
Absence of 4-Formylaminooxyvinylglycine Production by Pseudomonas fluorescens WH6 Results in Resource Reallocation from Secondary Metabolite Production to Rhizocompetence


Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Maintenance and Chemicals
2.2. RNA Isolation, RNAseq Library Preparation and Sequencing
2.3. Differential Expression Analysis
2.4. Genome Functional Annotation and Ortholog Identification
2.5. Quantitative Reverse-Transcriptase PCR
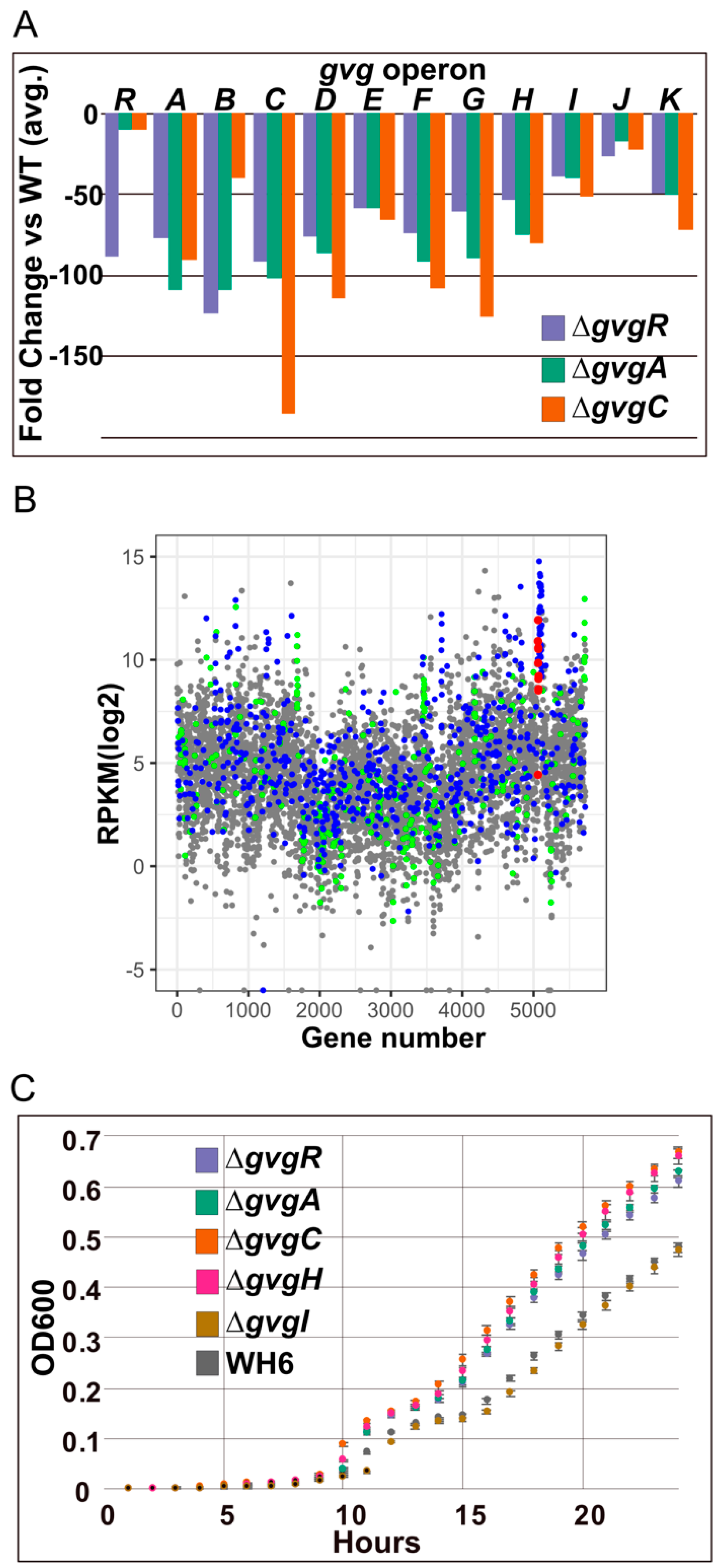
2.6. Motility and Biofilm Formation Assays
2.7. FVG-Inhibition Assay
3. Results
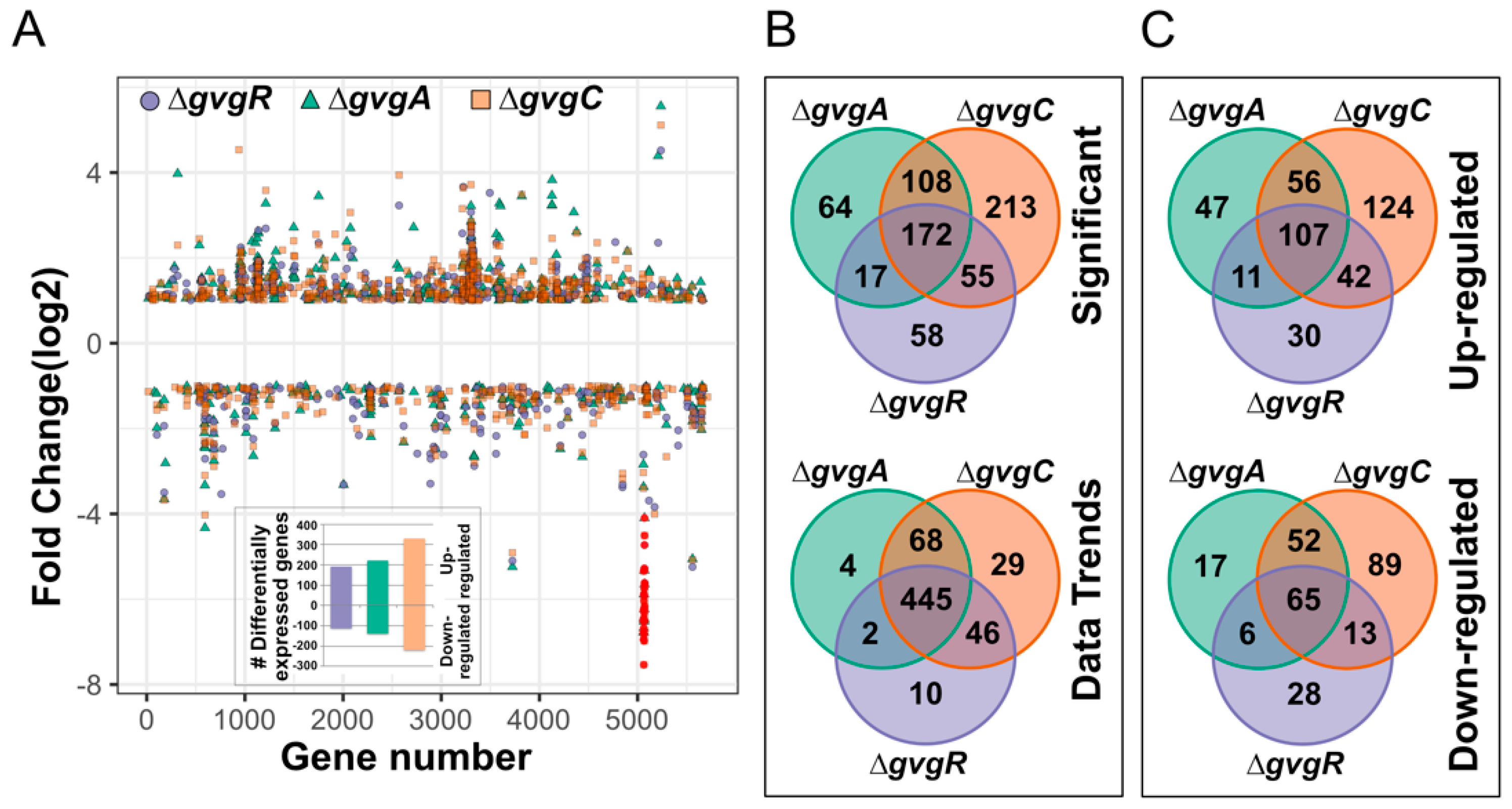
3.1. Absence of gvgR, A and C Result in Decreased Expression of the gvg Cluster and Large Transcriptome Changes Compared to Wild Type
3.1.1. Transcriptome Differences in ΔgvgR, C and A Strains Compared to Wild Type
3.1.2. gvg Cluster Expression in gvg Gene Knockout Strains
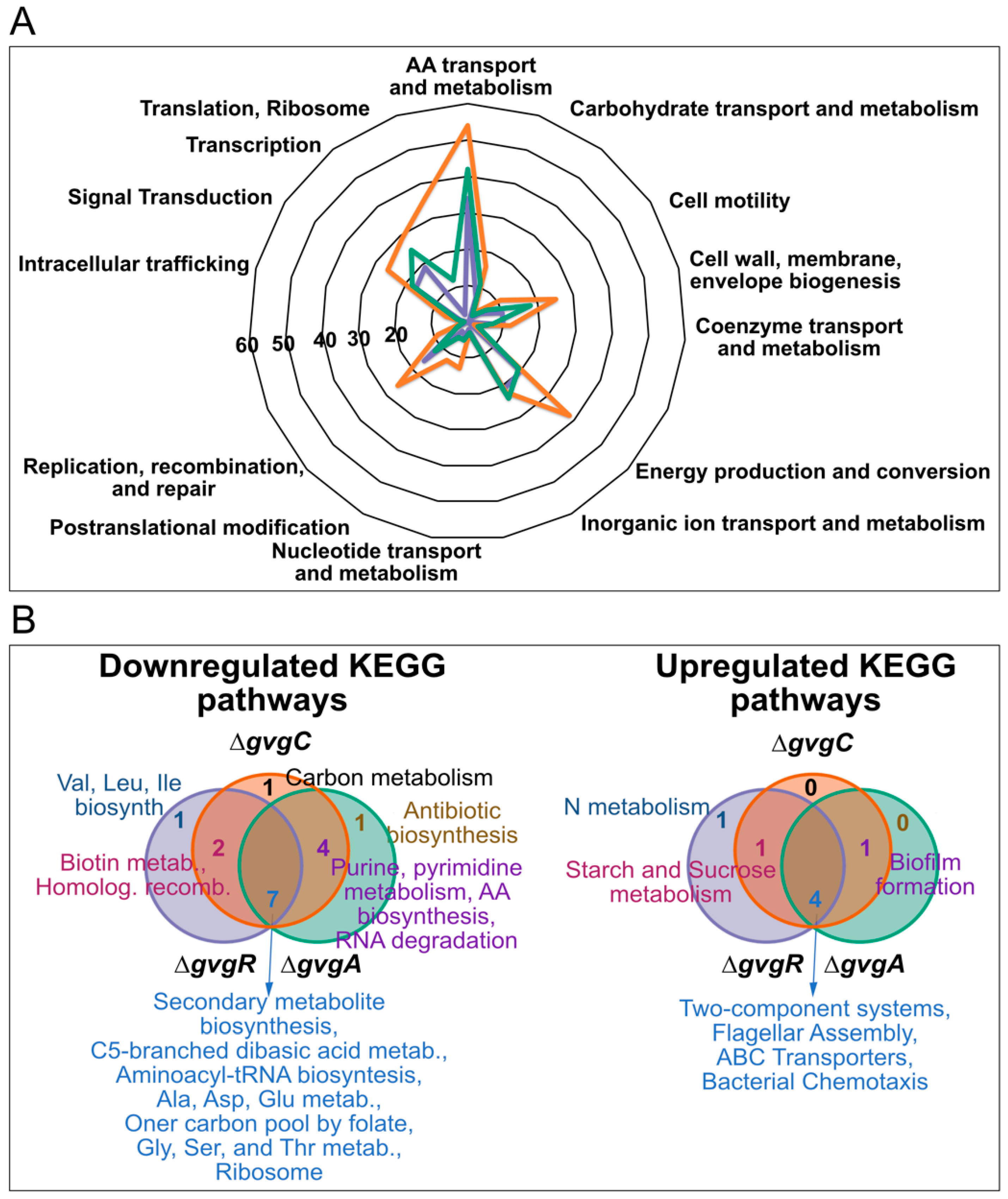
3.2. Functional Analysis of Transcriptome Shifts in ΔgvgR, A, and C Mutants Reveals a Role for Acidic Amino Acids and Their Amides in FVG Production
3.2.1. Functional Analyses of Genes Regulated in gvg Cluster Mutants
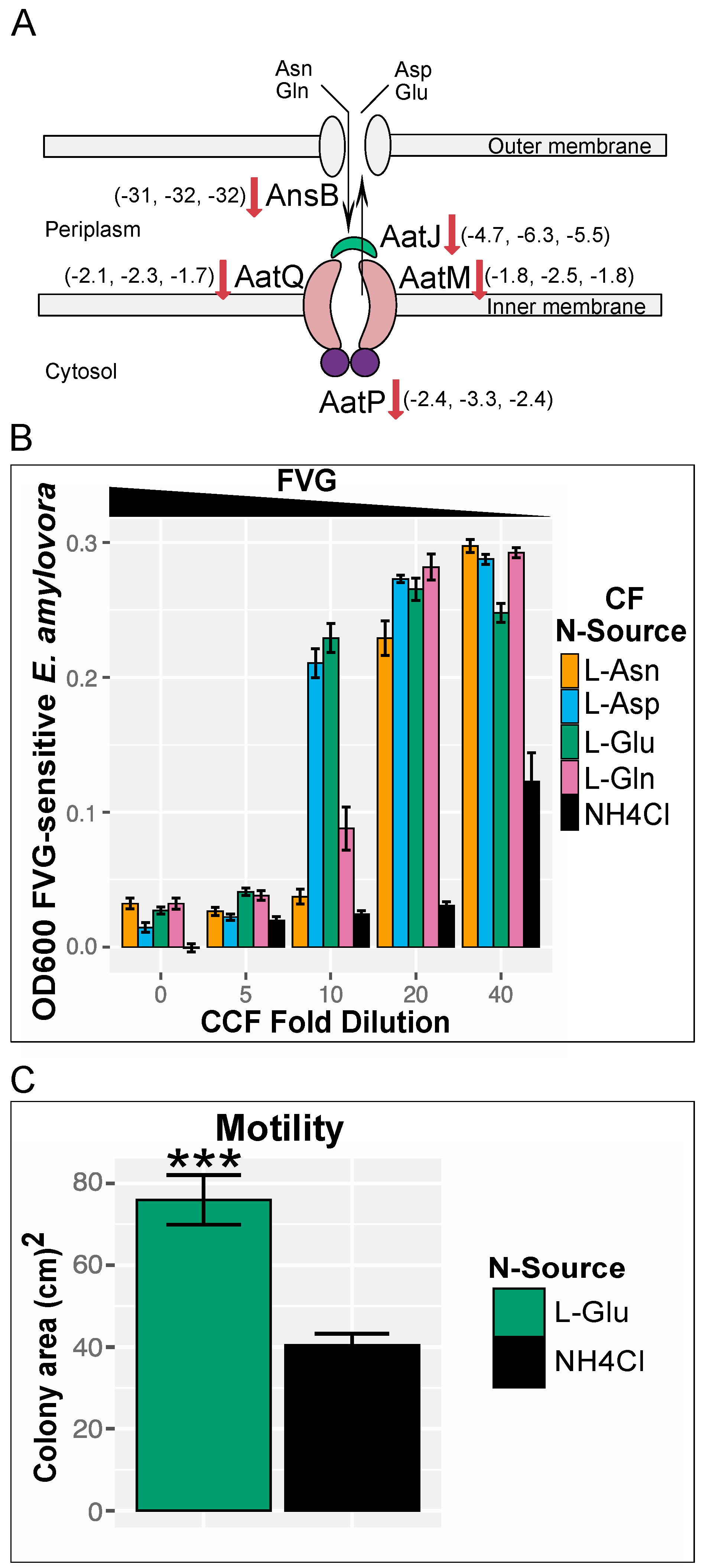
3.2.2. Transportome Changes in gvg Cluster Mutants
4. Discussion
4.1. Regulation of the gvg Operon
4.2. The Transcriptome of gvg Cluster Mutants Reveals Involvement of Acidic Amino Acids and Their Amides in the Regulation of FVG Production
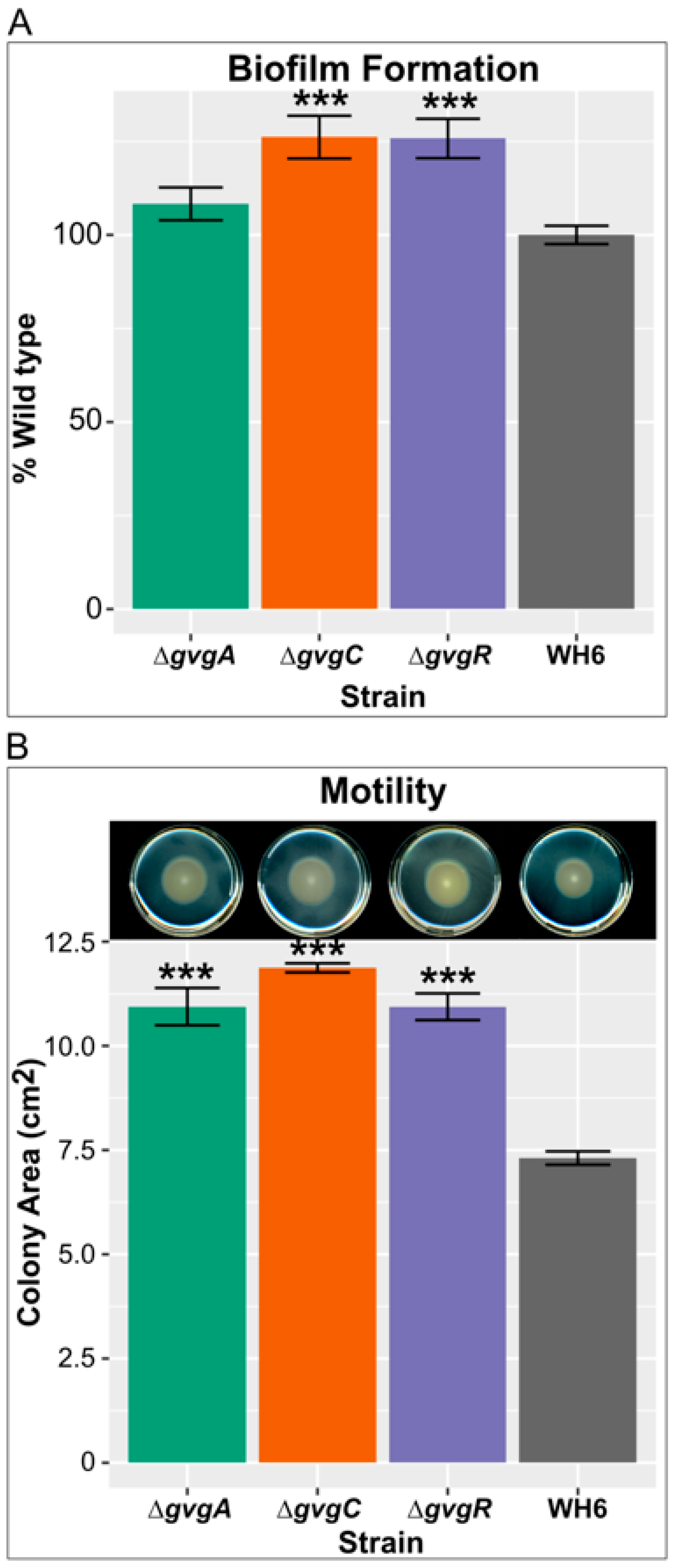
4.3. Genes Involved in Rhizocompetence Are Upregulated in the Absence of FVG
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keswani, C.; Singh, H.B.; García-Estrada, C.; Caradus, J.; He, Y.W.; Mezaache-Aichour, S.; Glare, T.R.; Borriss, R.; Sansinenea, E. Antimicrobial secondary metabolites from agriculturally important bacteria as next-generation pesticides. Appl. Microbiol. Biotechnol. 2020, 104, 1013–1034. [Google Scholar] [CrossRef] [PubMed]
- Gross, H.; Loper, J.E. Genomics of secondary metabolite production by Pseudomonas spp. Nat. Prod. Rep. 2009, 26, 1408–1446. [Google Scholar] [CrossRef]
- Elliott, L.F.; Azevedo, M.D.; Mueller-Warrant, G.W.; Horwath, W.R. Weed control with rhizobacteria. Soil Sci. Agrochem. Ecol. 1998, 33, 3–7. [Google Scholar]
- Shinde, S.; Cumming, J.R.; Collart, F.R.; Noirot, P.H.; Larsen, P.E. Pseudomonas fluorescens transportome is linked to strain-specific plant growth promotion in aspen seedlings under nutrient stress. Front. Plant Sci. 2017, 8, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halgren, A.; Azevedo, M.; Mills, D.; Armstrong, D.; Thimmaiah, M.; Mcphail, K.; Banowetz, G.M. Selective inhibition of Erwinia amylovora by the herbicidally active germination-arrest factor (GAF) produced by Pseudomonas bacteria. J. Appl. Microbiol. 2011, 111, 949–959. [Google Scholar] [CrossRef] [PubMed]
- McPhail, K.L.; Armstrong, D.J.; Azevedo, M.D.; Banowetz, G.M.; Mills, D.I. 4-formylaminooxyvinylglycine, an herbicidal germination-arrest factor from Pseudomonas rhizosphere bacteria. J. Nat. Prod. 2010, 73, 1853–1857. [Google Scholar] [CrossRef] [Green Version]
- Banowetz, G.M.; Azevedo, M.D.; Armstrong, D.J.; Halgren, A.B.; Mills, D.I. Germination-Arrest Factor (GAF): Biological properties of a novel, naturally-occurring herbicide produced by selected isolates of rhizosphere bacteria. Biol. Control 2008, 46, 380–390. [Google Scholar] [CrossRef]
- Okrent, R.A.; Trippe, K.M.; Manning, V.A.; Walsh, C.M. Detection of 4-formylaminooxyvinylglycine in culture filtrates of Pseudomonas fluorescens WH6 and Pantoea ananatis BRT175 by laser ablation electrospray ionization-mass spectrometry. PLoS ONE 2018, 13, e0200481. [Google Scholar] [CrossRef]
- Halgren, A.; Maselko, M.; Azevedo, M.; Mills, D.; Armstrong, D.; Banowetz, G. Genetics of germination-arrest factor (GAF) production by Pseudomonas fluorescens WH6: Identification of a gene cluster essential for GAF biosynthesis. Microbiology 2013, 159, 36–45. [Google Scholar] [CrossRef] [Green Version]
- Okrent, R.A.; Trippe, K.M.; Maselko, M.; Manning, V. Functional analysis of a biosynthetic cluster essential for production of 4-formylaminooxyvinylglycine, a germination-arrest factor from Pseudomonas fluorescens WH6. Microbiology 2017, 163, 207–217. [Google Scholar] [CrossRef]
- van Rij, E.T.; Girard, G.; Lugtenberg, B.J.J.; Bloemberg, G.V. Environmental and Molecular Regulation of Phenazine-1-carboxamide Biosynthesis in in Pseudomonas chlororaphis Strain PCL1391. Ph.D. Thesis, Leiden University, Leiden, The Netherlands, 2006. [Google Scholar]
- Tramonti, A.; Nardella, C.; di Salvo, M.L.; Pascarella, S.; Contestabile, R. The MocR-like transcription factors: Pyridoxal 5′-phosphate-dependent regulators of bacterial metabolism. FEBS J. 2018, 285, 3925–3944. [Google Scholar] [CrossRef] [Green Version]
- Berkowitz, D.B.; Charette, B.D.; Karukurichi, K.R.; McFadden, J.M. α-Vinylic amino acids: Occurrence, asymmetric synthesis, and biochemical mechanisms. Tetrahedron Asymmetry 2006, 17, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Okrent, R.A.; Halgren, A.B.; Azevedo, M.D.; Chang, J.H.; Mills, D.I.; Maselko, M.; Armstrong, D.J.; Banowetz, G.M.; Trippe, K.M. Negative regulation of germination-arrest factor production in Pseudomonas fluorescens WH6 by a putative extracytoplasmic function sigma factor. Microbiology 2014, 160, 2432–2442. [Google Scholar] [CrossRef] [Green Version]
- Burger, M.; Woods, R.G.; McCarthy, C.; Beacham, I.R. Temperature regulation of protease in Pseudomonas fluorescens LS 107d2 by an ECF sigma factor and a transmembrane activator. Microbiology 2000, 146, 3149–3155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 30 March 2021).
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Conesa, A.; Gotz, S.; García-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2go: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winsor, G.L.; Griffiths, E.J.; Lo, R.; Dhillon, B.K.; Shay, J.A.; Brinkman, F.S. Enhanced annotations and features for comparing thousands of Pseudomonas genomes in the Pseudomonas genome database. Nucleic Acids Res. 2016, 44, D646–D653. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Sato, Y.; Morishima, K. BlastKOALA and GhostKOALA: KEGG Tools for Functional Characterization of Genome and Metagenome Sequences. J. Mol. Biol. 2016, 428, 726–731. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Friedman, M.S.; Shedden, K.; Hankenson, K.D.; Woolf, P.J. GAGE: generally applicable gene set enrichment for pathway analysis. BMC Bioinformatics 2009, 10, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huerta-Cepas, J.; Szklarczyk, D.; Forslund, K.; Cook, H.; Heller, D.; Walter, M.C.; Rattei, T.; Mende, D.R.; Sunagawa, S.; Kuhn, M.; et al. EGGNOG 4.5: A hierarchical orthology framework with improved functional annotations for eukaryotic, prokaryotic and viral sequences. Nucleic Acids Res. 2016, 44, D286–D293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, G.A. Microtiter dish Biofilm formation assay. J. Vis. Exp. 2010, 47, 2437. [Google Scholar] [CrossRef]
- Härtig, E.; Schiek, U.; Vollack, K.U.; Zumft, W.G. Nitrate and nitrite control of respiratory nitrate reduction in denitrifying Pseudomonas stutzeri by a two-component regulatory system homologous to NarXL of Escherichia coli. J. Bacteriol. 1999, 181, 3658–3665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benkert, B.; Quäck, N.; Schreiber, K.; Jaensch, L.; Jahn, D.; Schobert, M. Nitrate-responsive NarX-NarL represses arginine-mediated induction of the Pseudomonas aeruginosa arginine fermentation arcDABC operon. Microbiology 2008, 154, 3053–3060. [Google Scholar] [CrossRef] [Green Version]
- Hasona, A.; Self, W.T.; Shanmugam, K.T. Transcriptional regulation of the moe (molybdate metabolism) operon of Escherichia coli. Arch. Microbiol. 2001, 175, 178–188. [Google Scholar] [CrossRef]
- Redondo-Nieto, M.; Barret, M.; Morrissey, J.; Germaine, K.; Martínez-Granero, F.; Barahona, E.; Navazo, A.; Sánchez-Contreras, M.; Moynihan, J.A.; Muriel, C.; et al. Genome sequence reveals that Pseudomonas fluorescens F113 possesses a large and diverse array of systems for rhizosphere function and host interaction. BMC Genom. 2013, 14, 54. [Google Scholar] [CrossRef] [Green Version]
- Zumft, W.G. Cell biology and molecular basis of denitrification. Microbiol. Mol. Biol. Rev. 1997, 61, 533–616. [Google Scholar] [CrossRef]
- Hay, I.D.; Wang, Y.; Moradali, M.F.; Rehman, Z.U.; Rehm, B.H.A. Genetics and regulation of bacterial alginate production. Environ. Microbiol. 2014, 16, 2997–3011. [Google Scholar] [CrossRef]
- Mann, E.E.; Wozniak, D.J. Pseudomonas biofilm matrix composition and niche biology. FEMS Microbiol. Rev. 2012, 36, 893–916. [Google Scholar] [CrossRef] [Green Version]
- Hay, I.D.; Rehman, Z.U.; Moradali, M.F.; Wang, Y.; Rehm, B.H.A. Microbial alginate production, modification and its applications. Microb. Biotechnol. 2013, 6, 637–650. [Google Scholar] [CrossRef] [Green Version]
- Ritchings, B.W.; Almira, E.C.; Lory, S.; Ramphal, R. Cloning and phenotypic characterization of fleS and fleR, new response regulators of Pseudomonas aeruginosa which regulate motility and adhesion to mucin. Infect. Immun. 1995, 63, 4868–4876. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, N.; Wolfgang, M.C.; Goodman, A.L.; Arora, S.K.; Jyot, J.; Lory, S.; Ramphal, R. A four-tiered transcriptional regulatory circuit controls flagellar biogenesis in Pseudomonas aeruginosa. Mol. Microbiol. 2003, 50, 809–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tart, A.H.; Blanks, M.J.; Wozniak, D.J. The AlgT-dependent transcriptional regulator AmrZ (AlgZ) inhibits flagellum biosynthesis in mucoid, nonmotile Pseudomonas aeruginosa cystic fibrosis isolates. J. Bacteriol. 2006, 188, 6483–6489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hüser, A.; Klöppner, U.; Röhm, K.H. Cloning, sequence analysis, and expression of ansB from Pseudomonas fluorescens, encoding periplasmic glutaminase/asparaginase. FEMS Microbiol. Lett. 1999, 178, 327–335. [Google Scholar] [CrossRef]
- Singh, B.; Röhm, K.H. Characterization of a Pseudomonas putida ABC transporter (AatJMQP) required for acidic amino acid uptake: Biochemical properties and regulation by the Aau two-component system. Microbiology 2008, 154, 797–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, K.B. Effect of Antagonistic Bacteria on Establishment of Honey Bee-Dispersed Erwinia amylovora in Pear Blossoms and on Fire Blight Control. Phytopathology 1993, 83, 995–1002. [Google Scholar] [CrossRef]
- Smith, D.D.; Williams, A.N.; Verrett, J.N.; Bergbusch, N.T.; Manning, V.; Trippe, K.; Stavrinides, J. Resistance to two vinylglycine antibiotic analogs is conferred by inactivation of two separate amino acid transporters in erwinia amylovora. J. Bacteriol. 2019, 201, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayan, F.E. Herbicide Discovery. Plants 2019, 8, 341. [Google Scholar] [CrossRef] [Green Version]
- Davis, E.; Okrent, R.A.; Manning, V.A.; Trippe, K.M. Distribution and diversity of the 4-formylaminooxyvinylglycine biosynthetic pathway in Pseudomonas. bioRxiv 2021. [Google Scholar] [CrossRef]
- Chou, H.T.; Li, J.Y.; Peng, Y.C.; Lu, C.D. Molecular Characterization of PauR and Its Role in Control of Putrescine and Cadaverine Catabolism through the gamma-Glutamylation Pathway in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2013, 195, 3906–3913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suvorova, I.A.; Rodionov, D.A. Comparative genomics of pyridoxal 5′-phosphate-dependent transcription factor regulons in Bacteria. Microb. Genom. 2016, 2, e000047. [Google Scholar] [CrossRef] [PubMed]
- Hassan, K.A.; Johnson, A.; Shaffer, B.T.; Ren, Q.; Kidarsa, T.A.; Elbourne, L.D.; Hartney, S.; Duboy, R.; Goebel, N.C.; Zabriskie, T.M.; et al. Inactivation of the GacA response regulator in Pseudomonas fluorescens Pf-5 has far-reaching transcriptomic consequences. Environ. Microbiol. 2010, 12, 899–915. [Google Scholar] [CrossRef]
- Wang, D.; Lee, S.H.; Seeve, C.; Yu, J.M.; Pierson, L.S.; Pierson, E.A. Roles of the Gac-Rsm pathway in the regulation of phenazine biosynthesis in Pseudomonas chlororaphis 30–84. Microbiologyopen 2013, 2, 505–524. [Google Scholar] [CrossRef] [PubMed]
- Paynel, F.; Murray, P.J.; Bernard Cliquet, J. Root exudates: A pathway for short-term N transfer from clover and ryegrass. Plant Soil 2001, 229, 235–243. [Google Scholar] [CrossRef]
- Bobille, H.; Limami, A.M.; Robins, R.J.; Cukier, C.; Le Floch, G.; Fustec, J. Evolution of the amino acid fingerprint in the unsterilized rhizosphere of a legume in relation to plant maturity. Soil Biol. Biochem. 2016, 101, 226–236. [Google Scholar] [CrossRef]
- Phillips, D.A.; Fox, T.C.; King, M.D.; Bhuvaneswari, T.V.; Teuber, L.R. Microbial products trigger amino acid exudation from plant roots. Plant Physiol. 2004, 136, 2887–2894. [Google Scholar] [CrossRef] [Green Version]
- Amaya-Gómez, C.V.; Porcel, M.; Mesa-Garrig, L.; Gómez-Álvarez, M.I. A framework for the selection of plant growth-promoting rhizobacteria based on bacterial competence mechanisms. Appl. Environ. Microbiol. 2020, 86, 1–13. [Google Scholar] [CrossRef]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [Green Version]
- Barret, M.; Morrissey, J.P.; O’Gara, F. Functional genomics analysis of plant growth-promoting rhizobacterial traits involved in rhizosphere competence. Biol. Fertil. Soils 2011, 47, 729–743. [Google Scholar] [CrossRef]
- Ghirardi, S.; Dessaint, F.; Mazurier, S.; Corberand, T.; Raaijmakers, J.M.; Meyer, J.M.; Dessaux, Y.; Lemanceau, P. Identification of Traits Shared by Rhizosphere-Competent Strains of Fluorescent Pseudomonads. Microb. Ecol. 2012, 64, 725–737. [Google Scholar] [CrossRef] [PubMed]
- Zboralski, A.; Filion, M. Genetic factors involved in rhizosphere colonization by phytobeneficial Pseudomonas spp. Comput. Struct. Biotechnol. J. 2020, 18, 3539–3554. [Google Scholar] [CrossRef] [PubMed]
- Capdevila, S.; Martínez-Granero, F.M.; Sánchez-Contreras, M.; Rivilla, R.; Martín, M. Analysis of Pseudomonas fluorescens F113 genes implicated in flagellar filament synthesis and their role in competitive root colonization. Microbiology 2004, 150, 3889–3897. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Granero, F.; Rivilla, R.; Martín, M. Rhizosphere selection of highly motile phenotypic variants of Pseudomonas fluorescens with enhanced competitive colonization ability. Appl. Environ. Microbiol. 2006, 72, 3429–3434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Weert, S.; Vermeiren, H.; Mulders, I.H.; Kuiper, I.; Hendrickx, N.; Bloemberg, G.V.; Vanderleyden, J.; De Mot, R.; Lugtenberg, B.J. Flagella-driven chemotaxis towards exudate components is an important trait for tomato root colonization by Pseudomonas fluorescens. Mol. Plant-Microbe Interact. 2002, 15, 1173–1180. [Google Scholar] [CrossRef] [Green Version]
- Oku, S.; Komatsu, A.; Tajima, T.; Nakashimada, Y.; Kato, J. Identification of Chemotaxis Sensory Proteins for Amino Acids in Pseudomonas fluorescens Pf0-1 and Their Involvement in Chemotaxis to Tomato Root Exudate and Root Colonization. Microbes Environ. 2012, 27, 462–469. [Google Scholar] [CrossRef] [Green Version]
- Ramey, B.E.; Koutsoudis, M.; Bodman, S.B.; Fuqua, C. Biofilm formation in plant-microbe associations. Curr. Opin. Microbiol. 2004, 7, 602–609. [Google Scholar] [CrossRef] [Green Version]
- Ueda, A.; Saneoka, H. Characterization of the Ability to Form Biofilms by Plant-Associated Pseudomonas Species. Curr. Microbiol. 2015, 70, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Noirot-Gros, M.F.; Shinde, S.; Larsen, P.E.; Zerbs, S.; Korajczyk, P.J.; Kemner, K.M.; Noirot, P.H. Dynamics of aspen roots colonization by Pseudomonads reveals strain-specific and mycorrhizal-specific patterns of biofilm formation. Front. Microbiol. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Chitnis, C.E.; Ohman, D.E. Genetic analysis of the alginate biosynthetic gene cluster of Pseudomonas aeruginosa shows evidence of an operonic structure. Mol. Microbiol. 1993, 8, 583–590. [Google Scholar] [CrossRef]
- Ghiglione, J.F.; Gourbiere, F.; Potier, P.; Philippot, L.; Lensi, R. Role of respiratory nitrate reductase in ability of Pseudomonas fluorescens YT101 to colonize the rhizosphere of maize. Appl. Environ. Microbiol. 2000, 66, 4012–4016. [Google Scholar] [CrossRef] [Green Version]
- Philippot, L.; Clays-josserand, A.; Lensi, R.; Microbienne, E.; Cnrs, U.R.; Ce, V. Use of Tn 5 Mutants To Assess the Role of the Dissimilatory Nitrite Reductase in the Competitive Abilities of Two Pseudomonas Strains in Soil. Appl. Environ. Microbiol. 1995, 61, 1426–1430. [Google Scholar] [CrossRef] [Green Version]
- Ghequire, M.G.; De Mot, R. Ribosomally encoded antibacterial proteins and peptides from Pseudomonas. FEMS Microbiol. Rev. 2014, 38, 523–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, J.B.; West, S.A.; Griffin, A.S. Bacteriocins and the assembly of natural Pseudomonas fluorescens populations. J. Evol. Biol. 2017, 30, 352–360. [Google Scholar] [CrossRef]
- Dorosky, R.J.; Iii, S.P.; Pierson, A. Crossm Pseudomonas chlororaphis produces multiple R-tailocin Particles That Broaden the Killing Spectrum and Contribute to Persistence in Rhizosphere Communities. Appl. Environ. Microbiol. 2018, 84, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couée, I.; Hummel, I.; Sulmon, C.; Gouesbet, G.; El Amrani, A. Involvement of polyamines in root development. Plant Cell. Tissue Organ Cult. 2004, 76, 1–10. [Google Scholar] [CrossRef]
- Forde, B.G. Glutamate signalling in roots. J. Exp. Bot. 2014, 65, 779–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saini, S.; Sharma, I.; Kaur, N.; Pati, P.K. Auxin: A master regulator in plant root development. Plant Cell Rep. 2013, 32, 741–757. [Google Scholar] [CrossRef]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef] [Green Version]
- Růžička, K.; Ljung, K.; Vanneste, S.; Podhorská, R.; Beeckman, T.; Friml, J.; Benková, E. Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution. Plant Cell 2007, 19, 2197–2212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glick, B.R.; Penrose, D.M.; Li, J. A model for the lowering of plant ethylene concentrations by plant growth-promoting bacteria. J. Theor. Biol. 1998, 190, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.A.; Peirson, D.; Ghosh, S.; Glick, B.R. Root elongation in various agronomic crops by the plant growth promoting rhizobacterium pseudomonas putida Gr12–2. Isr. J. Plant Sci. 1996, 44, 37–42. [Google Scholar] [CrossRef]
- Ribaudo, C.M.; Krumpholz, E.M.; Cassán, F.D.; Bottini, R.; Cantore, M.L.; Curá, J.A. Azospirillum sp. Promotes root hair development in tomato plants through a mechanism that involves ethylene. J. Plant Growth Regul. 2006, 25, 175–185. [Google Scholar] [CrossRef]
- Yu, Y.B.; Yang, S.F. Auxin-induced Ethylene Production and Its Inhibition by Aminoethyoxyvinylglycine and Cobalt Ion. Plant Physiol. 1979, 64, 1074–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Deunff, E.; Beauclair, P.; Deleu, C.; Lecourt, J. Inhibition of Aminotransferases by Aminoethoxyvinylglycine Triggers a Nitrogen Limitation Condition and Deregulation of Histidine Homeostasis That Impact Root and Shoot Development and Nitrate Uptake. Front. Plant Sci. 2019, 10, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
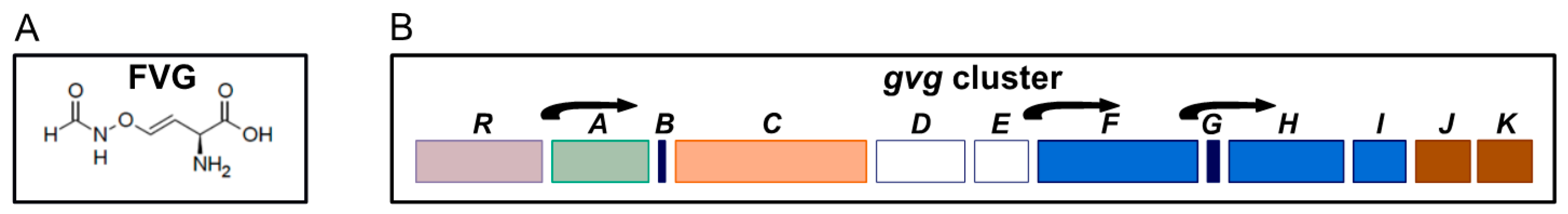
| NZ_CM001025.1 (CM001025.1) | Gene | Protein Features/ Conserved Domains | Putative Function | Antibacterial/Herbicidal Phenotype of Knockout a |
|---|---|---|---|---|
| PFWH6_RS25400 (PFWH6_5248) | gvgR | Aminotransferase class I and II; helix-turn-helix DNA-binding domain | GntR transcriptional regulator | − |
| PFWH6_RS25405 (PFWH6_5249) | gvgA | Isoamyl acetate hydrolase-like | Lipase/esterase | − |
| (PFWH6_5250) | gvgB | Lysine rich | − | |
| PFWH6_RS25410 (PFWH6_5251) | gvgC | Heme-oxygenase 2 | Redox enzyme | − |
| PFWH6_RS25415 (PHWH6_5252) | gvgD | Amidinotransferase | Amidinotransferase | + |
| PFWH6_RS25420 (PHWH6_5253) | gvgE | LysE type translocator | Amino acid exporter | + |
| PFWH6_RS25425 (PHWH6_5254) | gvgF | NodU family carbamoyltransferase | Carbamoyltransferase | − |
| (PHWH6_5255) | gvgG | Signal peptide | − | |
| PFWH6_RS25430 (PHWH6_5256) | gvgH | Aminotransferase class III | Aminotransferase | − |
| PFWH6_RS25435 (PHWH6_5257) | gvgI | Formyltransferase | Formyltransferase | + |
| PFWH6_RS25440 (PFWH6_5258) | gvgJ | LysE type translocator | Amino acid exporter | + |
| PFWH6_RS25445 (PHWH6_5259) | gvgK | LysE type translocator | Amino acid exporter | + |
| Strain | ||||||
|---|---|---|---|---|---|---|
| Locus Tag | ΔgvgA | ΔgvgC | ΔgvgR | Gene Name | Annotation | Pathway |
| Upregulated | ||||||
| PFWH6_RS05650 | 5.51 | 3.46 | 3.64 | pyocin terminase, ssu (pyocin cluster) | ||
| PFWH6_RS05685 | 6.12 | 3.42 | 3.82 | hypothetical protein | pyocin cluster | |
| PFWH6_RS05705 | 5.96 | 3.81 | 6.26 | hypothetical protein | pyocin cluster | |
| PFWH6_RS10425 | 6.49 | 8.37 | 4.13 | aldehyde dehydrogenase | ||
| PFWH6_RS16565 | 5.67 | 6.05 | 5.17 | narL | transcriptional regulator | nitrate assimilation |
| PFWH6_RS16575 | 9.31 | 13.15 | 8.42 | yhbT | lipid carrier protein | |
| PFWH6_RS16580 | 5.44 | 6.72 | 6.8 | yhbV | protease | |
| PFWH6_RS16585 | 7.26 | 7.23 | 6.43 | yhbU | protease | |
| PFWH6_RS16590 | 5.26 | 4.86 | 5.65 | moeA1 | molybdenum cofactor biosynthesis protein A | molybdenum cofactor biosynthesis |
| PFWH6_RS16595 | 5.69 | 6.67 | 6.11 | moaB1 | molybdopterin biosynthesis protein B | molybdenum cofactor biosynthesis |
| Downregulated | ||||||
| gvgB | −109.44 | −40.51 | −123.51 | gvgB | FVG biosynthesis | |
| PFWH6_RS25405 | −109.25 | −90.91 | −77.07 | gvgA | lipase/esterase | FVG biosynthesis |
| PFWH6_RS25410 | −102.18 | −185.79 | −91.89 | gvgC | redox enzyme | FVG biosynthesis |
| PFWH6_RS25425 | −92.04 | −107.74 | −74.38 | gvgF | Carbamoyltransferase | FVG biosynthesis |
| gvgG | −89.21 | −126.14 | −60.98 | gvgG | secreted protein | FVG biosynthesis |
| PFWH6_RS25415 | −87.03 | −113.93 | −76.31 | gvgD | amidinotransferase | FVG biosynthesis |
| PFWH6_RS25430 | −75.3 | −80.03 | −53.95 | gvgH | aminotransferase | FVG biosynthesis |
| PFWH6_RS25420 | −59.1 | −65.66 | −58.76 | gvgE | LysE transporter | FVG biosynthesis |
| PFWH6_RS25445 | −50.13 | −71.72 | −49.13 | gvgK | LysE transporter | FVG biosynthesis |
| PFWH6_RS25435 | −40.1 | −51.65 | −38.8 | gvgI | formyltransferase | FVG biosynthesis |
| Digital Expression (cpm a) Average % WT | Droplet Digital Expression Average % WT | |||||||
|---|---|---|---|---|---|---|---|---|
| Gene Detected | ΔgvgA | ΔgvgC | ΔgvgR | ΔgvgA | ΔgvgC | ΔgvgR | ΔgvgH | ΔgvgI |
| gvgR | 9.7 | 9.5 | 1.1 | 7.5 | 9.5 | 0.4 | 9.2 | 178.6 |
| gvgA | 0.9 | 1.1 | 1.3 | 0.0 | 0.5 | 0.7 | 0.3 | 138.3 |
| gvgC | 1.0 | 0.5 | 1.1 | 0.2 | 0.0 | 0.6 | 0.4 | 130.4 |
| gvgH | 1.3 | 1.2 | 1.9 | 0.6 | 0.6 | 1.0 | 0.0 | 111.3 |
| gvgI | 2.5 | 1.9 | 2.6 | 0.4 | 0.6 | 1.0 | 0.8 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manning, V.A.; Trippe, K.M. Absence of 4-Formylaminooxyvinylglycine Production by Pseudomonas fluorescens WH6 Results in Resource Reallocation from Secondary Metabolite Production to Rhizocompetence. Microorganisms 2021, 9, 717. https://doi.org/10.3390/microorganisms9040717
Manning VA, Trippe KM. Absence of 4-Formylaminooxyvinylglycine Production by Pseudomonas fluorescens WH6 Results in Resource Reallocation from Secondary Metabolite Production to Rhizocompetence. Microorganisms. 2021; 9(4):717. https://doi.org/10.3390/microorganisms9040717
Chicago/Turabian StyleManning, Viola A., and Kristin M. Trippe. 2021. "Absence of 4-Formylaminooxyvinylglycine Production by Pseudomonas fluorescens WH6 Results in Resource Reallocation from Secondary Metabolite Production to Rhizocompetence" Microorganisms 9, no. 4: 717. https://doi.org/10.3390/microorganisms9040717
APA StyleManning, V. A., & Trippe, K. M. (2021). Absence of 4-Formylaminooxyvinylglycine Production by Pseudomonas fluorescens WH6 Results in Resource Reallocation from Secondary Metabolite Production to Rhizocompetence. Microorganisms, 9(4), 717. https://doi.org/10.3390/microorganisms9040717