1. Identification of the S. suis Strain Expressing the Copper-Inducible YoeBVp Toxin
3.2. YoeBVp Expression Results in Growth Defect in S. suis
3.3. Establishment of the Cloning-Independent and Counterselectable Markerless Gene Deletion System in S. suis
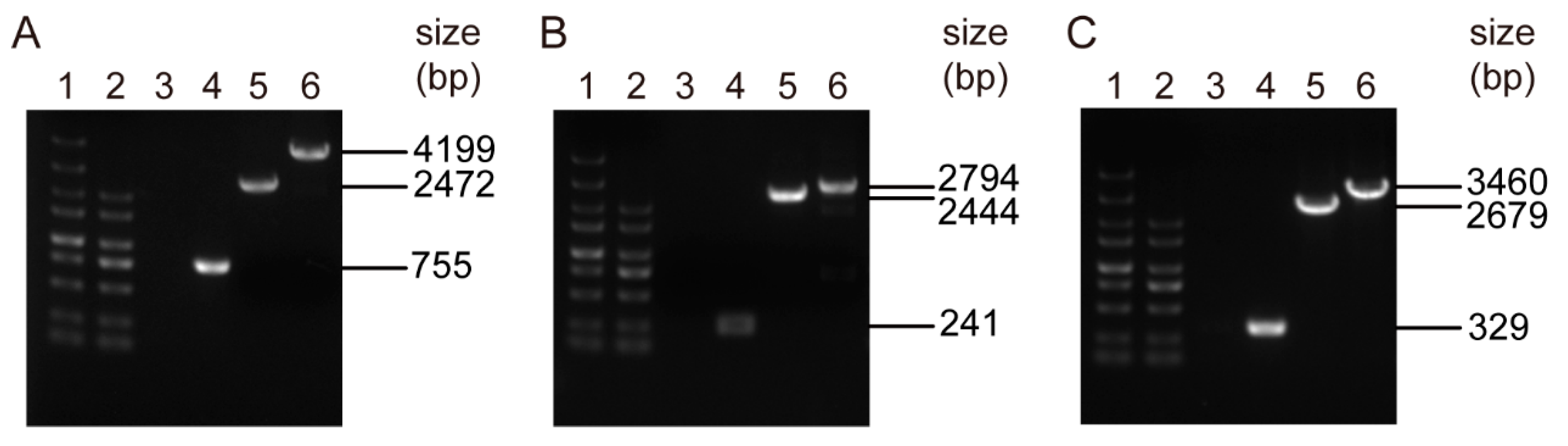
3.4. Markerless Deletion of the pmtA, perR, and lysR Genes in S. suis
3.5. The SCIY Cassette Is Highly Efficient for Counterselection in S. suis
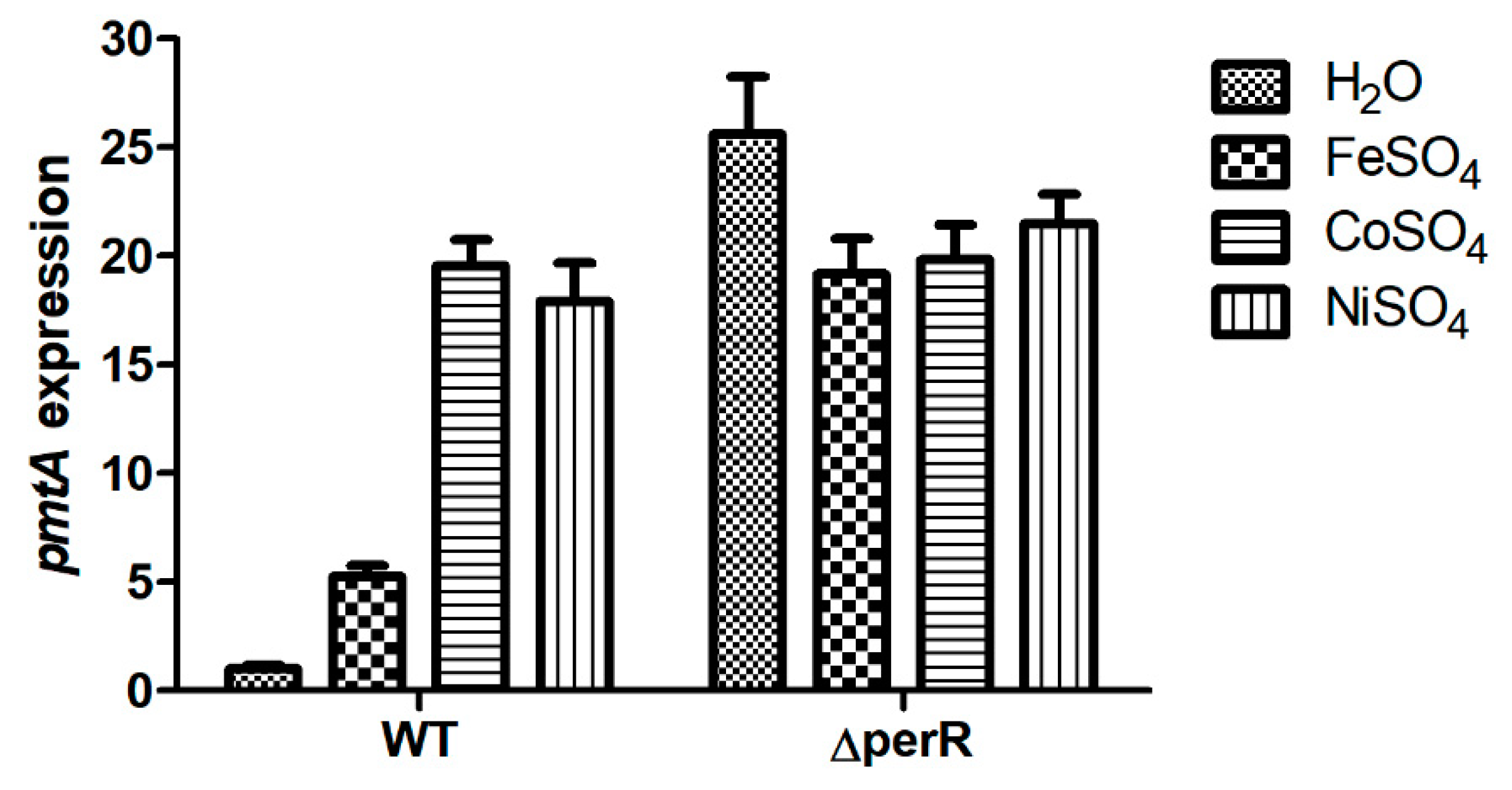
3.6. PerR Is a Transcriptional Repressor of the Ferrous Iron and Cobalt Efflux Pump in S. suis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feng, Y.J.; Zhang, H.M.; Wu, Z.W.; Wang, S.H.; Cao, M.; Hu, D.; Wang, C.J. Streptococcus suis infection An emerging/reemerging challenge of bacterial infectious diseases? Virulence 2014, 5, 477–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lun, Z.R.; Wang, Q.P.; Chen, X.G.; Li, A.X.; Zhu, X.Q. Streptococcus suis: An emerging zoonotic pathogen. Lancet Infect. Dis. 2007, 7, 201–209. [Google Scholar] [CrossRef]
- Segura, M.; Aragon, V.; Brockmeier, S.L.; Gebhart, C.; Greeff, A.; Kerdsin, A.; O’Dea, M.A.; Okura, M.; Salery, M.; Schultsz, C.; et al. Update on Streptococcus suis research and prevention in the era of antimicrobial restriction: 4th International Workshop on S. suis. Pathogens 2020, 9, 374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Ku, X.; Yu, X.; Sun, Q.; Wu, H.; Chen, F.; Zhang, X.; Guo, L.; Tang, X.; He, Q. Prevalence and antimicrobial susceptibilities of bacterial pathogens in Chinese pig farms from 2013 to 2017. Sci. Rep. 2019, 9, 9908. [Google Scholar] [CrossRef]
- Segura, M.; Calzas, C.; Grenier, D.; Gottschalk, M. Initial steps of the pathogenesis of the infection caused by Streptococcus suis: Fighting against nonspecific defenses. FEBS Lett. 2016, 590, 3772–3799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microbes Infect. 2014, 3, e45. [Google Scholar] [CrossRef]
- Tang, J.; Wang, C.; Feng, Y.; Yang, W.; Song, H.; Chen, Z.; Yu, H.; Pan, X.; Zhou, X.; Wang, H.; et al. Streptococcal toxic shock syndrome caused by Streptococcus suis serotype. PLoS Med. 2006, 3, e151. [Google Scholar]
- Yu, H.; Jing, H.; Chen, Z.; Zheng, H.; Zhu, X.; Wang, H.; Wang, S.; Liu, L.; Zu, R.; Luo, L.; et al. Human Streptococcus suis outbreak, Sichuan, China. Emerg. Infect. Dis. 2006, 12, 914–920. [Google Scholar] [CrossRef]
- Romay-Lema, E.M.; Ventura-Valcárcel, P.; Iñiguez-Vázquez, I.; García-Pais, M.J.; García-Garrote, F.; Rabuñal-Rey, R.; Alonso, M.P.; Corredoira-Sánchez, J. Streptococcus suis spondylodiscitis: 2 new cases and a literature review. Enferm. Infecc. Microbiol. Clin. 2020. [Google Scholar] [CrossRef]
- Agoston, Z.; Terhes, G.; Hannauer, P.; Gajdacs, M.; Urban, E. Fatal case of bacteremia caused by Streptococcus suis in a splenectomized man and a review of the European literature. Acta Microbiol. Immunol. Hung. 2020, 67, 148–155. [Google Scholar] [CrossRef] [Green Version]
- Jiang, F.; Guo, J.; Cheng, C.; Gu, B. Human infection caused by Streptococcus suis serotype 2 in China: Report of two cases and epidemic distribution based on sequence type. BMC Infect. Dis. 2020, 20, 223. [Google Scholar] [CrossRef] [Green Version]
- Olearo, F.; Marinosci, A.; Stephan, R.; Cherkaoui, A.; Renzi, G.; Gaia, N.; Leo, S.; Lazarevic, V.; Schrenzel, J. First case of Streptococcus suis infection in Switzerland: An emerging public health problem? Travel Med. Infect. Dis. 2020, 36, 101590. [Google Scholar] [CrossRef]
- Liu, S.S.; Wang, Y.; Xue, L.; Ma, C.; Li, C.H. Hemophagocytic lymphohistiocytosis due to Streptococcus suis in a 12-year-old girl: A case report. Medicine 2019, 98, e15136. [Google Scholar] [CrossRef]
- Németh, A.; Knausz, M.; Schmidt, P. Special case of purulent meningitis caused by Streptococcus suis. Case report. Orv. Hetil. 2019, 160, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Segura, M.; Fittipaldi, N.; Calzas, C.; Gottschalk, M. Critical Streptococcus suis virulence factors: Are they all really critical? Trends Microbiol. 2017, 25, 586–594. [Google Scholar] [CrossRef]
- Fittipaldi, N.; Segura, M.; Grenier, D.; Gottschalk, M. Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol. 2012, 7, 259–279. [Google Scholar] [CrossRef]
- Arenas, J.; Bossers-de Vries, R.; Harders-Westerveen, J.; Buys, H.; Ruuls-van Stalle, L.M.F.; Stockhofe-Zurwieden, N.; Zaccaria, E.; Tommassen, J.; Wells, J.M.; Smith, H.E.; et al. In vivo transcriptomes of Streptococcus suis reveal genes required for niche-specific adaptation and pathogenesis. Virulence 2019, 10, 334–351. [Google Scholar] [CrossRef] [Green Version]
- Arenas, J.; Zomer, A.; Harders-Westerveen, J.; Bootsma, H.J.; Jonge, M.I.D.; Stockhofe-Zurwieden, N.; Smith, H.E.; Greeff, A.D. Identification of conditionally essential genes for Streptococcus suis infection in pigs. Virulence 2020, 11, 446–464. [Google Scholar] [CrossRef]
- Pei, X.; Liu, M.; Zhou, H.; Fan, H. Screening for phagocytosis resistance-related genes via a transposon mutant library of Streptococcus suis serotype. Virulence 2020, 11, 825–838. [Google Scholar] [CrossRef]
- Takamatsu, D.; Osaki, M.; Sekizaki, T. Thermosensitive suicide vectors for gene replacement in Streptococcus suis. Plasmid 2001, 46, 140–148. [Google Scholar] [CrossRef]
- Zhu, Y.; Dong, W.; Ma, J.; Zhang, Y.; Pan, Z.; Yao, H. Utilization of the ComRS system for the rapid markerless deletion of chromosomal genes in Streptococcus suis. Future Microbiol. 2019, 14, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Zaccaria, E.; Van Baarlen, P.; Greeff, A.D.; Morrison, D.A.; Smith, H.; Wells, J.M. Control of competence for DNA transformation in Streptococcus suis by genetically transferable pherotypes. PLoS ONE 2014, 9, e99394. [Google Scholar] [CrossRef] [Green Version]
- Unterholzner, S.J.; Poppenberger, B.; Rozhon, W. Toxin-antitoxin systems: Biology, identification, and application. Mob. Genet. Elements 2013, 3, e26219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Orejas, R.; Espinosa, M.; Yeo, C.C. The importance of the expendable: Toxin-antitoxin genes in plasmids and chromosomes. Front Microbiol. 2017, 8, 1479. [Google Scholar] [CrossRef] [PubMed]
- Harms, A.; Brodersen, D.E.; Mitarai, N.; Gerdes, K. Toxins, targets, and triggers: An overview of toxin-antitoxin biology. Mol. Cell 2018, 70, 768–784. [Google Scholar] [CrossRef] [Green Version]
- Song, P.P.; Liu, S.; Guo, X.N.; Bai, X.J.; He, X.P.; Zhang, B.R. Scarless gene deletion in methylotrophic Hansenula polymorpha by using mazF as counter-selectable marker. Anal. Biochem. 2015, 468, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Z.; Yan, X.; Cui, Z.L.; Hong, Q.; Li, S.P. mazF, a novel counter-selectable marker for unmarked chromosomal manipulation in Bacillus subtilis. Nucleic Acids Res. 2006, 34, e71. [Google Scholar] [CrossRef]
- Luo, P.; He, X.; Liu, Q.; Hu, C. Developing universal genetic tools for rapid and efficient deletion mutation in Vibrio species based on suicide T-vectors carrying a novel counterselectable marker, vmi. PLoS ONE 2015, 10, e0144465. [Google Scholar] [CrossRef] [Green Version]
- Song, Q.; Li, Z.; Chen, R.; Ma, X.; Xiao, X.; Xu, J. Induction of a toxin-antitoxin gene cassette under high hydrostatic pressure enables markerless gene disruption in the hyperthermophilic archaeon Pyrococcus yayanosii. Appl. Environ. Microbiol. 2019, 85, e02662-18. [Google Scholar] [CrossRef] [Green Version]
- Lobato-Márquez, D.; Molina-García, L.; Moreno-Córdoba, I.; García-Del Portillo, F.; Díaz-Orejas, R. Stabilization of the virulence plasmid pSLT of Salmonella Typhimurium by three maintenance systems and its evaluation by using a new stability test. Front. Mol. Biosci. 2016, 3, 66. [Google Scholar] [CrossRef] [Green Version]
- Wei, M.; Qiu, J.; Jia, M.; Zheng, C. Identification of the chromosomal YefM-YoeB toxin-antitoxin system in Vibrio parahemolyticus. Microbiol. China 2021, 1–14. Available online: http://cn.oversea.cnki.net/KCMS/detail/detail.aspx?filename=WSWT20210202000&dbcode=CJFQ&dbname=DKFX2021 (accessed on 1 April 2021).
- Zheng, C.; Jia, M.; Gao, M.; Lu, T.; Li, L.; Zhou, P. PmtA functions as a ferrous iron and cobalt efflux pump in Streptococcus suis. Emerg. Microbes Infect. 2019, 8, 1254–1264. [Google Scholar] [CrossRef] [Green Version]
- Teng, L.; Dong, X.; Zhou, Y.; Li, Z.; Deng, L.; Chen, H.; Wang, X.; Li, J. Draft genome sequence of hypervirulent and vaccine candidate Streptococcus suis strain SC19. Genome Announc. 2017, 5, e01484-16. [Google Scholar] [CrossRef] [Green Version]
- Takamatsu, D.; Osaki, M.; Sekizaki, T. Construction and characterization of Streptococcus suis-Escherichia coli shuttle cloning vectors. Plasmid 2001, 45, 101–113. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Jia, M.; Lu, T.; Gao, M.; Li, L. CopA protects Streptococcus suis against copper toxicity. Int. J. Mol. Sci. 2019, 20, 2969. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.; Xu, J.; Ren, S.; Li, J.; Xia, M.; Chen, H.; Bei, W. Identification and characterization of the chromosomal yefM-yoeB toxin-antitoxin system of Streptococcus suis. Sci. Rep. 2015, 5, 13125. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.T.; Nieto, C.; Harikrishna, J.A.; Khoo, S.K.; Othman, R.Y.; Espinosa, M.; Yeo, C.C. Genetic regulation of the yefM-yoeB toxin-antitoxin locus of Streptococcus pneumoniae. J. Bacteriol. 2011, 193, 4612–4625. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.G.; Ong, C.L.Y.; Djoko, K.Y.; West, N.P.; Davies, M.R.; McEwan, A.G.; Walker, M.J. The PerR-regulated P1B-4-type ATPase (PmtA) acts as a ferrous iron efflux pump in Streptococcus pyogenes. Infect. Immun. 2017, 85, e00140-17. [Google Scholar] [CrossRef] [Green Version]
- Brenot, A.; Weston, B.F.; Caparon, M.G. A PerR-regulated metal transporter (PmtA) is an interface between oxidative stress and metal homeostasis in Streptococcus pyogenes. Mol. Microbiol. 2007, 63, 1185–1196. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain or Plasmid | Relevant Characteristics 1 | Source or Reference |
|---|---|---|
| Strains | ||
| SC19 | Virulent S. suis strain isolated from the brain of a dead pig | [33] |
| SC19/pSET2-PcopA-yoeBVp | Strain SC19 expressing the copper-inducible YoeBVp toxin | This study |
| SC19/pSET2-PcopA-yoeBSs | Strain SC19 expressing the copper-inducible YoeBSs toxin | This study |
| SC19/pSET2 | Strain SC19 carrying the pSET2 vector | This study |
| ΔpmtA | pmtA deletion mutant of strain SC19 | This study |
| ΔperR | perR deletion mutant of strain SC19 | This study |
| ΔlysR | lysR deletion mutant of strain SC19 | This study |
| MC1061 | Cloning host for recombinant vector | AngYuBio, Shanghai, China |
| Plasmids | ||
| pSET2 | E. coli-S. suis shuttle vector; SpcR | [34] |
| pSET2-PcopA-yoeBVp | pSET2 containing the yoeBVp gene and PcopA promoter | This study |
| pSET2-PcopA-yoeBSs | pSET2 containing the yoeBSs gene and PcopA promoter | This study |
| Gene | Repetition | Spectinomycin-Sensitive Colonies (Mutants) (%) 1 | ||||
|---|---|---|---|---|---|---|
| First Dilution | Second Dilution | Third Dilution | Fourth Dilution | Fifth Dilution | ||
| pmtA | Rep_1 | 5 | 75 | 98 | ||
| Rep_2 | 3 | 91 | 98 | |||
| Rep_3 | 1 | 65 | 95 | |||
| perR | Rep_1 | 0 | 0 | 63 | 57 | 49 |
| Rep_2 | 0 | 32 | 98 | 100 | 98 | |
| Rep_3 | 0 | 5 | 83 | 93 | 97 | |
| lysR | Rep_1 | 2 | 1 | 27 | 24 | 86 |
| Rep_2 | 0 | 1 | 0 | 15 | 52 | |
| Rep_3 | 2 | 0 | 1 | 32 | 87 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, C.; Wei, M.; Qiu, J.; Li, J. A Markerless Gene Deletion System in Streptococcus suis by Using the Copper-Inducible Vibrio parahaemolyticus YoeB Toxin as a Counterselectable Marker. Microorganisms 2021, 9, 1095. https://doi.org/10.3390/microorganisms9051095
Zheng C, Wei M, Qiu J, Li J. A Markerless Gene Deletion System in Streptococcus suis by Using the Copper-Inducible Vibrio parahaemolyticus YoeB Toxin as a Counterselectable Marker. Microorganisms. 2021; 9(5):1095. https://doi.org/10.3390/microorganisms9051095
Chicago/Turabian StyleZheng, Chengkun, Man Wei, Jun Qiu, and Jinquan Li. 2021. "A Markerless Gene Deletion System in Streptococcus suis by Using the Copper-Inducible Vibrio parahaemolyticus YoeB Toxin as a Counterselectable Marker" Microorganisms 9, no. 5: 1095. https://doi.org/10.3390/microorganisms9051095
APA StyleZheng, C., Wei, M., Qiu, J., & Li, J. (2021). A Markerless Gene Deletion System in Streptococcus suis by Using the Copper-Inducible Vibrio parahaemolyticus YoeB Toxin as a Counterselectable Marker. Microorganisms, 9(5), 1095. https://doi.org/10.3390/microorganisms9051095