
A Review on the Methods Used for the Detection and Diagnosis of Rabbit Hemorrhagic Disease Virus (RHDV)



Abstract
:1. Introduction
2. Hemagglutination Test
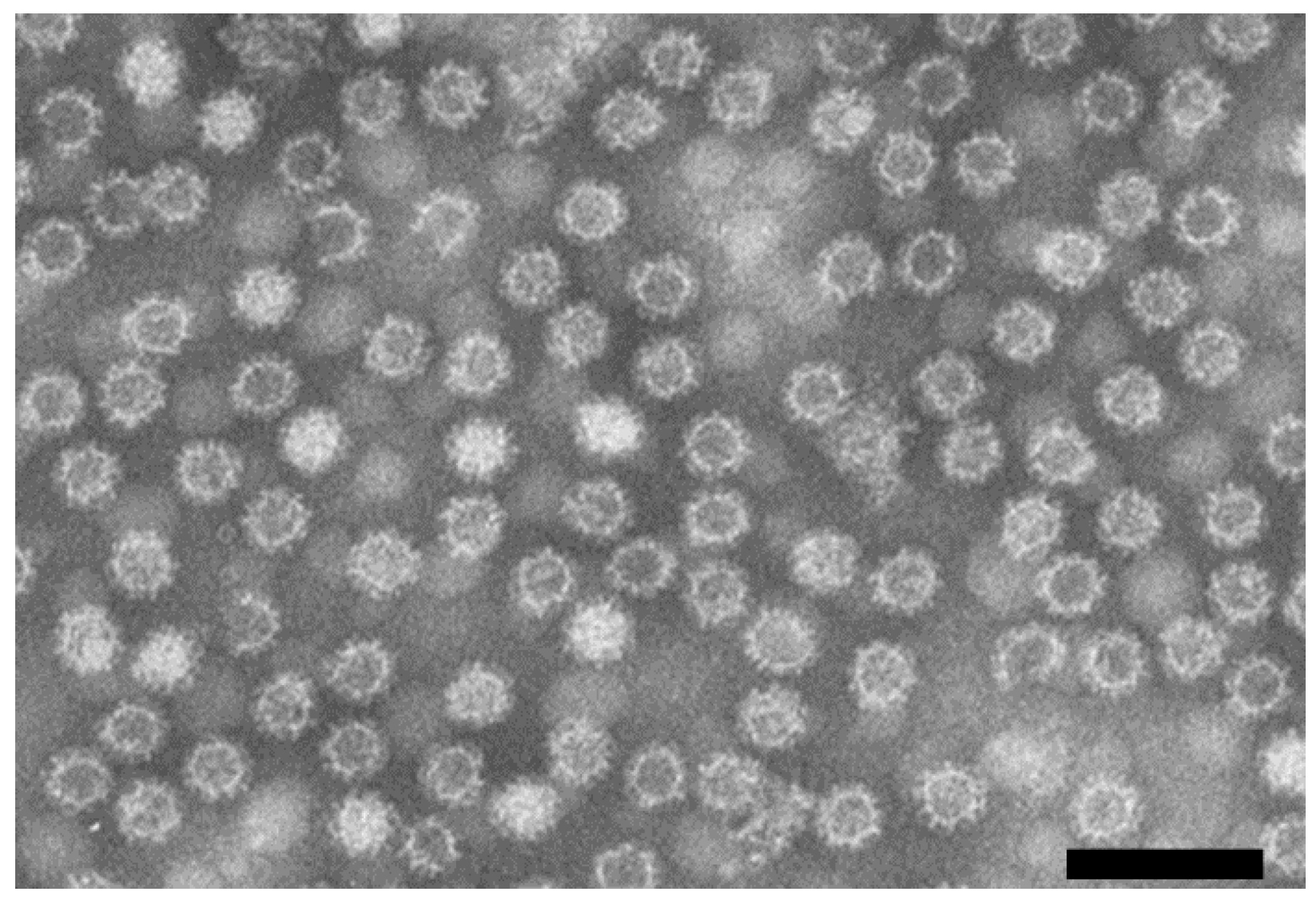
3. Electron Microscopy
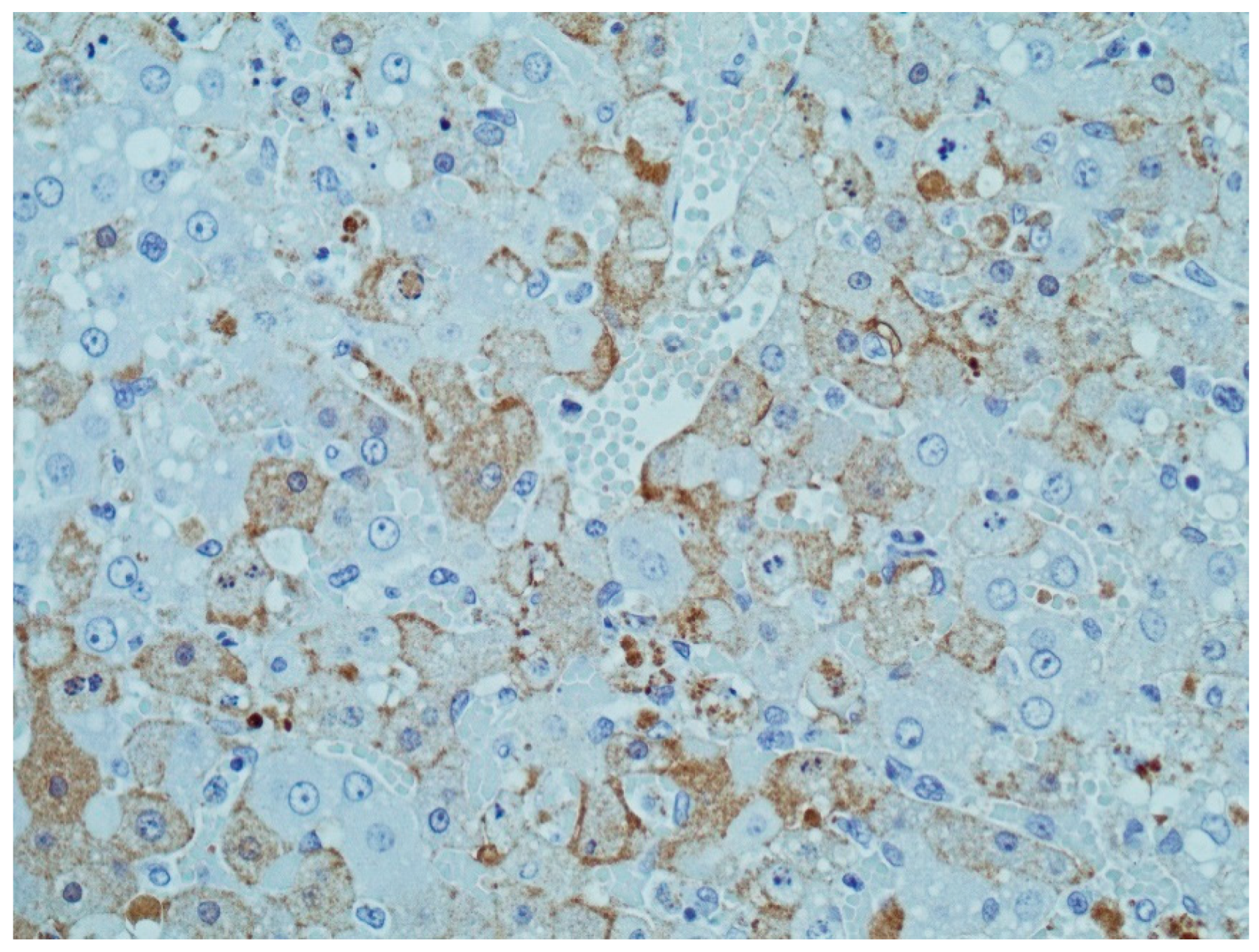
4. Immunohistochemistry
5. In Situ Hybridization
Other Labelled Probes-Based Detection Methods
6. Western Blot
7. Hemagglutination Inhibition Test
8. Enzyme-Linked Immunosorbent Assay
8.1. Antigen Capture ELISA
8.2. Antibody ELISA
9. Reverse Transcription Polymerase Chain Reaction
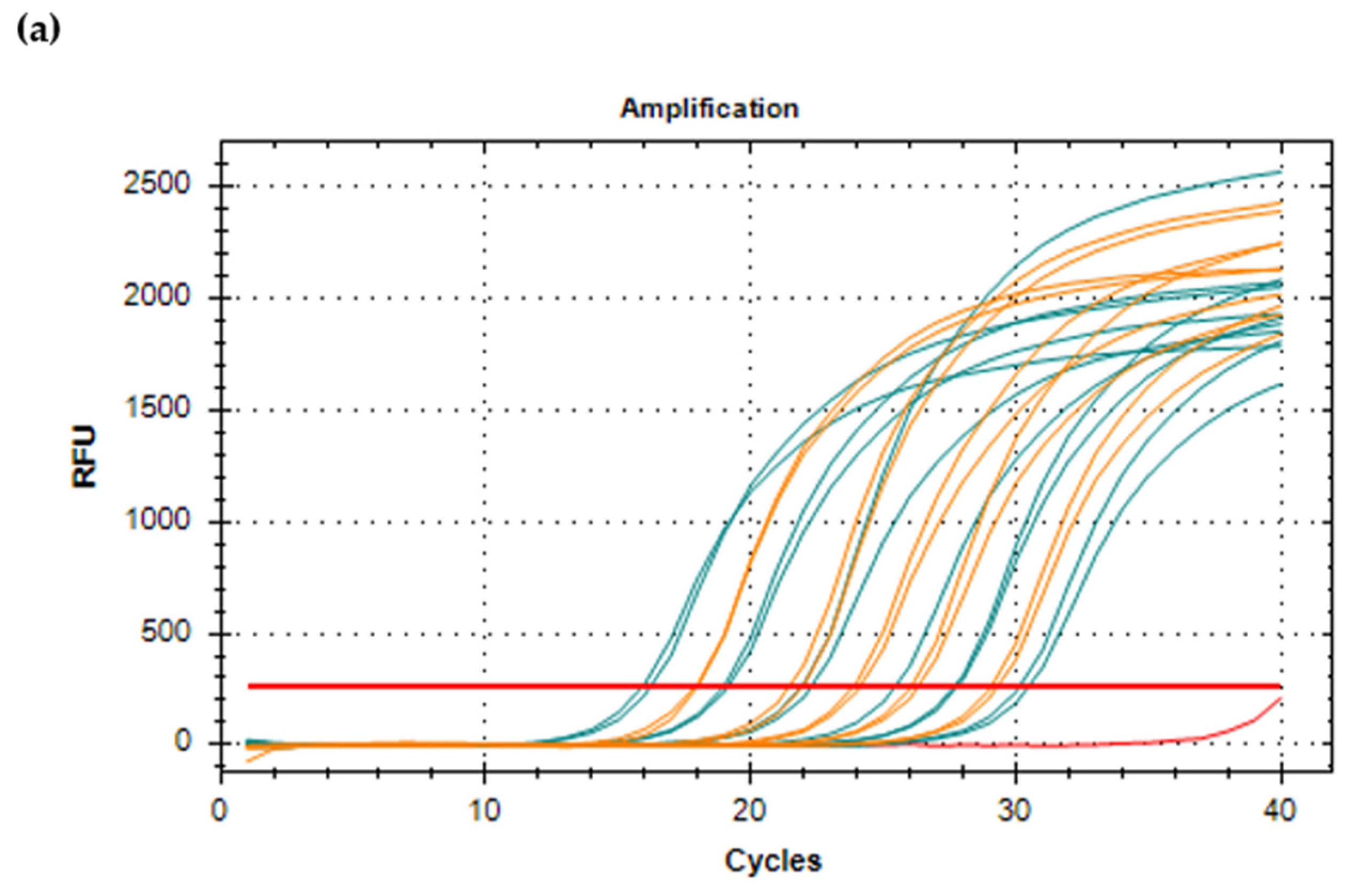
10. Real-Time Quantitative RT-PCR
11. Next-Generation Sequencing
12. Lateral Flow Immunoassay
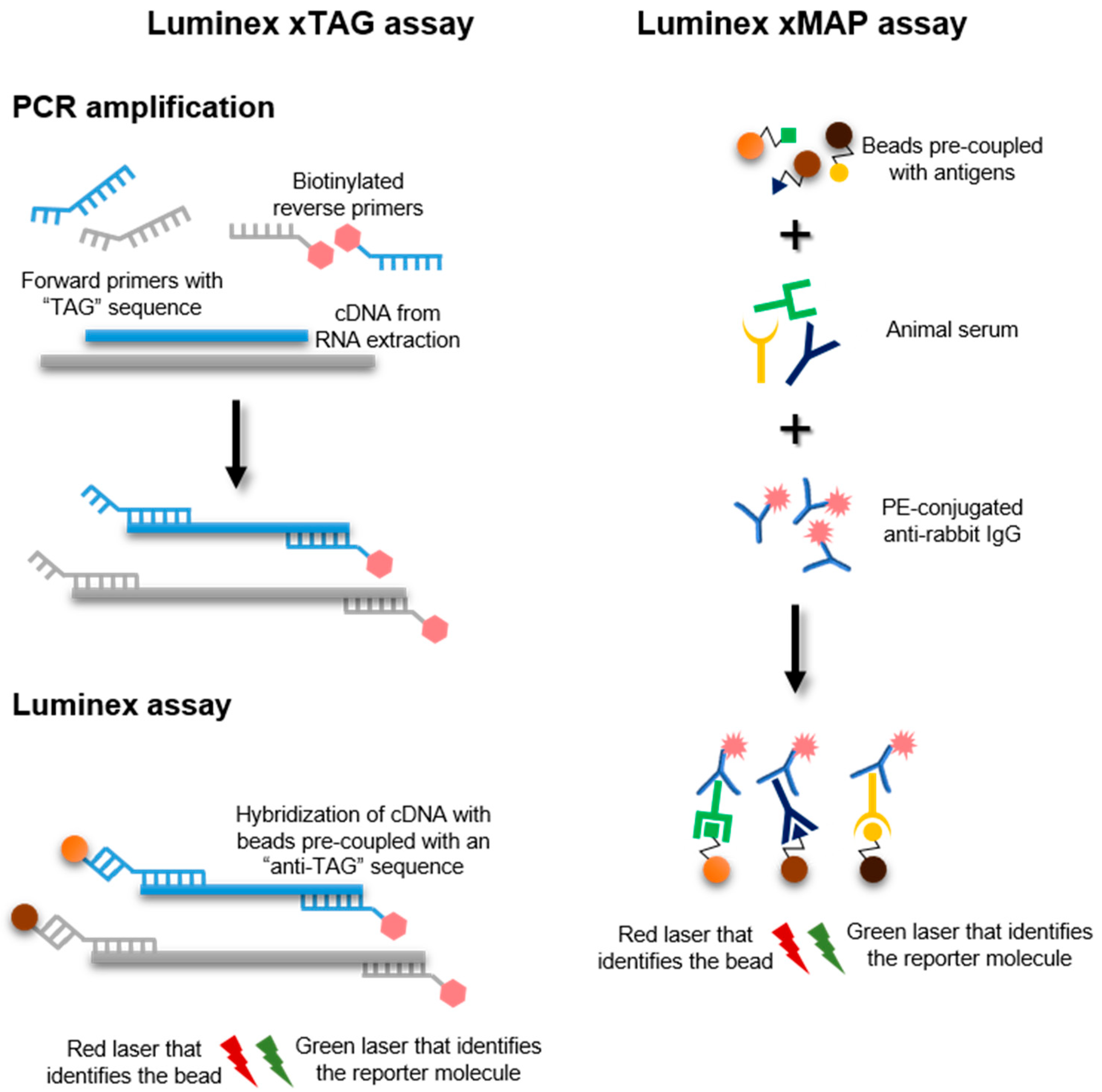
13. Luminex xTAG and xMAP Assays
14. Staphylococcus Protein A Coagglutination Test
15. Experimental Infections
16. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abrantes, J.; van der Loo, W.; Le Pendu, J.; Esteves, P.J. Rabbit haemorrhagic disease (RHD) and rabbit haemorrhagic disease virus (RHDV): A review. Vet. Res. 2012, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Xue, H.; Pu, B.; Qian, N. A new viral disease in rabbit. Anim. Husb. Vet. Med. 1984, 16, 253–255. [Google Scholar]
- Cooke, B.D.; Fenner, F. Rabbit haemorrhagic disease and the biological control of wild rabbits, Oryctolagus cuniculus, in Australia and New Zealand. Wildl. Res. 2002, 29, 689–706. [Google Scholar] [CrossRef]
- Lavazza, A.; Capucci, L. Chapter 3.6.2.—Rabbit haemorrhagic disease. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2019, 8th ed.; OIE: Paris, France, 2018; pp. 1389–1406. [Google Scholar]
- Moss, S.R.; Turner, S.L.; Trout, R.C.; White, P.J.; Hudson, P.J.; Desai, A.; Armesto, M.; Forrester, N.L.; Gould, E.A. Molecular epidemiology of Rabbit haemorrhagic disease virus. J. Gen. Virol. 2002, 83, 2461–2467. [Google Scholar] [CrossRef]
- Neimanis, A.; Larsson Pettersson, U.; Huang, N.; Gavier-Widén, D.; Strive, T. Elucidation of the pathology and tissue distribution of Lagovirus europaeus GI.2/RHDV2 (rabbit haemorrhagic disease virus 2) in young and adult rabbits (Oryctolagus cuniculus). Vet. Res. 2018, 49, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Gall, A.; Hoffmann, B.; Teifke, J.P.; Lange, B.; Schirrmeier, H. Persistence of viral RNA in rabbits which overcome an experimental RHDV infection detected by a highly sensitive multiplex real-time RT-PCR. Vet. Microbiol. 2007, 120, 17–32. [Google Scholar] [CrossRef]
- Meyers, G.; Wirblich, C.; Thiel, H.J. Rabbit hemorrhagic disease virus—Molecular cloning and nucleotide sequencing of a calicivirus genome. Virology 1991, 184, 664–676. [Google Scholar] [CrossRef]
- Wirblich, C.; Thiel, H.J.; Meyers, G. Genetic map of the calicivirus rabbit hemorrhagic disease virus as deduced from in vitro translation studies. J. Virol. 1996, 70, 7974–7983. [Google Scholar] [CrossRef] [Green Version]
- Boga, J.A.; Marín, M.S.; Casais, R.; Prieto, M.; Parra, F. In vitro translation of a subgenomic mRNA from purified virions of the Spanish field isolate AST/89 of Rabbit Hemorrhagic Disease Virus (RHDV). Virus Res. 1992, 26, 33–40. [Google Scholar] [CrossRef]
- Parra, F.; Prieto, M. Purification and characterization of a calicivirus as the causative agent of a lethal hemorrhagic disease in rabbits. J. Virol. 1990, 64, 4013–4015. [Google Scholar] [CrossRef] [Green Version]
- Henning, J.; Meers, J.; Davies, P.R.; Morris, R.S. Survival of rabbit haemorrhagic disease virus (RHDV) in the environment. Epidemiol. Infect. 2005, 133, 719–730. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.M.; Magalhães, M.J.; Alves, P.C.; Esteves, P.J.; Abrantes, J. An update on the rabbit hemorrhagic disease virus (RHDV) strains circulating in Portugal in the 1990s: Earliest detection of G3–G5 and G6. Arch. Virol. 2017, 162, 2061–2065. [Google Scholar] [CrossRef]
- Lopes, A.M.; Marques, S.; Silva, E.; Magalhaes, M.J.; Pinheiro, A.; Alves, P.C.; Le Pendu, J.; Esteves, P.J.; Thompson, G.; Abrantes, J. Detection of RHDV strains in the Iberian hare (Lepus granatensis): Earliest evidence of rabbit lagovirus cross-species infection. Vet. Res. 2014, 45, 94. [Google Scholar] [CrossRef] [Green Version]
- Le Pendu, J.; Abrantes, J.; Bertagnoli, S.; Guitton, J.-S.; Le Gall-Reculé, G.; Lopes, A.M.; Marchandeau, S.; Alda, F.; Almeida, T.; Alves, P.C.; et al. Proposal for a unified classification system and nomenclature of lagoviruses. J. Gen. Virol. 2017, 98, 1658–1666. [Google Scholar] [CrossRef]
- Le Gall-Reculé, G.; Zwingelstein, F.; Laurent, S.; de Boisséson, C.; Portejoie, Y.; Rasschaert, D. Phylogenetic analysis of rabbit haemorrhagic disease virus in France between 1993 and 2000, and the characterisation of RHDV antigenic variants. Arch. Virol. 2003, 148, 65–81. [Google Scholar] [CrossRef]
- Müller, A.; Freitas, J.; Silva, E.; Le Gall-Reculé, G.; Zwingelstein, F.; Abrantes, J.; Esteves, P.J.; Alves, P.C.; van der Loo, W.; Kolodziejek, J.; et al. Evolution of rabbit haemorrhagic disease virus (RHDV) in the European rabbit (Oryctolagus cuniculus) from the Iberian Peninsula. Vet. Microbiol. 2009, 135, 368–373. [Google Scholar] [CrossRef] [Green Version]
- Farnós, O.; Rodríguez, D.; Valdés, O.; Chiong, M.; Parra, F.; Toledo, J.R.; Fernández, E.; Lleonart, R.; Suárez, M. Molecular and antigenic characterization of rabbit hemorrhagic disease virus isolated in Cuba indicates a distinct antigenic subtype. Arch. Virol. 2007, 152, 1215–1221. [Google Scholar] [CrossRef]
- Capucci, L.; Fallacara, F.; Grazioli, S.; Lavazza, A.; Pacciarini, M.L.; Brocchi, E. A further step in the evolution of rabbit hemorrhagic disease virus: The appearance of the first consistent antigenic variant. Virus Res. 1998, 58, 115–126. [Google Scholar] [CrossRef]
- Le Gall-Reculé, G.; Zwingelstein, F.; Boucher, S.; Le Normand, B.; Plassiart, G.; Portejoie, Y.; Decors, A.; Bertagnoli, S.; Guérin, J.L.; Marchandeau, S. Detection of a new variant of rabbit haemorrhagic disease virus in France. Vet. Rec. 2011, 168, 137–138. [Google Scholar] [CrossRef] [Green Version]
- Delibes-Mateos, M.; Ferreira, C.; Carro, F.; Escudero, M.A.; Gortázar, C. Ecosystem effects of variant rabbit hemorrhagic disease virus, Iberian Peninsula. Emerg. Infect. Dis. 2014, 20, 2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neimanis, A.S.; Ahola, H.; Larsson Pettersson, U.; Lopes, A.M.; Abrantes, J.; Zohari, S.; Esteves, P.J.; Gavier-Widén, D. Overcoming species barriers: An outbreak of Lagovirus europaeus GI.2/RHDV2 in an isolated population of mountain hares (Lepus timidus). BMC Vet. Res. 2018, 14, 367. [Google Scholar] [CrossRef] [Green Version]
- Puggioni, G.; Cavadini, P.; Maestrale, C.; Scivoli, R.; Botti, G.; Ligios, C.; Le Gall-Reculé, G.; Lavazza, A.; Capucci, L. The new French 2010 Rabbit Hemorrhagic Disease Virus causes an RHD-like disease in the Sardinian Cape Hare (Lepus capensis mediterraneus). Vet. Res. 2013, 44, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camarda, A.; Pugliese, N.; Cavadini, P.; Circella, E.; Capucci, L.; Caroli, A.; Legretto, M.; Mallia, E.; Lavazza, A. Detection of the new emerging rabbit haemorrhagic disease type 2 virus (RHDV2) in Sicily from rabbit (Oryctolagus cuniculus) and Italian hare (Lepus corsicanus). Res. Vet. Sci. 2014, 97, 642–645. [Google Scholar] [CrossRef]
- Hall, R.N.; Peacock, D.E.; Kovaliski, J.; Mahar, J.E.; Mourant, R.; Piper, M.; Strive, T. Detection of RHDV2 in European brown hares (Lepus europaeus) in Australia. Vet. Rec. 2017, 180, 121. [Google Scholar] [CrossRef] [PubMed]
- Capucci, L.; Fusi, P.; Lavazza, A.; Pacciarini, M.L.; Rossi, C. Detection and preliminary characterization of a new rabbit calicivirus related to rabbit hemorrhagic disease virus but nonpathogenic. J. Virol. 1996, 70, 8614–8623. [Google Scholar] [CrossRef] [Green Version]
- Strive, T.; Wright, J.D.; Robinson, A.J. Identification and partial characterisation of a new lagovirus in Australian wild rabbits. Virology 2009, 384, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forrester, N.L.; Moss, S.R.; Turner, S.L.; Scirrmeier, H.; Gould, E.A. Recombination in rabbit haemorrhagic disease virus: Possible impact on evolution and epidemiology. Virology 2008, 376, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Abrantes, J.; Esteves, P.J.; van der Loo, W. Evidence for recombination in the major capsid gene VP60 of the rabbit haemorrhagic disease virus (RHDV). Arch. Virol. 2008, 153, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.M.; Dalton, K.P.; Magalhães, M.J.; Parra, F.; Esteves, P.J.; Holmes, E.C.; Abrantes, J. Full genomic analysis of new variant rabbit hemorrhagic disease virus revealed multiple recombination events. J. Gen. Virol. 2015, 96, 1309–1319. [Google Scholar] [CrossRef]
- Hall, R.N.; Mahar, J.E.; Read, A.J.; Mourant, R.; Piper, M.; Huang, N.; Strive, T. A strain-specific multiplex RT-PCR for Australian rabbit haemorrhagic disease viruses uncovers a new recombinant virus variant in rabbits and hares. Transbound. Emerg. Dis. 2018, 65, e444–e456. [Google Scholar] [CrossRef]
- Abrantes, J.; Droillard, C.; Lopes, A.M.; Lemaitre, E.; Lucas, P.; Blanchard, Y.; Marchandeau, S.; Esteves, P.J.; Le Gall-Reculé, G. Recombination at the emergence of the pathogenic rabbit haemorrhagic disease virus Lagovirus europaeus/GI.2. Sci. Rep. 2020, 10, 14502. [Google Scholar] [CrossRef]
- Monterroso, P.; Garrote, G.; Serronha, A.; Santos, E.; Delibes-Mateos, M.; Abrantes, J.; Perez de Ayala, R.; Silvestre, F.; Carvalho, J.; Vasco, I.; et al. Disease-mediated bottom-up regulation: An emergent virus affects a keystone prey, and alters the dynamics of trophic webs. Sci. Rep. 2016, 6, 36072. [Google Scholar] [CrossRef] [Green Version]
- Sheng, Y.X.; Wang, Z.J.; Xu, Y.F. Pathological studies on a viral septicaemia in rabbits. Shangai J. Anim. Husb. Vet. Med. 1985, 4, 3–6. (In Chinese) [Google Scholar]
- Xu, H.X.; Pu, B.Q.; Shi, D.M.; Fan, J.Z. Epidemiology of an infectious viral disease in rabbits in Wuxi Country. Shangai J. Anim. Husb. Vet. Med. 1985, 3, 14–16. (In Chinese) [Google Scholar]
- Hanchun, Y.; Xu, M.Y.; Du, N. Hemagglutination characteristics of rabbit haemorrhagic disease virus. In Proceedings of the International Symposium on Rabbit Haemorrhagic Disease, Beijing, China, 6–10 August 1991. [Google Scholar]
- Ruvoën-Clouet, N.; Ganière, J.P.; André-Fontaine, G.; Blanchard, D.; Le Pendu, J. Binding of rabbit hemorrhagic disease virus to antigens of the ABO histo-blood group family. J. Virol. 2000, 74, 11950–11954. [Google Scholar] [CrossRef] [Green Version]
- Oriol, R.A.; Mollicone, R.O.; Coullin, P.H.; Dalix, A.M.; Candelier, J.J. Genetic regulation of the expression of ABH and Lewis antigens in tissues. APMIS Suppl. 1992, 27, 28–38. [Google Scholar]
- Xu, W.X. Viral haemorrhagic disease of rabbits in the People’s Republic of China: Epidemiology and virus characterization. Int. Off. Epizoot. 1991, 10, 393–408. [Google Scholar] [CrossRef] [Green Version]
- Pu, B.; Quian, N.H.; Cui, S.J. Micro HA and HI tests for the detection of antibody titres to so-called “haemorrhagic pneumonia” in rabbits. Chin. J. Vet. Med. 1985, 11, 16–17. (In Chinese) [Google Scholar]
- Ahmad, S.T.; El-Samadony, H.A.; Mahgoub, K.M. Immunological and virological studies on rabbit hemorrhagic disease virus. Glob. Vet. 2011, 7, 545–556. [Google Scholar]
- Capucci, L.; Scicluna, M.T.; Lavazza, A. Diagnosis of viral haemorrhagic disease of rabbits and the European brown hare syndrome. Rev. Sci. Tech. Off. Int. Epizoot. 1991, 10, 347–370. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.K. A new viral disease in rabbits—Rabbit pest. Sci. Agric. Sin. 1988, 21, 6–11. (In Chinese) [Google Scholar]
- Lei, Y.X.; Yaol, T.; He, S.L. Detection of haemagglutination titre in tissues from rabbits infected with viral haemorrhagic disease. Shanghai J. Anim. Husb. Vet. Med. 1985, 6, 19–20. (In Chinese) [Google Scholar]
- Granzow, H.; Weiland, F.; Strebelow, H.G.; Liu, C.M.; Schirrmeier, H. Rabbit hemorrhagic disease virus (RHDV): Ultrastructure and biochemical studies of typical and core-like particles present in liver homogenates. Virus Res. 1996, 41, 163–172. [Google Scholar] [CrossRef]
- Rabbit Haemorrhagic Disease. OIE Technical Disease Cards. Available online: https://www.oie.int/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Disease_cards/RHD.pdf (accessed on 27 December 2020).
- Chasey, D.; Lucas, M.H.; Westcott, D.G.; Sharp, G.; Kitching, A.; Hughes, S.K. Development of a diagnostic approach to the identification of rabbit haemorrhagic disease. Vet. Rec. 1995, 137, 158–160. [Google Scholar] [CrossRef]
- Tian, L.; Liao, J.; Li, J.-W.; Zhou, W.-R.; Zhang, X.-L.; Wang, H.-N. Isolation and identification of a non-haemagglutinating strain of rabbit hemorrhagic disease virus from China and sequence analysis for the VP60 Gene. Virus Genes 2007, 35, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Shakal, M.A.; Khelfa, D.E.; Salman, O.G.A.; Yousif, A.A.; Salwa, E.A.A. Use of VP60 RT-PCR to overcome the limitation of haemagglutination inhibition diagnosis of rabbit viral haemorrhagic disease. World Rabbit Sci. 2011, 19. [Google Scholar] [CrossRef] [Green Version]
- Biermann, U.; Herbst, W.; Fau-Baljer, G.; Baljer, G. Rabbit hemorrhagic disease (RHD)-comparative diagnostic studies using the hemagglutination test and electron microscopy. Berl. Munch. Tierarztl. Wochenschr. 1992, 105, 86–87. (In Chinese) [Google Scholar] [PubMed]
- Kesy, A.; Fitzner, A.; Niedbalski, W.; Paprocka, G.; Walkowiak, B. A new variant of the viral haemorrhagic disease of rabbits virus. Rev. Sci. Tech. Off. Int. Epizoot. 1996, 15, 1029–1035. [Google Scholar] [CrossRef]
- Prieto, J.M.; Martin, J.M.; Espi, A.; Parra, F. A new nonhaemagglutinating strain of RHDV. In Proceedings of the 5th International Congress of the European Society for Veterinary Virology, Brescia, Italy, 27–30 August; pp. 204–205.
- Capucci, L.; Chasey, D.; Lavazza, A.; Westcott, D. Preliminary characterization of a non-haemagglutinating strain of rabbit haemorrhagic disease virus from the United Kingdom. Zentralblatt fur Veterinarmedizin. Reihe B 1996, 43, 245–250. [Google Scholar] [CrossRef]
- Le Gall-Reculé, G.; Lavazza, A.; Marchandeau, S.; Bertagnoli, S.; Zwingelstein, F.; Cavadini, P.; Martinelli, N.; Lombardi, G.; Guerin, J.-L.; Lemaitre, E.; et al. Emergence of a new lagovirus related to Rabbit Haemorrhagic Disease Virus. Vet. Res. 2013, 44, 81. [Google Scholar] [CrossRef] [Green Version]
- Dalton, K.P.; Nicieza, I.; Balseiro, A.; Muguerza, M.A.; Rosell, J.M.; Casais, R.; Álvarez, Á.L.; Parra, F. Variant rabbit hemorrhagic disease virus in young rabbits, Spain. Emerg. Infect. Dis. 2012, 18, 2009–2012. [Google Scholar] [CrossRef] [PubMed]
- Ohlinger, V.F.; Haas, B.; Meyers, G.; Weiland, F.; Thiel, H.J. Identification and characterization of the virus causing rabbit hemorrhagic disease. J. Virol. 1990, 64, 3331–3336. [Google Scholar] [CrossRef] [Green Version]
- Valíček, L.; Šmíd, B.; Rodák, L.; Kudrna, J. Electron and immunoelectron microscopy of rabbit haemorrhagic disease virus (RHDV). Arch. Virol. 1990, 112, 271–275. [Google Scholar] [CrossRef]
- Xu, Z.J.; Chen, W.X. Viral hemorrhagic disease in rabbits: A review. Vet. Res. Commun. 1989, 13, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Schramlová, J.; Arientová, S.; Hulínská, D. The role of electron microscopy in the rapid diagnosis of viral infections--review. Folia Microbiol. 2010, 55, 88–101. [Google Scholar] [CrossRef] [PubMed]
- Embury-Hyatt, C.; Postey, R.; Hisanaga, T.; Burton, L.; Hooper-McGrevy, K.; McIntyre, L.; Millar, K.; Pasick, J. The first reported case of rabbit hemorrhagic disease in Canada. Can. Vet. J. Rev. Vet. Can. 2012, 53, 998–1002. [Google Scholar]
- Schirrmeier, H.; Reimann, I.; Köllner, B.; Granzow, H. Pathogenic, antigenic and molecular properties of rabbit haemorrhagic disease virus (RHDV) isolated from vaccinated rabbits: Detection and characterization of antigenic variants. Arch. Virol. 1999, 144, 719–735. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Itakura, C. Detection of rabbit haemorrhagic disease virus antigen in tissues by immunohistochemistry. Res. Vet. Sci. 1992, 52, 299–306. [Google Scholar] [CrossRef]
- Park, J.H.; Ochiai, K.; Itakura, C. Detection of rabbit haemorrhagic disease virus particles in the rabbit liver tissues. J. Comp. Pathol. 1992, 107, 329–340. [Google Scholar] [CrossRef]
- Lavazza, A.; Scicluna, M.T.; Capucci, L. Susceptibility of hares and rabbits to the European brown hare syndrome virus (EBHSV) and rabbit haemorrhagic disease virus (RHDV) under experimental conditions. Zentralbl. Veterinarmed. B 1996, 43, 401–410. [Google Scholar] [CrossRef]
- Nagesha, H.S.; Wang, L.F.; Hyatt, A.D.; Morrissy, C.J.; Lenghaus, C.; Westbury, H.A. Self-assembly, antigenicity, and immunogenicity of the rabbit haemorrhagic disease virus (Czechoslovakian strain V-351) capsid protein expressed in baculovirus. Arch. Virol. 1995, 140, 1095–1108. [Google Scholar] [CrossRef] [PubMed]
- Peacey, M.; Wilson, S.; Baird, M.A.; Ward, V.K. Versatile RHDV virus-like particles: Incorporation of antigens by genetic modification and chemical conjugation. Biotechnol. Bioeng. 2007, 98, 968–977. [Google Scholar] [CrossRef] [PubMed]
- Qi, R.; Miao, Q.; Zhu, J.; Tang, J.; Tang, A.; Wang, X.; Dong, D.; Guo, H.; Liu, G. Construction and immunogenicity of novel bivalent virus-like particles bearing VP60 genes of classic RHDV(GI.1) and RHDV2(GI.2). Vet. Microbiol. 2020, 240, 108529. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.; Ulrich, R.; Franzke, K.; Müller, M.; Köllner, B. Crude extracts of recombinant baculovirus expressing rabbit hemorrhagic disease virus 2 VLPs from both insect and rabbit cells protect rabbits from rabbit hemorrhagic disease caused by RHDV2. Arch. Virol. 2019, 164, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Tian, X.; Zhai, Y.; Xu, W.; Zheng, D.; Sun, F. Cryo-electron microscopy reconstructions of two types of wild rabbit hemorrhagic disease viruses characterized the structural features of Lagovirus. Protein Cell 2010, 1, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xu, F.; Liu, J.; Gao, B.; Liu, Y.; Zhai, Y.; Ma, J.; Zhang, K.; Baker, T.S.; Schulten, K.; et al. Atomic model of rabbit hemorrhagic disease virus by cryo-electron microscopy and crystallography. PLoS Pathog. 2013, 9, e1003132. [Google Scholar] [CrossRef] [Green Version]
- Katpally, U.; Voss, N.R.; Cavazza, T.; Taube, S.; Rubin, J.R.; Young, V.L.; Stuckey, J.; Ward, V.K.; Virgin, H.W.t.; Wobus, C.E.; et al. High-resolution cryo-electron microscopy structures of murine norovirus 1 and rabbit hemorrhagic disease virus reveal marked flexibility in the receptor binding domains. J. Virol. 2010, 84, 5836–5841. [Google Scholar] [CrossRef] [Green Version]
- Herzik, M.A.J. Cryo-electron microscopy reaches atomic resolution. Nature 2020, 587, 39–40. [Google Scholar] [CrossRef]
- van de Bildt, M.W.G.; van Bolhuis, G.H.; van Zijderveld, F.; van Riel, D.; Drees, J.M.; Osterhaus, A.D.M.E.; Kuiken, T. Confirmation and Phylogenetic Analysis of Rabbit Hemorrhagic Disease Virus in Free-living Rabbits from the Netherlands. J. Wildl. Dis. 2006, 42, 808–812. [Google Scholar] [CrossRef] [Green Version]
- Marques, R.M.; Teixeira, L.; Águas, A.P.; Ribeiro, J.C.; Costa-e-Silva, A.; Ferreira, P.G. Immunosuppression abrogates resistance of young rabbits to Rabbit Haemorrhagic Disease (RHD). Vet. Res. 2014, 45, 14. [Google Scholar] [CrossRef] [Green Version]
- Nyström, K.; Le Gall-Reculé, G.; Grassi, P.; Abrantes, J.; Ruvoën-Clouet, N.; Le Moullac-Vaidye, B.; Lopes, A.M.; Esteves, P.J.; Strive, T.; Marchandeau, S.; et al. Histo-blood group antigens act as attachment factors of rabbit hemorrhagic disease virus infection in a virus strain-dependent manner. PLoS Pathog. 2011, 7, e1002188. [Google Scholar] [CrossRef] [Green Version]
- Stoercklé-Berger, N.; Keller-Berger, B.; Ackermann, M.; Ehrensperger, F. Immunohistological Diagnosis of Rabbit Haemorrhagic Disease (RHD). J. Vet. Med. Ser. B 1992, 39, 237–245. [Google Scholar] [CrossRef]
- Jung, J.Y.; Lee, B.J.; Tai, J.H.; Park, J.H.; Lee, Y.S. Apoptosis in Rabbit Haemorrhagic Disease. J. Comp. Pathol. 2000, 123, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Alonso, C.; Oviedo, J.M.; Martín-Alonso, J.M.; Díaz, E.; Boga, J.A.; Parra, F. Programmed cell death in the pathogenesis of rabbit hemorrhagic disease. Arch. Virol. 1998, 143, 321–332. [Google Scholar] [CrossRef]
- Ramiro-Ibáñez, F.; Martı́n-Alonso, J.M.; Garcı́a Palencia, P.; Parra, F.; Alonso, C. Macrophage tropism of rabbit hemorrhagic disease virus is associated with vascular pathology. Virus Res. 1999, 60, 21–28. [Google Scholar] [CrossRef]
- Mikami, O.; Park, J.H.; Kimura, T.; Ochiai, K.; Itakura, C. Hepatic lesions in young rabbits experimentally infected with rabbit haemorrhagic disease virus. Res. Vet. Sci. 1999, 66, 237–242. [Google Scholar] [CrossRef]
- Prieto, J.M.; Fernandez, F.; Alvarez, V.; Espi, A.; Garc, Í.A.; MarÍN, J.F.; Alvarez, M.; MartÍN, J.M.; Parra, F. Immunohistochemical localisation of rabbit haemorrhagic disease virus VP-60 antigen in early infection of young and adult rabbits. Res. Vet. Sci. 2000, 68, 181–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marques, R.M.; Costa-E-Silva, A.; Águas, A.P.; Teixeira, L.; Ferreira, P.G. Early inflammatory response of young rabbits attending natural resistance to calicivirus (RHDV) infection. Vet. Immunol. Immunopathol. 2012, 150, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Gelmetti, D.; Grieco, V.; Rossi, C.; Capucci, L.; Lavazza, A. Detection of rabbit haemorrhagic disease virus (RHDV) by in situ hybridisation with a digoxigenin labelled RNA probe. J. Virol. Methods 1998, 72, 219–226. [Google Scholar] [CrossRef]
- Wang, L.; Xia, T.; Guo, T.; Ru, Y.; Jiang, Y.; Cui, W.; Zhou, H.; Qiao, X.; Tang, L.; Xu, Y.; et al. Recombinant Lactobacillus casei Expressing Capsid Protein VP60 can Serve as Vaccine Against Rabbit Hemorrhagic Disease Virus in Rabbits. Vaccines 2019, 7, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, D.; Qu, L.; Liu, J.; Guo, D.; Jiang, Q.; Lin, H.; Si, C. DNA vaccination with a gene encoding VP60 elicited protective immunity against rabbit hemorrhagic disease virus. Vet. Microbiol. 2013, 164, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Yu, Z.; Liu, J.; Kong, D.; Guo, D.; Quan, C.; Li, B.; Hu, X.; Qu, L. Recombinant canine adenovirus type 2 expressing rabbit hemorrhagic disease virus VP60 protein provided protection against RHD in rabbits. Vet. Microbiol. 2018, 213, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Collins, B.J.; White, J.R.; Lenghaus, C.; Boyd, V.; Westbury, H.A. A competition ELISA for the detection of antibodies to rabbit haemorrhagic disease virus. Vet. Microbiol. 1995, 43, 85–96. [Google Scholar] [CrossRef]
- Kong, D.; Liu, J.; Jiang, Q.; Yu, Z.; Hu, X.; Guo, D.; Huang, Q.; Jiao, M.; Qu, L. Production, Characterization, and Epitope Mapping of Monoclonal Antibodies Against Different Subtypes of Rabbit Hemorrhagic Disease Virus (RHDV). Sci. Rep. 2016, 6, 20857. [Google Scholar] [CrossRef] [PubMed]
- Collins, B.J.; White, J.R.; Lenghaus, C.; Morrissy, C.J.; Westbury, H.A. Presence of rabbit haemorrhagic disease virus antigen in rabbit tissues as revealed by a monoclonal antibody dependent capture ELISA. J. Virol. Methods 1996, 58, 145–154. [Google Scholar] [CrossRef]
- Kimura, T.; Mitsui, I.; Okada, Y.; Furuya, T.; Ochiai, K.; Umemura, T.; Itakura, C. Distribution of Rabbit Haemorrhagic Disease Virus RNA in Experimentally Infected Rabbits. J. Comp. Pathol. 2001, 124, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Hoehn, M.; Kerr, P.J.; Strive, T. In situ hybridisation assay for localisation of rabbit calicivirus Australia-1 (RCV-A1) in European rabbit (Oryctolagus cuniculus) tissues. J. Virol. Methods 2013, 188, 148–152. [Google Scholar] [CrossRef]
- Guittré, C.; Baginski, I.; Le Gall, G.; Prave, M.; Trépo, C.; Cova, L. Detection of rabbit haemorrhagic disease virus isolates and sequence comparison of the N-terminus of the capsid protein gene by the polymerase chain reaction. Res. Vet. Sci. 1995, 58, 128–132. [Google Scholar] [CrossRef]
- Tham, K.; Barnes, S.M.; Hunter, S.N. Polymerase Chain Reaction Amplification and Gene Sequence Analysis of a Calicivirus from a Feral Rabbit. Virus Genes 1999, 18, 235–242. [Google Scholar] [CrossRef]
- Pšikal, I.; Šmíd, B.; Kubalíková, R.; Valíček, L.; Rodák, L.; Kosinová, E. Colorimetric detection of lagomorphs’ calicivirus genomic sequences by polymerase chain reaction incorporating digoxigenin dUTP. Vet. Microbiol. 1997, 57, 55–67. [Google Scholar] [CrossRef]
- Ruvoen-Clouet, N.; Blanchard, D.; Andre-Fontaine, G.; Song, B.; Ganiere, J.P. Detection of Antibodies to Rabbit Haemorrhagic Disease Virus: An Immunoblotting Method using Virus-coated Human Erythrocyte Membranes. J. Vet. Med. Ser. B 1995, 42, 197–204. [Google Scholar] [CrossRef]
- Rodák, L.; Šmíd, B.; Valíček, L.; Veselý, T.; Štěpánek, J.; Hampl, J.; Jurák, E. Enzyme-linked immunosorbent assay of antibodies to rabbit haemorrhagic disease virus and determination of its major structural proteins. J. Gen. Virol. 1990, 71, 1075–1080. [Google Scholar] [CrossRef]
- Farnós, O.; Boué, O.; Parra, F.; Martín-Alonso, J.M.; Valdés, O.; Joglar, M.; Navea, L.; Naranjo, P.; Lleonart, R. High-level expression and immunogenic properties of the recombinant rabbit hemorrhagic disease virus VP60 capsid protein obtained in Pichia pastoris. J. Biotechnol. 2005, 117, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qiu, L.; Hao, H.; Zhang, W.; Fu, X.; Zhang, H.; He, S.; Zhang, S.; Du, E.; Yang, Z. Adenovirus-based oral vaccine for rabbit hemorrhagic disease. Vet. Immunol. Immunopathol. 2012, 145, 277–282. [Google Scholar] [CrossRef]
- Rodák, L.; Granátová, M.; Valíček, L.; Šmíd, B.; Veselý, T.; Nevoránková, Z. Monoclonal antibodies to rabbit haemorrhagic disease virus and their use in the diagnosis of infection. J. Gen. Virol. 1990, 71, 2593–2598. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Gilda, J.E.; Gomes, A.V. The necessity of and strategies for improving confidence in the accuracy of western blots. Expert Rev. Proteom. 2014, 11, 549–560. [Google Scholar] [CrossRef] [Green Version]
- Robinson, A.J.; Kirkland, P.D.; Forrester, R.I.; Capucci, L.; Cooke, B.D.; Philbey, A.W. Serological evidence for the presence of a calicivirus in Australian wild rabbits, Oryctolagus cuniculus, before the introduction of rabbit haemorrhagic disease virus (RHDV): Its potential influence on the specificity of a competitive ELISA for RHDV. Wildl. Res. 2002, 29, 655–662. [Google Scholar] [CrossRef]
- Marchandeau, S.; Le Gall-Reculé, G.; Bertagnoli, S.; Aubineau, J.; Botti, G.; Lavazza, A. Serological evidence for a non-protective RHDV-like virus. Vet. Res. 2005, 36, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Trout, R.C.; Chasey, D.; Sharp, G. Seroepidemiology of rabbit haemorrhagic disease (RHD) in wild rabbits (Oryctolagus cuniculus) in the United Kingdom. J. Zool. 1997, 243, 846–853. [Google Scholar] [CrossRef]
- Capucci, L.; Lavazza, A.; Civardi, A.; Frigoli, G.; Barbieri, I.; Rossi, C. Seroconversion in asymptomatic rabbits provides further evidence of putative RHDV-like virus. In Proceedings of the Third Congress of the European Society of Veterinary Virology, Intelaken, Switzerland, 4–7 September 1994; pp. 88–93. [Google Scholar]
- Nagesha, H.S.; McColl, K.A.; Collins, B.J.; Morrissy, C.J.; Wang, L.F.; Westbury, H.A. The presence of cross-reactive antibodies to rabbit haemorrhagic disease virus in Australian wild rabbits prior to the escape of virus from quarantine. Arch. Virol. 2000, 145, 749–757. [Google Scholar] [CrossRef]
- Frölich, K.; Klima, F.; Dedek, J. Antibodies against rabbit hemorrhagic disease virus in free-ranging red foxes from Germany. J. Wildl. Dis. 1998, 34, 436–442. [Google Scholar] [CrossRef] [Green Version]
- Henning, J.; Davies, P.R.; Meers, J. Seropositivity to rabbit haemorrhagic disease virus in non-target mammals during periods of viral activity in a population of wild rabbits in New Zealand. Wildl. Res. 2006, 33, 305–311. [Google Scholar] [CrossRef]
- Parkes, J.P.; Heyward, R.P.; Henning, J.; Motha, M.X. Antibody responses to rabbit haemorrhagic disease virus in predators, scavengers, and hares in New Zealand during epidemics in sympatric rabbit populations. N. Z. Vet. J. 2004, 52, 85–89. [Google Scholar] [CrossRef]
- Dalton, K.P.; Podadera, A.; Granda, V.; Nicieza, I.; del Llano, D.; González, R.; de los Toyos, J.R.; García Ocaña, M.; Vázquez, F.; Martín Alonso, J.M.; et al. ELISA for detection of variant rabbit haemorrhagic disease virus RHDV2 antigen in liver extracts. J. Virol. Methods 2018, 251, 38–42. [Google Scholar] [CrossRef]
- Bouslama, A.; De Mia, G.M.; Hammami, S.; Aouina, T.; Soussi, H.; Frescura, T. Identification of the virus of rabbit haemorrhagic disease in Tunisia. Vet. Rec. 1996, 138, 108–110. [Google Scholar] [CrossRef]
- Zheng, T.; Lu, G.; Napier, A.M.; Lockyer, S.J. No virus replication in domestic cats fed with RHDV-infected rabbit livers. Vet. Microbiol. 2003, 95, 61–73. [Google Scholar] [CrossRef]
- Campagnolo, E.R.; Ernst, M.J.; Berninger, M.L.; Gregg, D.A.; Shumaker, T.J.; Boghossian, A.M. Outbreak of rabbit hemorrhagic disease in domestic lagomorphs. J. Am. Vet. Med. Assoc. 2003, 223, 1151–1155. [Google Scholar] [CrossRef]
- McIntosh, M.T.; Behan, S.C.; Mohamed, F.M.; Lu, Z.; Moran, K.E.; Burrage, T.G.; Neilan, J.G.; Ward, G.B.; Botti, G.; Capucci, L.; et al. A pandemic strain of calicivirus threatens rabbit industries in the Americas. Virol. J. 2007, 4, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovaliski, J. Monitoring the spread of rabbit hemorrhagic disease virus as a new biological agent for control of wild European rabbits in Australia. J. Wildl. Dis. 1998, 34, 421–428. [Google Scholar] [CrossRef]
- Cooke, B.D.; Robinson, A.J.; Merchant, J.C.; Nardin, A.; Capucci, L. Use of ELISAs in field studies of rabbit haemorrhagic disease (RHD) in Australia. Epidemiol. Infect. 2000, 124, 563–576. [Google Scholar] [CrossRef]
- Mahar, J.E.; Read, A.J.; Gu, X.; Urakova, N.; Mourant, R.; Piper, M.; Haboury, S.; Holmes, E.C.; Strive, T.; Hall, R.N. Detection and Circulation of a Novel Rabbit Hemorrhagic Disease Virus in Australia. Emerg. Infect. Dis. 2018, 24, 22–31. [Google Scholar] [CrossRef]
- Martin-Alonso, A.; Martin-Carrillo, N.; Garcia-Livia, K.; Valladares, B.; Foronda, P. Emerging rabbit haemorrhagic disease virus 2 (RHDV2) at the gates of the African continent. Infect. Genet. Evol. 2016, 44, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Dalton, K.P.; Nicieza, I.; Abrantes, J.; Esteves, P.J.; Parra, F. Spread of new variant RHDV in domestic rabbits on the Iberian Peninsula. Vet. Microbiol. 2014, 169, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Buehler, M.; Jesse, S.T.; Kueck, H.; Lange, B.; Koenig, P.; Jo, W.K.; Osterhaus, A.; Beineke, A. Lagovirus europaeus GI.2 (rabbit hemorrhagic disease virus 2) infection in captive mountain hares (Lepus timidus) in Germany. BMC Vet. Res. 2020, 16, 166. [Google Scholar] [CrossRef] [PubMed]
- Zheng, T.; Parkes, J.P. Rabbit haemorrhagic disease: Advantages of cELISA in assessing immunity in wild rabbits (Oryctolagus cuniculus). Vet. Microbiol. 2011, 153, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Rouco, C.; Abrantes, J.; Serronha, A.; Lopes, A.M.; Maio, E.; Magalhães, M.J.; Blanco, E.; Bárcena, J.; Esteves, P.J.; Santos, N.; et al. Epidemiology of RHDV2 (Lagovirus europaeus/GI.2) in free-living wild European rabbits in Portugal. Transbound. Emerg. Dis. 2017, 65, e373–e382. [Google Scholar] [CrossRef]
- Strive, T.; Piper, M.; Huang, N.; Mourant, R.; Kovaliski, J.; Capucci, L.; Cox, T.E.; Smith, I. Retrospective serological analysis reveals presence of the emerging lagovirus RHDV2 in Australia in wild rabbits at least five months prior to its first detection. Transbound. Emerg. Dis. 2020, 67, 822–833. [Google Scholar] [CrossRef]
- Lavazza, A.; Cavadini, P.; Barbieri, I.; Tizzani, P.; Pinheiro, A.; Abrantes, J.; Esteves, P.J.; Grilli, G.; Gioia, E.; Zanoni, M.; et al. Field and experimental data indicate that the eastern cottontail (Sylvilagus floridanus) is susceptible to infection with European brown hare syndrome (EBHS) virus and not with rabbit haemorrhagic disease (RHD) virus. Vet. Res. 2015, 46, 13. [Google Scholar] [CrossRef] [Green Version]
- Henning, J.; Meers, J.; Davies, P.R. Exposure of rabbits to ultraviolet light-inactivated rabbit haemorrhagic disease virus (RHDV) and subsequent challenge with virulent virus. Epidemiol. Infect. 2005, 133, 731–735. [Google Scholar] [CrossRef] [Green Version]
- Capucci, L.; Nardin, A.; Lavazza, A. Seroconversion in an industrial unit of rabbits infected with a non-pathogenic rabbit haemorrhagic disease-like virus. Vet. Rec. 1997, 140, 647. [Google Scholar] [CrossRef]
- Philbey, A.W.; Kirkland, P.D.; Saunders, G.R. Assessment of antibodies to rabbit haemorrhagic disease virus in fox serum as an indicator of infection in sympatric rabbit populations. Aust. Vet. J. 2005, 83, 97–100. [Google Scholar] [CrossRef]
- Liu, J.; Kerr, P.J.; Strive, T. A sensitive and specific blocking ELISA for the detection of rabbit calicivirus RCV-A1 antibodies. Virol. J. 2012, 9, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Kerr, P.J.; Wright, J.D.; Strive, T. Serological assays to discriminate rabbit haemorrhagic disease virus from Australian non-pathogenic rabbit calicivirus. Vet. Microbiol. 2012, 157, 345–354. [Google Scholar] [CrossRef]
- Belák, S.; Ballagi-Pordány, A. Application of the polymerase chain reaction (PCR) in veterinary diagnostic virology. Vet. Res. Commun. 1993, 17, 55–72. [Google Scholar] [CrossRef]
- Meyers, G.; Wirblich, C.; Thiel, H.J. Genomic and subgenomic RNAs of rabbit hemorrhagic disease virus are both protein-linked and packaged into particles. Virology 1991, 184, 677–686. [Google Scholar] [CrossRef]
- Yang, L.; Wang, F.; Hu, B.; Xue, J.; Hu, Y.; Zhou, B.; Wang, D.; Xu, W. Development of an RT-PCR for rabbit hemorrhagic disease virus (RHDV) and the epidemiology of RHDV in three eastern provinces of China. J. Virol. Methods 2008, 151, 24–29. [Google Scholar] [CrossRef]
- Schwensow, N.I.; Cooke, B.; Kovaliski, J.; Sinclair, R.; Peacock, D.; Fickel, J.; Sommer, S. Rabbit haemorrhagic disease: Virus persistence and adaptation in Australia. Evol. Appl. 2014, 7, 1056–1067. [Google Scholar] [CrossRef]
- Abrantes, J.; Lopes, A.M.; Esteves, P.J. Complete genomic sequences of rabbit hemorrhagic disease virus G1 strains isolated in the European rabbit original range. J. Virol. 2012, 86, 13886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalton, K.P.; Arnal, J.L.; Benito, A.A.; Chacón, G.; Martín Alonso, J.M.; Parra, F. Conventional and real time RT-PCR assays for the detection and differentiation of variant rabbit hemorrhagic disease virus (RHDVb) and its recombinants. J. Virol. Methods 2018, 251, 118–122. [Google Scholar] [CrossRef]
- Bascuñana, C.R.; Nowotny, N.; Belák, S. Detection and differentiation of rabbit hemorrhagic disease and European brown hare syndrome viruses by amplification of VP60 genomic sequences from fresh and fixed tissue specimens. J. Clin. Microbiol. 1997, 35, 2492–2495. [Google Scholar] [CrossRef] [Green Version]
- Rozas, A.P.D.; González, J.; Aloy, N.; Badiola, I.; Xicato, G.; Trocino, A.; Lukefahr, S.D. Standardization of nested-PCR for the detection of Pasteurella multocida, Staphylococcus aureus, myxomatosis virus, and rabbit haemorrhagic disease virus. In Proceedings of the 9th World Rabbit Congress, Verona, Italy, 10–13 June 2008; pp. 1035–1040. [Google Scholar]
- Gould, A.R.; Kattenbelt, J.A.; Lenghaus, C.; Morrissy, C.; Chamberlain, T.; Collins, B.J.; Westbury, H.A. The complete nucleotide sequence of rabbit haemorrhagic disease virus (Czech strain V351): Use of the polymerase chain reaction to detect replication in Australian vertebrates and analysis of viral population sequence variation. Virus Res. 1997, 47, 7–17. [Google Scholar] [CrossRef]
- Kwit, E.; Rzeżutka, A. Molecular methods in detection and epidemiologic studies of rabbit and hare viruses: A review. J. Vet. Diagn. Investig. 2019, 31, 497–508. [Google Scholar] [CrossRef]
- Polz, M.F.; Cavanaugh, C.M. Bias in template-to-product ratios in multitemplate PCR. Appl. Environ. Microbiol. 1998, 64, 3724–3730. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-T.; Zhang, J.; Sun, D.-H.; Ma, L.-N.; Liu, X.-T.; Cai, X.-P.; Liu, Y.-S. Development of reverse transcription loop-mediated isothermal amplification for rapid detection of H9 avian influenza virus. J. Virol. Methods 2008, 151, 200–203. [Google Scholar] [CrossRef]
- Peyrefitte, C.N.; Boubis, L.; Coudrier, D.; Bouloy, M.; Grandadam, M.; Tolou, H.J.; Plumet, S. Real-time reverse-transcription loop-mediated isothermal amplification for rapid detection of rift valley Fever virus. J. Clin. Microbiol. 2008, 46, 3653–3659. [Google Scholar] [CrossRef] [Green Version]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagamine, K.; Hase, T.; Notomi, T. Accelerated reaction by loop-mediated isothermal amplification using loop primers. Mol. Cell. Probes 2002, 16, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Guo, D.; Liu, J.; Si, C.; Jiang, Q.; Lin, H.; Yang, T.; Qu, L. Development of a reverse-transcription loop-mediated isothermal amplification method for detection of rabbit hemorrhagic disease virus. J. Virol. Methods 2013, 187, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Jansen, R.W.; Siegl, G.; Lemon, S.M. Molecular epidemiology of human hepatitis A virus defined by an antigen-capture polymerase chain reaction method. Proc. Natl. Acad. Sci. USA 1990, 87, 2867–2871. [Google Scholar] [CrossRef] [Green Version]
- Le Gall-Reculé, G.; Zwingelstein, F.; Portejoie, Y.; Le Gall, G. Immunocapture-RT-PCR assay for detection and molecular epidemiology studies of Rabbit Haemorrhagic Disease and European Brown Hare Syndrome viruses. J. Virol. Methods 2001, 97, 49–57. [Google Scholar] [CrossRef]
- Le Gall, G.; Arnauld, C.; Boilletot, E.; Morisse, J.P.; Rasschaert, D. Molecular epidemiology of rabbit haemorrhagic disease virus outbreaks in France during 1988 to 1995. J. Gen. Virol. 1998, 79, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowotny, N.; Bascuñana, C.R.; Ballagi-Pordány, A.; Gavier-Widén, D.; Uhlén, M.; Belák, S. Phylogenetic analysis of rabbit haemorrhagic disease and European brown hare syndrome viruses by comparison of sequences from the capsid protein gene. Arch. Virol. 1997, 142, 657–673. [Google Scholar] [CrossRef]
- Asgari, S.; Hardy, J.R.E.; Sinclair, R.G.; Cooke, B.D. Field evidence for mechanical transmission of rabbit haemorrhagic disease virus (RHDV) by flies (Diptera: Calliphoridae) among wild rabbits in Australia. Virus Res. 1998, 54, 123–132. [Google Scholar] [CrossRef]
- Shien, J.H.; Shieh, H.K.; Lee, L.H. Experimental infections of rabbits with rabbit haemorrhagic disease virus monitored by polymerase chain reaction. Res. Vet. Sci. 2000, 68, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Guittré, C.; Ruvoen-Clouet, N.; Barraud, L.; Cherel, Y.; Baginski, I.; Prave, M.; Ganiere, J.P.; Trépo, C.; Cova, L. Early stages of rabbit haemorrhagic disease virus infection monitored by polymerase chain reaction. Zentralblatt fur Veterinarmedizin. Reihe B 1996, 43, 109–118. [Google Scholar] [CrossRef]
- Kerr, P.J.; Kitchen, A.; Holmes, E.C. Origin and phylodynamics of rabbit hemorrhagic disease virus. J. Virol. 2009, 83, 12129–12138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvério, D.; Lopes, A.M.; Melo-Ferreira, J.; Monterroso, P.; Serronha, A.; Alves, P.C.; Magalhaes, M.; Esteves, P.J.; Abrantes, J. Insights into the evolution of the new variant rabbit hemorrhagic disease virus (GI.2) and the identification of novel recombinant strains. Transbound. Emerg. Dis. 2018, 65, 983–992. [Google Scholar] [CrossRef] [PubMed]
- Calvete, C.; Mendoza, M.; Alcaraz, A.; Sarto, M.P.; Jiménez-de-Bagüéss, M.P.; Calvo, A.J.; Monroy, F.; Calvo, J.H. Rabbit haemorrhagic disease: Cross-protection and comparative pathogenicity of GI.2/RHDV2/b and GI.1b/RHDV lagoviruses in a challenge trial. Vet. Microbiol. 2018, 219, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Dang, R.; Wang, X. Development of a SYBR-based real-time PCR to detect rabbit hemorrhagic disease virus (RHDV) and analyze its tissue distribution in experimentally infected rabbits. Virol. Sin. 2015, 30, 228–230. [Google Scholar] [CrossRef]
- Duarte, M.D.; Carvalho, C.L.; Barros, S.C.; Henriques, A.M.; Ramos, F.; Fagulha, T.; Luís, T.; Duarte, E.L.; Fevereiro, M. A real time Taqman RT-PCR for the detection of rabbit hemorrhagic disease virus 2 (RHDV2). J. Virol. Methods 2015, 219, 90–95. [Google Scholar] [CrossRef] [Green Version]
- Le Gall-Reculé, G.; Lemaitre, E.; Bertagnoli, S.; Hubert, C.; Top, S.; Decors, A.; Marchandeau, S.; Guitton, J.-S. Large-scale lagovirus disease outbreaks in European brown hares (Lepus europaeus) in France caused by RHDV2 strains spatially shared with rabbits (Oryctolagus cuniculus). Vet. Res. 2017, 48, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzner, A.; Niedbalski, W.I.; Kęsy, A.N.; Paprocka, G.R. Detection of RHD virus by a real-time reverse transcription PCR. Bull. Vet. Inst. Pulawy 2011, 55, 581–586. [Google Scholar]
- Niedźwiedzka-Rystwej, P.; Hukowska-Szematowicz, B.; Działo, J.; Tokarz-Deptuła, B.; Deptuła, W. Real time PCR detection of rabbit haemorrhagic disease virus in rabbits infected with different European strains of RHDV. Pol. J. Vet. Sci. 2013, 16, 39–43. [Google Scholar] [CrossRef] [Green Version]
- Strive, T.; Wright, J.; Kovaliski, J.; Botti, G.; Capucci, L. The non-pathogenic Australian lagovirus RCV-A1 causes a prolonged infection and elicits partial cross-protection to rabbit haemorrhagic disease virus. Virology 2010, 398, 125–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefever, S.; Pattyn, F.; Hellemans, J.; Vandesompele, J. Single-Nucleotide Polymorphisms and Other Mismatches Reduce Performance of Quantitative PCR Assays. Clin. Chem. 2013, 59, 1470–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthaei, M.; Kerr, P.J.; Read, A.J.; Hick, P.; Haboury, S.; Wright, J.D.; Strive, T. Comparative quantitative monitoring of rabbit haemorrhagic disease viruses in rabbit kittens. Virol. J. 2014, 11, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvete, C.; Mendoza, M.; Sarto, M.P.; Jiménez de Bagüés, M.; Luján, L.; Molín, J.; Calvo, A.J.; Monroy, F.; Calvo, J.H. Detection of Rabbit Hemorrhagic Disease Virus GI.2/RHDV2/b in the Mediterranean Pine Vole (Microtus duodecimcostatus) and White-Toothed Shrew (Crocidura russula). J. Wildl. Dis. 2019, 55, 467–472. [Google Scholar] [CrossRef]
- Elsworth, P.; Cooke, B.D.; Kovaliski, J.; Sinclair, R.; Holmes, E.C.; Strive, T. Increased virulence of rabbit haemorrhagic disease virus associated with genetic resistance in wild Australian rabbits (Oryctolagus cuniculus). Virology 2014, 464–465, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Hall, R.N.; Huang, N.; Roberts, J.; Strive, T. Carrion flies as sentinels for monitoring lagovirus activity in Australia. Transbound. Emerg. Dis. 2019, 66, 2025–2032. [Google Scholar] [CrossRef]
- Mahar, J.E.; Hall, R.N.; Shi, M.; Mourant, R.; Huang, N.; Strive, T.; Holmes, E.C. The discovery of three new hare lagoviruses reveals unexplored viral diversity in this genus. Virus Evol. 2019, 5, vez005. [Google Scholar] [CrossRef] [Green Version]
- Zinter, M.S.; Mayday, M.Y.; Ryckman, K.K.; Jelliffe-Pawlowski, L.L.; DeRisi, J.L. Towards precision quantification of contamination in metagenomic sequencing experiments. Microbiome 2019, 7, 62. [Google Scholar] [CrossRef] [PubMed]
- Eden, J.-S.; Kovaliski, J.; Duckworth, J.A.; Swain, G.; Mahar, J.E.; Strive, T.; Holmes, E.C. Comparative Phylodynamics of Rabbit Hemorrhagic Disease Virus in Australia and New Zealand. J. Virol. 2015, 89, 9548–9558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, R.N.; Capucci, L.; Matthaei, M.; Esposito, S.; Kerr, P.J.; Frese, M.; Strive, T. An in vivo system for directed experimental evolution of rabbit haemorrhagic disease virus. PLoS ONE 2017, 12, e0173727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Happi, A.N.; Ogunsanya, O.A.; Oguzie, J.U.; Oluniyi, P.E.; Olono, A.S.; Heeney, J.L.; Happi, C.T. Microbial Metagenomic Approach Uncovers the First Rabbit Haemorrhagic Disease Virus genome in Sub-Saharan Africa. bioRxiv 2020. [Google Scholar] [CrossRef]
- Bergin, I.L.; Wise, A.G.; Bolin, S.R.; Mullaney, T.P.; Kiupel, M.; Maes, R.K. Novel calicivirus identified in rabbits, Michigan, USA. Emerg. Infect. Dis. 2009, 15, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, E.; Zwingelstein, F.; Marchandeau, S.; Le Gall-Reculé, G. First complete genome sequence of a European non-pathogenic rabbit calicivirus (lagovirus GI.3). Arch. Virol. 2018, 163, 2921–2924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, A.M.; Silvério, D.; Magalhães, M.J.; Areal, H.; Alves, P.C.; Esteves, P.J.; Abrantes, J. Characterization of old RHDV strains by complete genome sequencing identifies a novel genetic group. Sci. Rep. 2017, 7, 13599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szillat, K.P.; Höper, D.; Beer, M.; König, P. Full-genome sequencing of German rabbit haemorrhagic disease virus uncovers recombination between RHDV (GI.2) and EBHSV (GII.1). Virus Evol. 2020, 6. [Google Scholar] [CrossRef]
- Dalton, K.P.; Nicieza, I.; Podadera, A.; de Llano, D.; Martin Alonso, J.M.; de los Toyos, J.R.; García Ocaña, M.; Vázquez-Villa, F.; Velasco, B.; Landeta, O.; et al. Fast specific field detection of RHDVb. Transbound. Emerg. Dis. 2018, 65, 232–234. [Google Scholar] [CrossRef] [Green Version]
- Bębnowska, D.; Niedźwiedzka-Rystwej, P. Characteristics of a new variant of rabbit haemorrhagic disease virus—RHDV2. Acta Biol. 2019, 26, 83–97. [Google Scholar] [CrossRef]
- Wu, M.; Zhu, Y.; Cong, F.; Rao, D.; Yuan, W.; Wang, J.; Huang, B.; Lian, Y.; Zhang, Y.; Huang, R.; et al. Rapid detection of three rabbit pathogens by use of the Luminex x-TAG assay. BMC Vet. Res. 2018, 14, 127. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Yuan, W.; Zhang, T.; Zhu, Y.; Lian, Y.; Zhang, Y.; Huang, R.; Guo, P. Simultaneous detection of 4 prototypic rat parvoviruses using the luminex xTAG assay in laboratory animal health monitoring. J. Virol. Methods 2017, 248, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Ma, L.; Cong, F.; Zhu, Y.; Xu, F.; Lian, Y.; Huang, B.; Xiao, L.; Chen, M.; Zhang, Y.; et al. High-throughput Luminex xMAP assay for simultaneous detection of antibodies against rabbit hemorrhagic disease virus, Sendai virus and rabbit rotavirus. Arch. Virol. 2019, 164, 1639–1646. [Google Scholar] [CrossRef] [PubMed]
- Peshev, R.; Alexandrov, M.; Ivanov, Y.; Gogov, Y. Demonstration of rabbit haemorrhagic disease virus antigen by Staphylococcus protein a coagglutination test. J. Virol. Methods 1996, 57, 15–18. [Google Scholar] [CrossRef]
- Kronvall, G. A Rapid Slide-Agglutination Method for Typing Pneumococci by Means of Specific Antibody Adsorbed to Protein A-Containing Staphylococci. J. Med. Microbiol. 1973, 6, 187–190. [Google Scholar] [CrossRef] [Green Version]
- Capucci, L.; Cavadini, P.; Schiavitto, M.; Lombardi, G.; Lavazza, A. Increased pathogenicity in rabbit haemorrhagic disease virus type 2 (RHDV2). Vet. Rec. 2017, 180, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strive, T.; Elsworth, P.; Liu, J.; Wright, J.D.; Kovaliski, J.; Capucci, L. The non-pathogenic Australian rabbit calicivirus RCV-A1 provides temporal and partial cross protection to lethal Rabbit Haemorrhagic Disease Virus infection which is not dependent on antibody titres. Vet. Res. 2013, 44, 51. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Type of Assay | Type of Sample | Detection Limit | Ability to Distinguish Variants and Genotypes | Application Scope |
|---|---|---|---|---|---|
| RHDV particles | HA | Liver, spleen, serum | n.d. | No | Virus detection |
| EM | Liver, lung, kidney, spleen | 105 viral particles/mL | No | Virus detection, localization, distribution, VLP correct self-assembly | |
| RHDV antigens | IHC | Liver 1 | 107–108 viral copies/mg | Yes 2,3 | Virus detection, localization, distribution |
| ELISA-Ag | Liver, spleen, blood, serum, heart | <HA | Yes 2,3 | Virus detection | |
| LFIA | Liver, abdominal liquid exsudate | 7.8 ng/mL | Yes 2 | Virus detection | |
| Sp A COAT | Liver, spleen, kidney, lung | <HA | No | Virus detection | |
| RHDV RNA | ISH | Liver, duodenum 4 | <IHC | Yes 2,3 | Virus detection, replication sites |
| RT-PCR | Liver, kidney, spleen, lung, heart, brain, bone marrow, muscle, spinal cord, thymus, lymph, nasal secretions, urine, feces, sera | 12 copies | Yes 2,3 | Virus detection, genome characterization 5 | |
| RT-qPCR | Same as RT-PCR | 9 copies | Yes 2,3 | Virus detection, genome characterization 5 | |
| NGS | Liver 1 | n.d. | Yes | Virus detection, genome characterization, virus differentiation | |
| Luminex xTAG | Liver 1 | 100 copies/µL | No | Virus detection | |
| RHDV capsid proteins | WB | Liver | n.d. | Yes 2,3 | Virus detection, validation of VLPs size |
| Anti-RHDV antibodies | WB | Serum | n.a. | No | Serosurveillance; mapping epitopes, confirming MAbs specificity and suitability |
| HIT | n.a. | No | Serosurveillance | ||
| ELISA-Ab | n.a. | Yes 2,3 | Serosurveillance | ||
| Luminex xMAP | n.a. | No | Serosurveillance |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abrantes, J.; Lopes, A.M. A Review on the Methods Used for the Detection and Diagnosis of Rabbit Hemorrhagic Disease Virus (RHDV). Microorganisms 2021, 9, 972. https://doi.org/10.3390/microorganisms9050972
Abrantes J, Lopes AM. A Review on the Methods Used for the Detection and Diagnosis of Rabbit Hemorrhagic Disease Virus (RHDV). Microorganisms. 2021; 9(5):972. https://doi.org/10.3390/microorganisms9050972
Chicago/Turabian StyleAbrantes, Joana, and Ana M. Lopes. 2021. "A Review on the Methods Used for the Detection and Diagnosis of Rabbit Hemorrhagic Disease Virus (RHDV)" Microorganisms 9, no. 5: 972. https://doi.org/10.3390/microorganisms9050972
APA StyleAbrantes, J., & Lopes, A. M. (2021). A Review on the Methods Used for the Detection and Diagnosis of Rabbit Hemorrhagic Disease Virus (RHDV). Microorganisms, 9(5), 972. https://doi.org/10.3390/microorganisms9050972