
Transposon Mutagenesis of Pseudomonas syringae Pathovars syringae and morsprunorum to Identify Genes Involved in Bacterial Canker Disease of Cherry

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Transposon Insertion Library Construction
2.3. Screening of Transposon Insertion Libraries
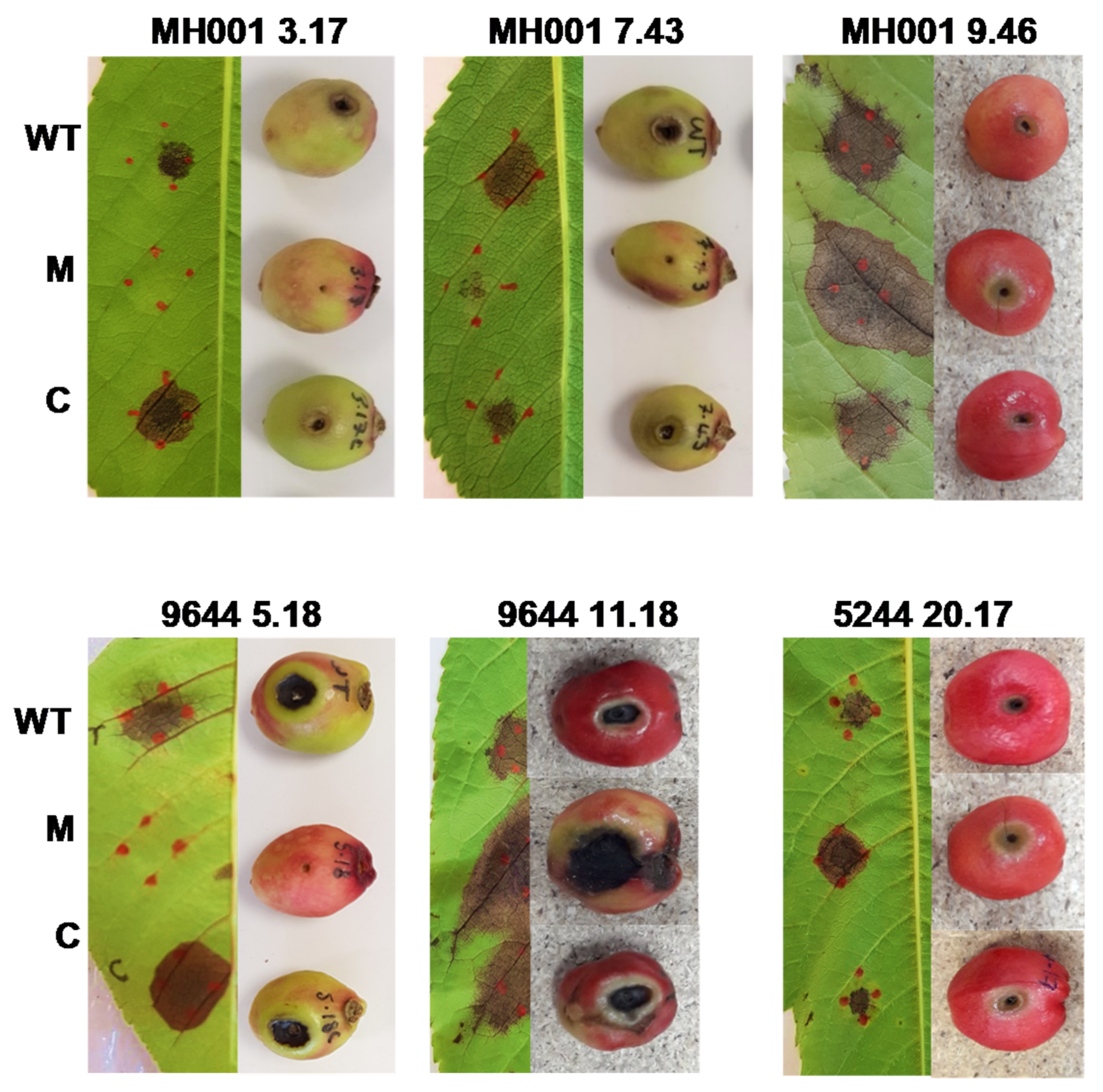
2.3.1. Pathogenicity Testing
2.3.2. Colony Morphology
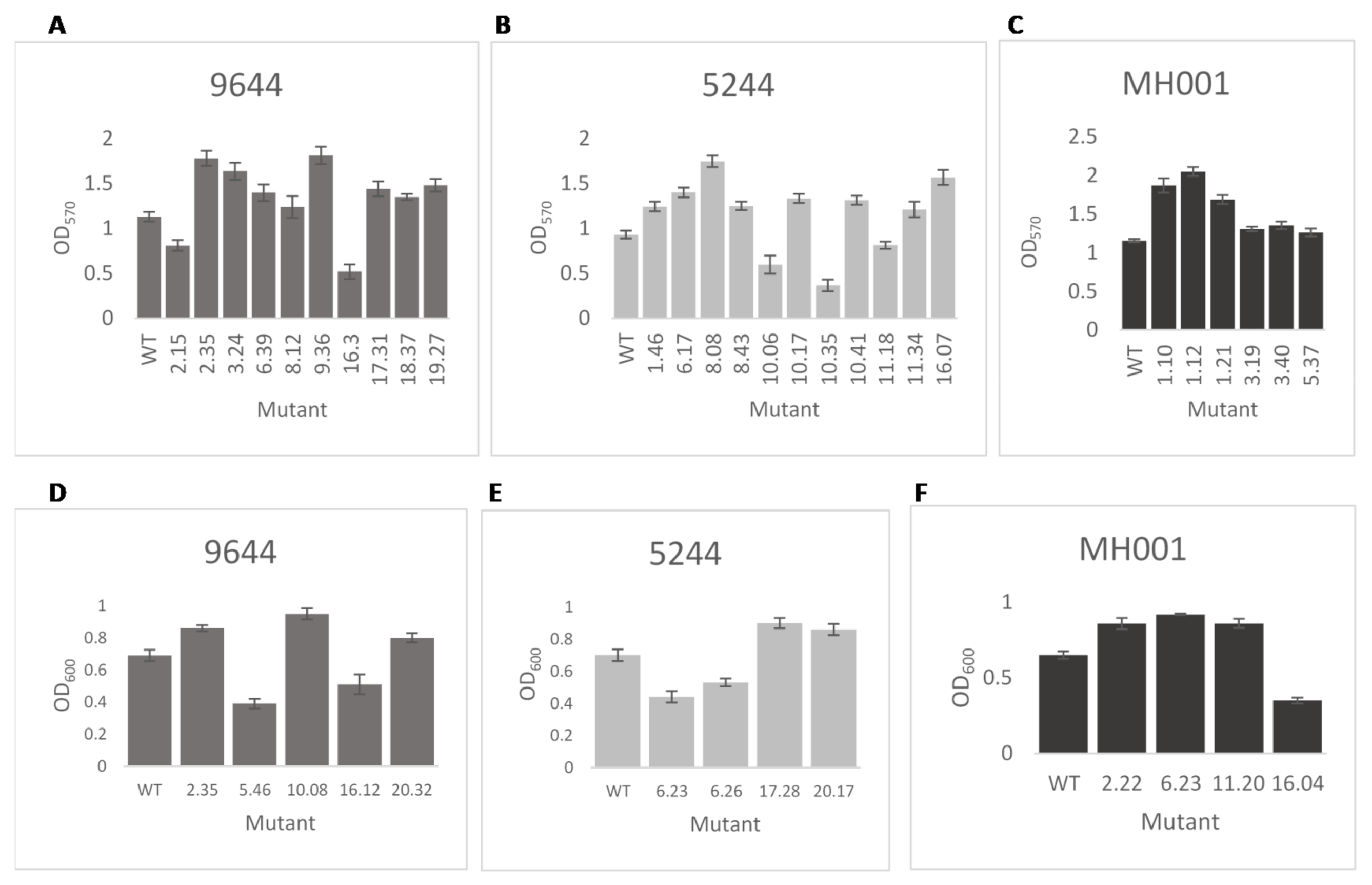
2.3.3. Biofilm Attachment
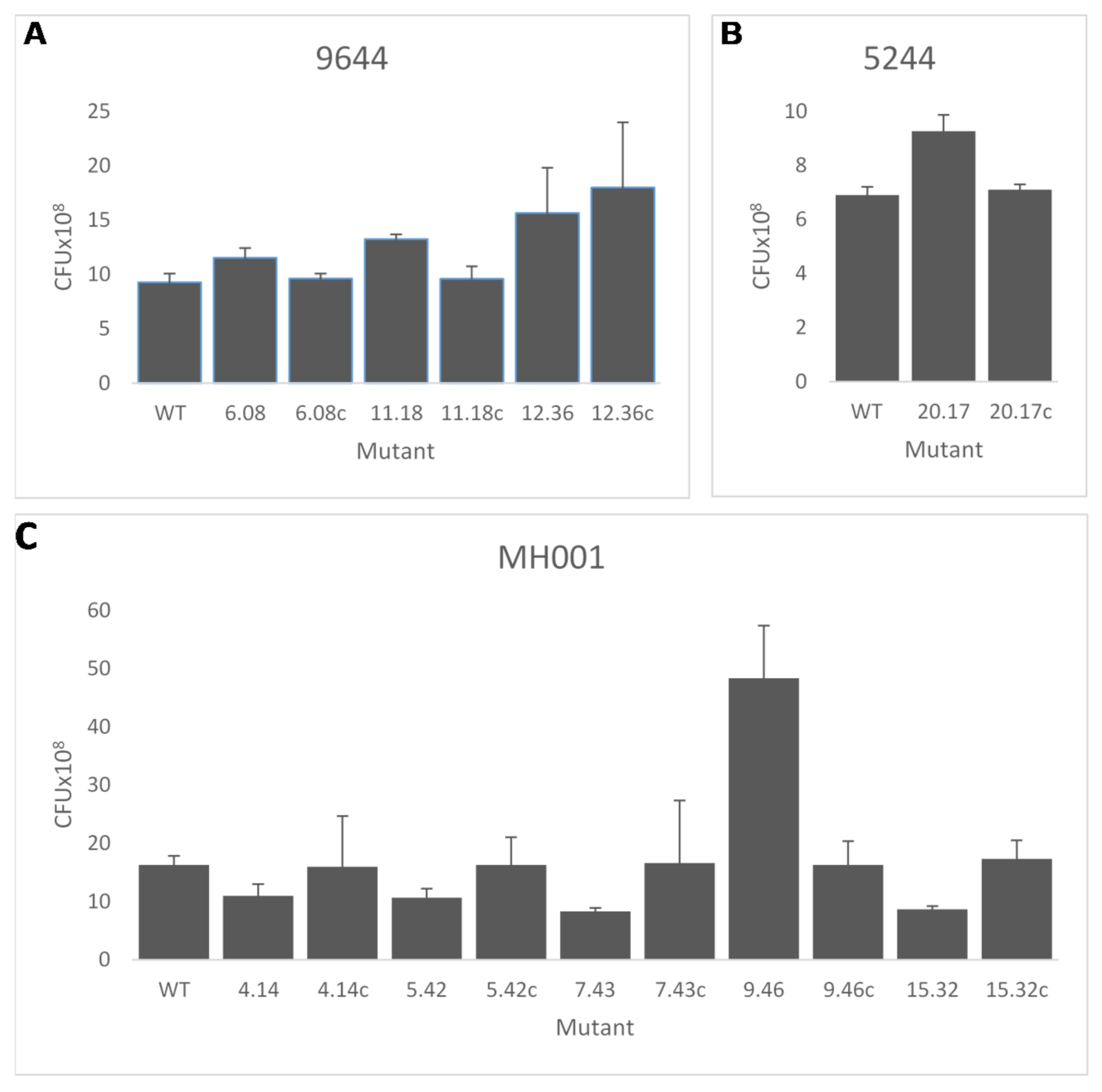
2.3.4. In Vitro Growth Rate
2.3.5. Swarming Ability
2.4. Cloning and Complementation
3. Results
3.1. Pathogenicity Screening of Pseudomonas syringae Transposon Mutants
3.2. In Vitro Screening of Transposon Mutants
3.3. Selection of Transposon Mutants for Further Analysis
3.4. Swarming Ability of Selected Transposon Mutants
3.5. Identification of Disrupted Genes in Selected Transposon Mutants
3.6. Complementation of Transposon Mutants
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hulin, M.T.; Armitage, A.D.; Vicente, J.G.; Holub, E.B.; Baxter, L.; Bates, H.J.; Mansfield, J.W.; Jackson, R.W.; Harrison, R.J. Comparative genomics of Pseudomonas syringae reveals convergent gene gain and loss associated with specialization onto cherry (Prunus avium). New Phytol. 2018, 219, 672–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spotts, R.A.; Wallis, K.M.; Serdani, M.; Azarenko, A.N. Bacterial Canker of Sweet Cherry in Oregon—Infection of Horticultural and Natural Wounds, and Resistance of Cultivar and Rootstock Combinations. Plant Dis. 2010, 94, 345–350. [Google Scholar] [CrossRef] [Green Version]
- Xin, X.-F.; Kvitko, B.; He, S.Y. Pseudomonas syringae: What it takes to be a pathogen. Nat. Rev. Genet. 2018, 16, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Crosse, J.E. Epidemiological Relations of the Pseudomonad Pathogens of Deciduous Fruit Trees. Annu. Rev. Phytopathol. 1966, 4, 291–310. [Google Scholar] [CrossRef]
- Hulin, M.T.; Jackson, R.W.; Harrison, R.J.; Mansfield, J.W. Cherry picking by pseudomonads: After a century of research on canker, genomics provides insights into the evolution of pathogenicity towards stone fruits. Plant Pathol. 2020, 69, 962–978. [Google Scholar] [CrossRef]
- Bender, C.L.; Alarcón-Chaidez, F.; Gross, D.C. Pseudomonas syringae phytotoxins: Mode of action, regulation, and biosynthesis by peptide and polyketide synthetases. Microbiol. Mol. Biol. Rev. 1999, 63, 266–292. [Google Scholar] [CrossRef] [Green Version]
- Mansfield, J.W. Antimicrobial Compounds. In Mechanisms of Resistance to Plant Diseases; Slusarenko, A.J., Frase, R.S.S., van Loon, L.C., Eds.; Kluwer Academic Press: Dordrecht, The Netherlands, 2000; pp. 325–370. [Google Scholar] [CrossRef]
- Alfano, J.R.; Collmer, A. Type III Secretion System Effector Proteins: Double Agents in Bacterial Disease and Plant Defense. Annu. Rev. Phytopathol. 2004, 42, 385–414. [Google Scholar] [CrossRef] [Green Version]
- Lindgren, P.B.; Panopoulos, N.J.; Staskawicz, B.J.; Dahlbeck, D. Genes required for pathogenicity and hypersensitivity are conserved and interchangeable among pathovars of Pseudomonas syringae. Mol. Genet. Genom. 1988, 211, 499–506. [Google Scholar] [CrossRef]
- Cuppels, D.A. Generation and Characterization of Tn5 Insertion Mutations in Pseudomonas syringae pv. tomato. Appl. Environ. Microbiol. 1986, 51, 323–327. [Google Scholar] [CrossRef] [Green Version]
- Bender, C.L.; Cooksey, D.A. Molecular cloning of copper resistance genes from Pseudomonas syringae pv. tomato. J. Bacteriol. 1987, 169, 470–474. [Google Scholar] [CrossRef] [Green Version]
- Sakata, N.; Ishiga, T.; Saito, H.; Nguyen, V.T.; Ishiga, Y. Transposon mutagenesis revealsPseudomonas cannabinapv.alisalensisoptimizes its virulence factors for pathogenicity on different hosts. PeerJ 2019, 7, e7698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manoharan, B.; Neale, H.C.; Jackson, R.W.; Hancock, J.T.; Arnold, D.L. Use of Pseudomonas syringae transposon libraries to identify genes involved in plant colonisation. PLoS ONE 2015, 10, e0137355. [Google Scholar] [CrossRef] [PubMed]
- van Opijnen, T.; Bodi, K.L.; Camilli, A. Tn-seq; high-throughput parallel sequencing for fitness and genetic interaction studies in microorganisms. Nat. Methods 2009, 6, 767–772. [Google Scholar] [CrossRef]
- Helmann, T.; Deutschbauer, A.; Lindow, S. Genome-wide identification of Pseudomonas syringae genes required for competitive fitness during colonization of the leaf surface and apoplast. Proc. Natl. Acad. Sci. USA 2019, 116, 18900–18910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hulin, M.T.; Mansfield, J.W.; Brain, P.; Xu, X.; Jackson, R.W.; Harrison, R.J. Characterization of the pathogenicity of strains of Pseudomonas syringae towards cherry and plum. Plant Pathol. 2018, 67, 1177–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giddens, S.R.; Jackson, R.W.; Moon, C.D.; Jacobs, M.A.; Zhang, X.-X.; Gehrig, S.M.; Rainey, P.B. Mutational activation of niche-specific genes provides insight into regulatory networks and bacterial function in a complex environment. Proc. Natl. Acad. Sci. USA 2007, 104, 18247–18252. [Google Scholar] [CrossRef] [Green Version]
- Kovach, M.E.; Elzer, P.H.; Hill, D.S.; Robertson, G.; Farris, M.A.; Roop, R.; Peterson, K.M. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 1995, 166, 175–176. [Google Scholar] [CrossRef]
- Spiers, A.J.; Bohannon, J.; Gehrig, S.M.; Rainey, P.B. Biofilm formation at the air-liquid interface by the Pseudomonas fluorescens SBW25 wrinkly spreader requires an acetylated form of cellulose. Mol. Microbiol. 2003, 50, 15–27. [Google Scholar] [CrossRef]
- Keen, N.T.; Shen, H.; Cooksey, D.A. Introduction of cloned DNA into plant pathogenic bacteria. In Molecular Plant Pathology: A Practical Approach; Gurr, S.J., McPherson, M.J., Bowles, D.J., Eds.; IRL Press: Oxford, UK, 1992. [Google Scholar]
- Xu, N.; Wang, L.; Cheng, H.; Liu, Q.; Liu, J.; Ma, Y. In vitro functional characterization of the Na+/H+ antiporters in Coryne-bacterium glutamicum. FEMS Microbiol. Lett. 2016, 363, fnv237. [Google Scholar] [CrossRef] [Green Version]
- Herz, K.; Vimont, S.; Padan, E.; Berche, P. Roles of NhaA, NhaB, and NhaD Na+ /H+ Antiporters in Survival of Vibrio cholerae in a Saline Environment. J. Bacteriol. 2003, 185, 1236–1244. [Google Scholar] [CrossRef] [Green Version]
- Xu, N.; Zheng, Y.; Wang, X.; Krulwich, T.A.; Ma, Y.; Liu, J. The Lysine 299 Residue Endows the Multisubunit Mrp1 Antiporter with Dominant Roles in Na+Resistance and pH Homeostasis in Corynebacterium glutamicum. Appl. Environ. Microbiol. 2018, 84, e00110-18. [Google Scholar] [CrossRef] [Green Version]
- Matas, I.M.; Lambertsen, L.; Rodríguez-Moreno, L.; Ramos, C. Identification of novel virulence genes and metabolic pathways required for full fitness of Pseudomonas savastanoi pv. savastanoi in olive (Olea europaea) knots. New Phytol. 2012, 196, 1182–1196. [Google Scholar] [CrossRef] [Green Version]
- Rico, A.; Preston, G.M. Pseudomonas syringae pv. tomato DC3000 Uses Constitutive and Apoplast-Induced Nutrient Assimilation Pathways to Catabolize Nutrients That Are Abundant in the Tomato Apoplast. Mol. Plant Microbe Interact. 2008, 21, 269–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.; Jayaraman, J.; Segonzac, C.; Park, H.-J.; Park, H.; Han, S.-W.; Sohn, K.H. Pseudomonas syringae pv. actinidiae Type III Effectors Localized at Multiple Cellular Compartments Activate or Suppress Innate Immune Responses in Nicotiana benthamiana. Front. Plant Sci. 2017, 8, 2157. [Google Scholar] [CrossRef] [PubMed]
- Macho, A.P.; Zumaquero, A.; Plaza, J.J.G.; Ortiz-Martín, I.; Rufián, J.S.; Beuzón, C.R. Genetic Analysis of the Individual Contribution to Virulence of the Type III Effector Inventory of Pseudomonas syringae pv. phaseolicola. PLoS ONE 2012, 7, e35871. [Google Scholar] [CrossRef] [Green Version]
- Huguet, E.; Hahn, K.; Wengelnik, K.; Bonas, U. hpaA mutants of Xanthomonas campestris pv. vesicatoriaare affected in path-ogenicity but retain the ability to induce host-specific hypersensitive reaction. Mol. Microbiol. 1998, 29, 1379–1390. [Google Scholar] [CrossRef]
- Morris, V.L.; Jackson, D.P.; Grattan, M.; Ainsworth, T.; Cuppels, D.A. Isolation and sequence analysis of the Pseudomonas syringae pv. tomato gene encoding a 2,3-diphosphoglycerate-independent phosphoglyceromutase. J. Bacteriol. 1995, 177, 1727–1733. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, N.; Wolfgang, M.C.; Goodman, A.L.; Arora, S.K.; Jyot, J.; Lory, S.; Ramphal, R. A four-tiered transcriptional regulatory circuit controls flagellar biogenesis in Pseudomonas aeruginosa. Mol. Microbiol. 2003, 50, 809–824. [Google Scholar] [CrossRef] [Green Version]
- Jarrell, K.F.; McBride, M.J. The surprisingly diverse ways that prokaryotes move. Nat. Rev. Microbiol. 2008, 6, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Duan, Q.; Zhou, M.; Zhu, L.; Zhu, G. Flagella and bacterial pathogenicity. J. Basic Microbiol. 2012, 53, 1–8. [Google Scholar] [CrossRef]
- Haiko, J.; Westerlund-Wikström, B. The Role of the Bacterial Flagellum in Adhesion and Virulence. Biology 2013, 2, 1242–1267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, N.A.; Urban, M.; Hammond-Kosack, K.E. The trans-kingdom identification of negative regulators of pathogen hy-pervirulence. FEMS Microbiol. Rev. 2015, 40, 19–40. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Description | Reference |
|---|---|---|
| Pseudomonas | ||
| Psm 5244 | NFR. Race 1 | [16] |
| Psm MH001 | NFR. Race 2 | [16] |
| Pss 9644 | RifR. Race 1 | [16] |
| E. coli | ||
| pCR2.1 TOP10F | Competent cells | Invitrogen, UK |
| S17-1 λpir | KMR. Containing plasmid pSCR001 | [17] |
| Plasmids | ||
| pBBR1MCS-5 | GmR. Broad host range cloning vector | [18] |
| pSCR001 | KmR. Carries IS-Ω-Km/hah transposon | [17] |
| pCR2.1 | KmR, AmpR. Cloning vector | Invitrogen, UK |
| Strain | Pathogenicity Phenotype Compared to WT (+) | |||||
| -- | - | ++ | +++ | |||
| 9644 | 7 | 30 | 25 | 14 | ||
| MH001 | 11 | 22 | 53 | 27 | ||
| 5244 | 25 | 8 | 17 | 3 | ||
| Morphology | ||||||
| Larger | Pathogenicity | Smaller | Pathogenicity | Irregular Shape | Pathogenicity | |
| 9644 | 24 | 1 reduced 4 increased | 34 | 9 reduced 2 increased | 14 | 4 reduced 1 increased |
| MH001 | 14 | 9 increased | 15 | 1 reduced | 0 | - |
| 5244 | 32 | 6 increased | 37 | 20 reduced | 5 | 3 reduced |
| Biofilm | ||||||
| Increased | Pathogenicity | Decreased | Pathogenicity | |||
| 9644 | 9 | 1 reduced 4 increased | 2 | 2 reduced | ||
| MH001 | 6 | 2 reduced 4 increased | 0 | - | ||
| 5244 | 8 | 3 reduced 5 increased | 3 | 2 increased | ||
| In Vitro Growth | ||||||
| Increased | Pathogenicity | Decreased | Pathogenicity | |||
| 9644 | 3 | 2 increased | 2 | 2 reduced | ||
| MH001 | 3 | 3 increased | 1 | 1 reduced | ||
| 5244 | 2 | 2 increased | 2 | 2 reduced | ||
| Mutant Number | Symptoms 1 | Swarming % WT | Gene Name/ Description | Biological Process | Locus Tag |
|---|---|---|---|---|---|
| Pss 9644 | |||||
| 1.38 | +++ | WT | Adenine DNA glycosylase | Base excision repair | BKM12_20200 |
| 3.15 | - | 92 ± 3 | Serine hydroxymethyltransferase 2 | Amino acid biosynthesis | BKM12_25860 |
| 5.18 | - | WT | Glycosyl transferase family 1 | N-glycan biosynthesis | BKM12_16730 |
| 6.08 | +++ | WT | Pyridoxine/pyridoxamine 5′-phosphate oxidase | Vitamin B6 biosynthesis | BKM12_05440 |
| 11.18 | +++ | WT | Na(+)/H(+) antiporter, nhaA | Sodium ion transport | BKM12_00120 |
| 12.36 | +++ | WT | Arginine-ornithine antiporter, argE | Arginine deiminase pathway | BKM12_00245 |
| Psm R1 5244 | |||||
| 20.17 | +++ | 65 ± 6 | Acetylornithine deacetylase | Arginine biosynthesis | BKM19_RS02785 |
| Psm R2 MH001 | |||||
| 3.17 | - | WT | Molecular chaperone, dnaJ | DNA replication | BKM03_RS28190 |
| 3.27 | - | WT | MFS transporter | Transmembrane transporter activity | BKM03_RS09500 |
| 4.02 | - | WT | Ornithine carbamoyltransferase | Arginine biosynthesis | BKM03_RS22585 |
| 4.14 | - | 87 ± 2 | Phosphoglycerate mutase, 2,3-bisphosphoglycerate | Glycolysis pathway | BKM03_RS28005 |
| 5.42 | - | 78 ± 5 | DNA ligase D | DNA replication | BKM03_RS11910 |
| 6.37 | - | WT | Beta-glucosidase | Cellulose catabolic process | BKM03_RS12410 |
| 7.43 | - | WT | T3SS protein, hrcS | Bacterial secretion system | BKM03_RS07835 |
| 8.11 | - | WT | Cu+ exporting protein | Metal ion binding | BKM03_RS10500 |
| 9.16 | - | WT | Acetolactate synthase | Amino acid biosynthesis | BKM03_RS05770 |
| 9.46 | ++ | WT | Type III effector, hopAU1 | Virulence | BKM03_RS30415 |
| 15.32 | - | 32 ± 3 | Flagellar hook-length protein, fliK | Flagellar assembly | BKM03_RS07850 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neale, H.C.; Hulin, M.T.; Harrison, R.J.; Jackson, R.W.; Arnold, D.L. Transposon Mutagenesis of Pseudomonas syringae Pathovars syringae and morsprunorum to Identify Genes Involved in Bacterial Canker Disease of Cherry. Microorganisms 2021, 9, 1328. https://doi.org/10.3390/microorganisms9061328
Neale HC, Hulin MT, Harrison RJ, Jackson RW, Arnold DL. Transposon Mutagenesis of Pseudomonas syringae Pathovars syringae and morsprunorum to Identify Genes Involved in Bacterial Canker Disease of Cherry. Microorganisms. 2021; 9(6):1328. https://doi.org/10.3390/microorganisms9061328
Chicago/Turabian StyleNeale, Helen C., Michelle T. Hulin, Richard J. Harrison, Robert W. Jackson, and Dawn L. Arnold. 2021. "Transposon Mutagenesis of Pseudomonas syringae Pathovars syringae and morsprunorum to Identify Genes Involved in Bacterial Canker Disease of Cherry" Microorganisms 9, no. 6: 1328. https://doi.org/10.3390/microorganisms9061328
APA StyleNeale, H. C., Hulin, M. T., Harrison, R. J., Jackson, R. W., & Arnold, D. L. (2021). Transposon Mutagenesis of Pseudomonas syringae Pathovars syringae and morsprunorum to Identify Genes Involved in Bacterial Canker Disease of Cherry. Microorganisms, 9(6), 1328. https://doi.org/10.3390/microorganisms9061328