
Temperature Sensitivity and Composition of Nitrate-Reducing Microbiomes from a Full-Scale Woodchip Bioreactor Treating Agricultural Drainage Water

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Woodchip Bioreactor Facility and Sampling
2.2. Enrichment Cultures
2.3. Temperature Dependence of NO3− Depletion, Denitrification and Growth
2.4. Metagenomic Analyses
2.5. Calculations and Statistical Analyses
3. Results
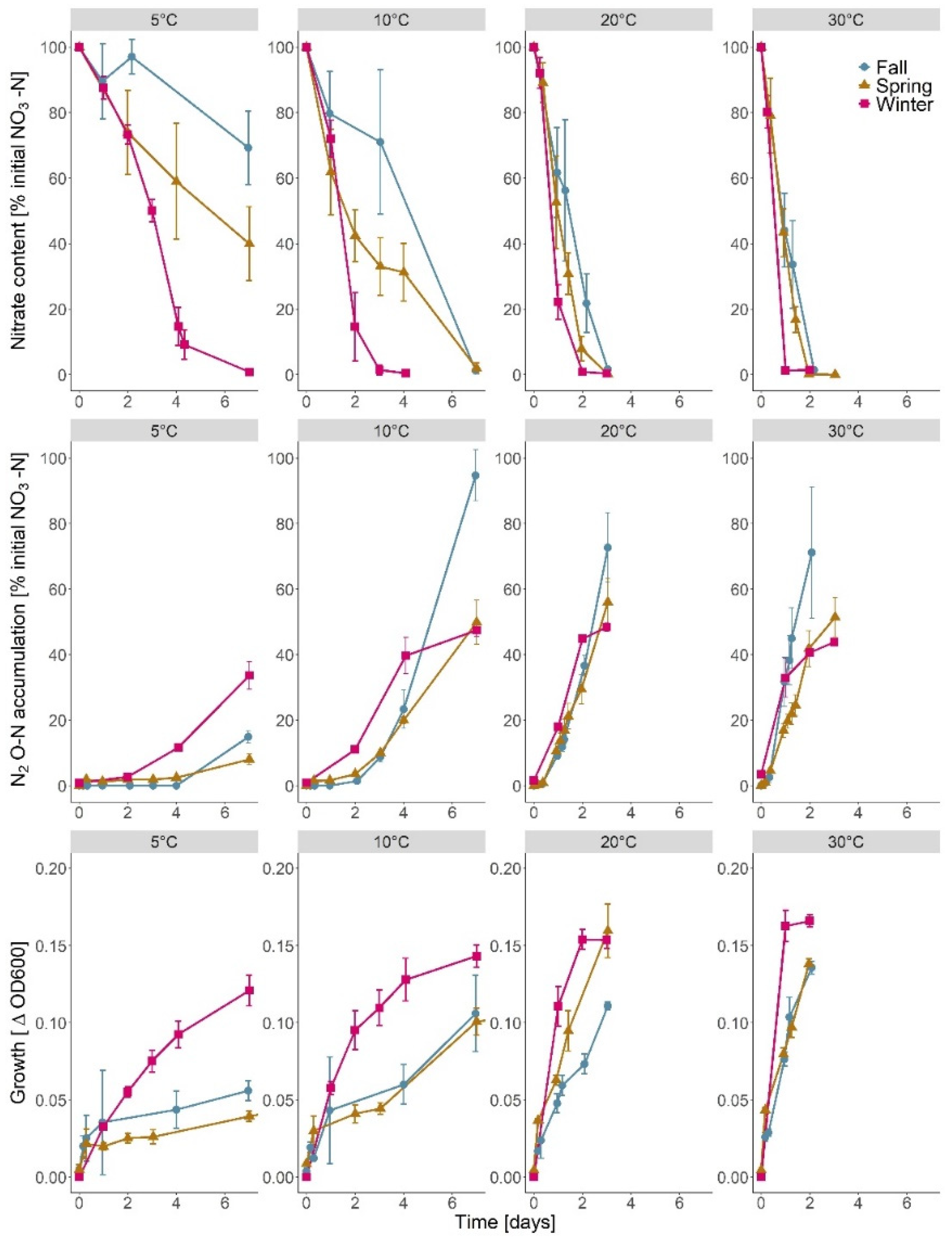
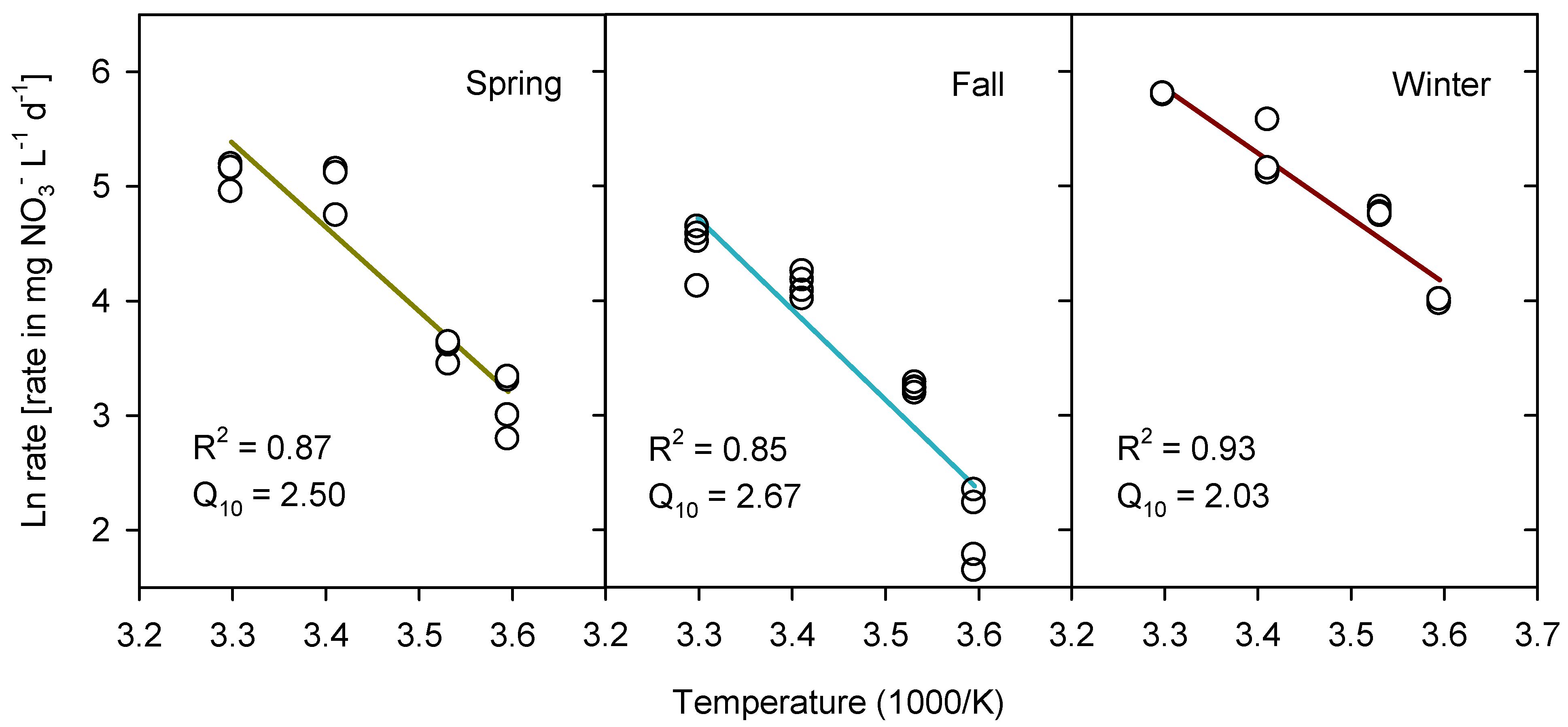
3.1. Nitrate Depletion, Denitrification and Growth
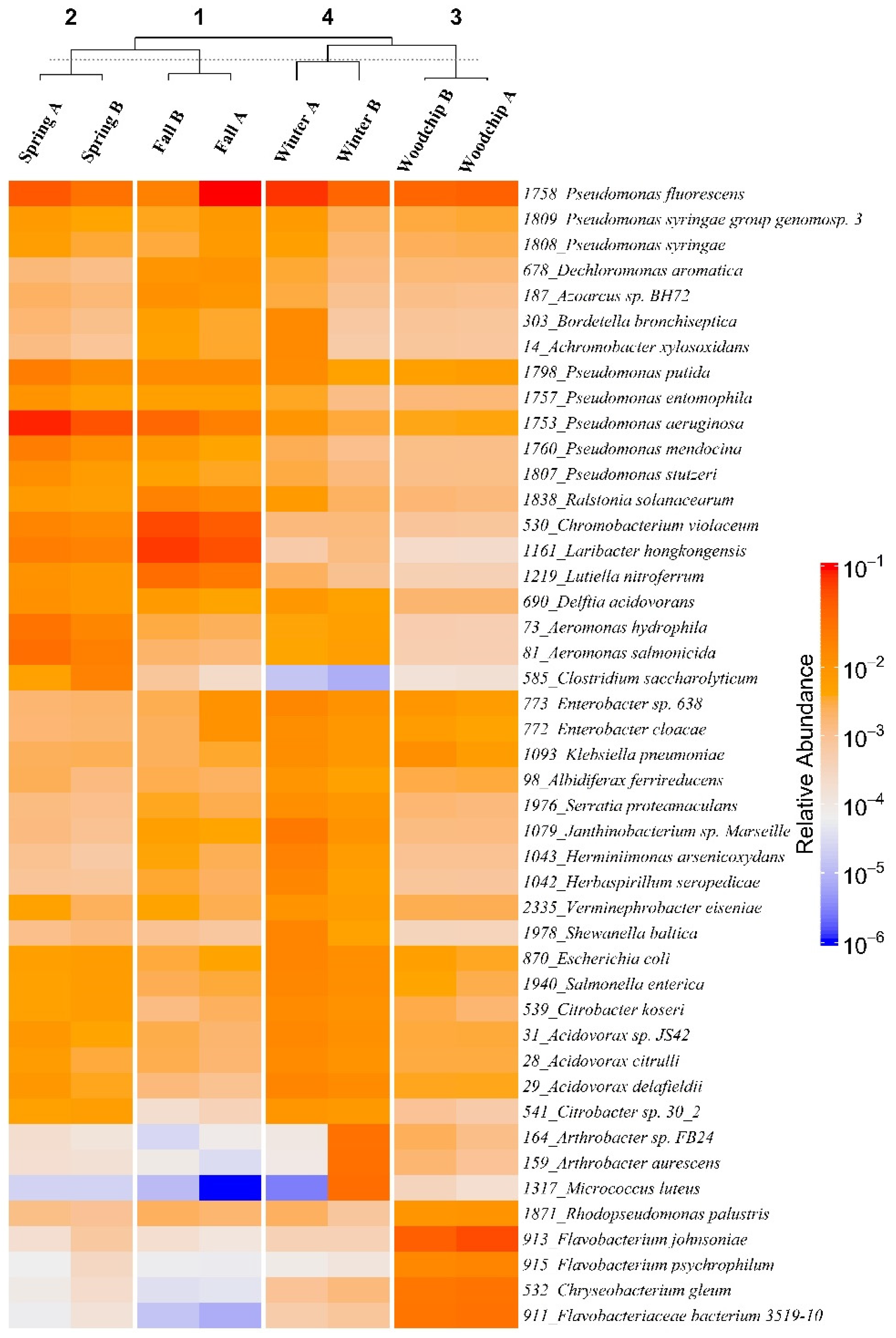
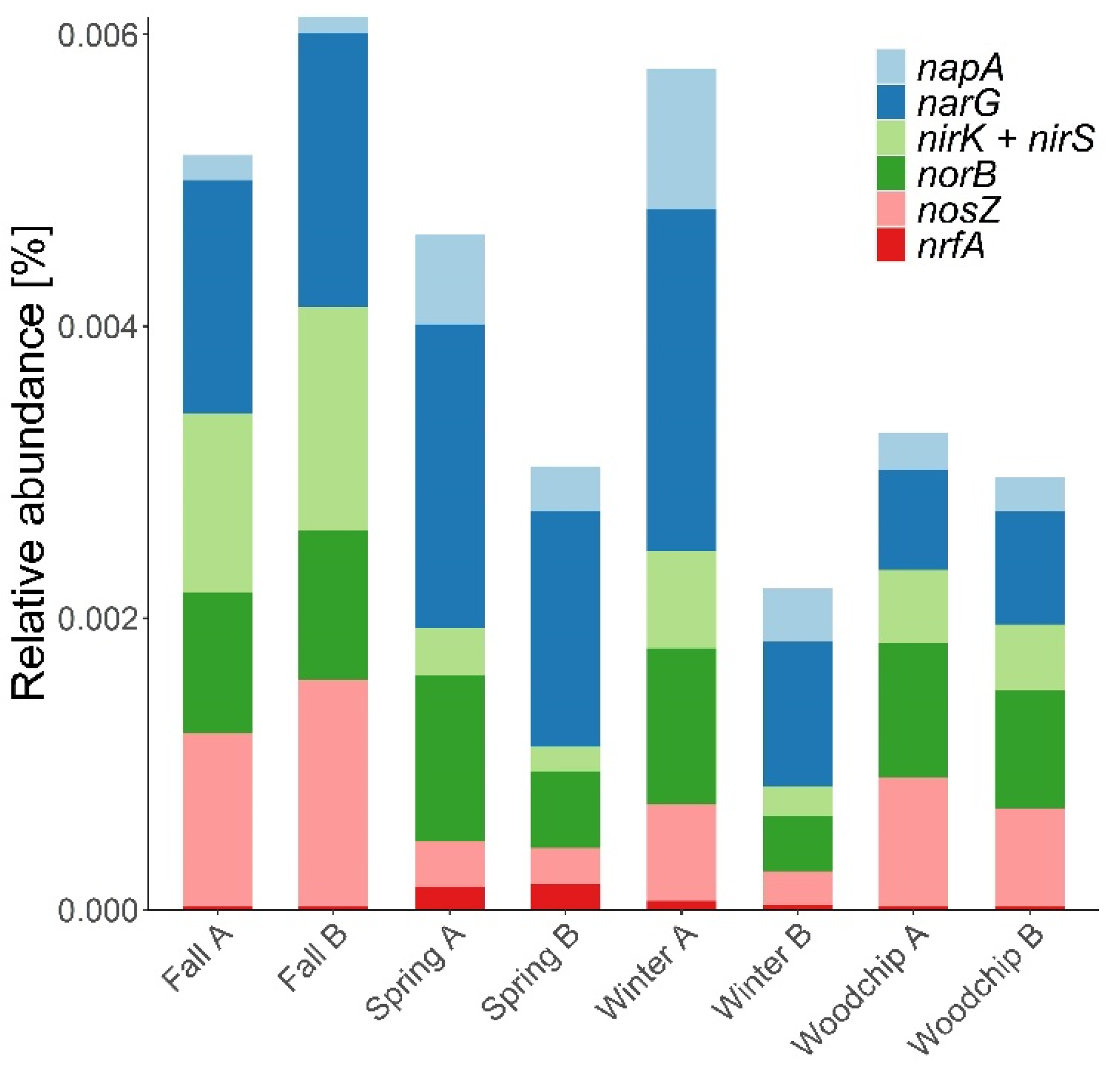
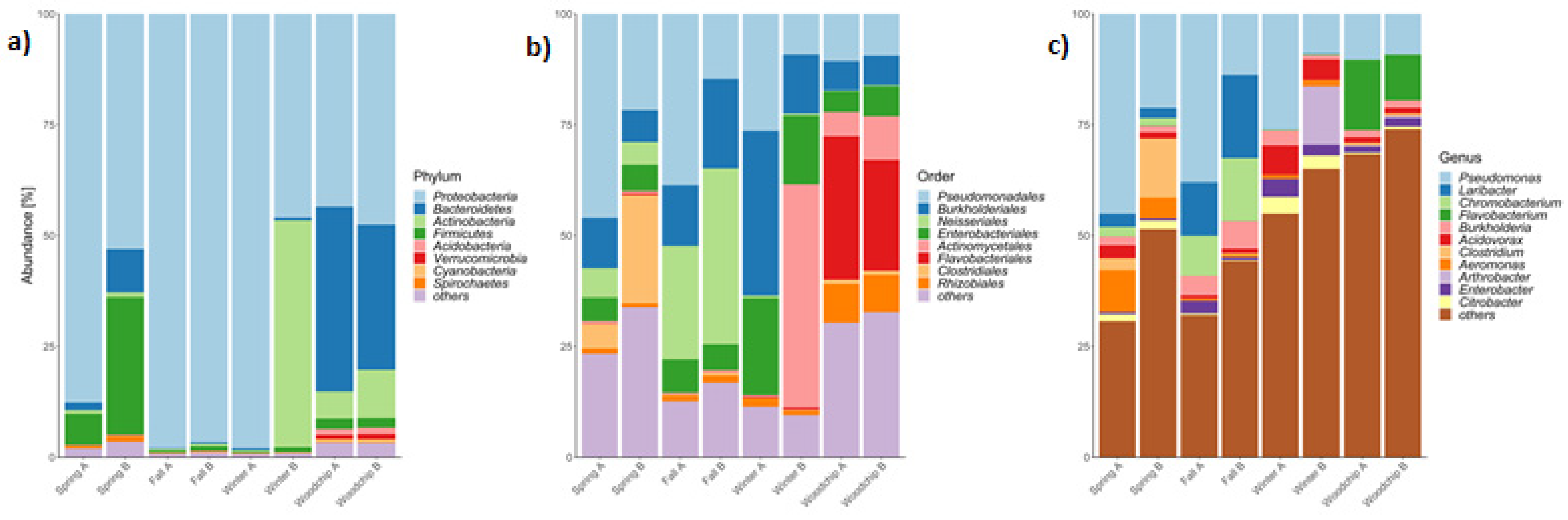
3.2. Microbiome Composition and Functional Genes
4. Discussion
4.1. Temperature Dependence of Nitrate Removal
4.2. Taxonomic and Functional Composition
4.3. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | MG-RAST ID | ENA Project ID | ENA Sample ID | Base Pairs Count |
|---|---|---|---|---|
| Spring A | mgm4922152.3 | PRJRB45276 | ERS6495196 | 1,531,600,118 |
| Spring B | mgm4922153.3 | PRJRB45276 | ERS6495197 | 1,514,469,895 |
| Fall A | mgm4922158.3 | PRJRB45276 | ERS6495194 | 1,173,823,046 |
| Fall B | mgm4922157.3 | PRJRB45276 | ERS6495195 | 1,655,202,384 |
| Winter A | mgm4922159.3 | PRJRB45276 | ERS6495198 | 1,347,767,217 |
| Winter B | mgm4922154.3 | PRJRB45276 | ERS6495199 | 849,914,651 |
| Woodchip A | mgm4922155.3 | PRJRB45276 | ERS6495200 | 1,930,225,161 |
| Woodchip B | mgm4922160.3 | PRJRB45276 | ERS6495201 | 1,417,188,496 |
| Gene | KO | Name |
|---|---|---|
| nar | K00370 | narG: nitrate reductase 1, alpha subunit [EC:1.7.99.4]; |
| K02567 | napA: periplasmic nitrate reductase [EC:1.7.99.4] | |
| nir | K00368 | nir (nirS and nirK): nitrite reductase (NO-forming) [EC:1.7.2.1] |
| nor | K04561 | norB: nitric oxide reductase subunit B [EC:1.7.2.5] |
| nosZ | K00376 | nosZ: nitrous-oxide reductase [EC:1.7.2.4] |
| nrfA | K03385 | nrfA: nitrite reductase by formate, cytochrome c-552 [EC:1.7.2.2] |
| Enrichment | NO3− Depletion Rate (mg NO3− L−1 d−1) | |||
|---|---|---|---|---|
| 5 °C | 10 °C | 20 °C | 30 °C | |
| Spring | 22.9 b (±2.8) | 35.8 b (±1.5) | 155.8 a (±13.8) | 166.5 b (±8.6) |
| Fall | 7.7 c (±1.3) | 25.4 c (±0.5) | 62.5 b (±3.3) | 88.4 c (±9.3) |
| Winter | 54.2 a (±0.5) | 117.6 a (±2.0) | 193.6 a (±23.4) | 329.7 a (±1.0) |

References
- Lu, C.; Tian, H. Global nitrogen and phosphorus fertilizer use for agriculture production in the past half century: Shifted hot spots and nutrient imbalance. Earth Syst. Sci. Data 2017, 9, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Di, H.J.; Cameron, K.C. Nitrate leaching in temperate agroecosystems: Sources, factors and mitigating strategies. Nutr. Cycl. Agroecosyst. 2002, 64, 237–256. [Google Scholar] [CrossRef]
- Howarth, R.; Chan, F.; Conley, D.J.; Garnier, J.; Doney, S.C.; Marino, R.; Billen, G. Coupled biogeochemical cycles: Eutrophication and hypoxia in temperate estuaries and coastal marine ecosystems. Front. Ecol. Environ. 2011, 9, 18–26. [Google Scholar] [CrossRef] [Green Version]
- Møller, A.B.; Beucher, A.; Iversen, B.V.; Greve, M.H. Predicting artificially drained areas by means of a selective model ensemble. Geoderma 2018, 320, 30–42. [Google Scholar] [CrossRef]
- Kronvang, B.; Andersen, H.E.; Børgesen, C.; Dalgaard, T.; Larsen, S.E.; Bøgestrand, J.; Blicher-Mathiasen, G. Effects of policy measures implemented in Denmark on nitrogen pollution of the aquatic environment. Environ. Sci. Policy 2008, 11, 144–152. [Google Scholar] [CrossRef]
- Hoffmann, C.C.; Zak, D.; Kronvang, B.; Kjaergaard, C.; Carstensen, M.V.; Audet, J. An overview of nutrient transport mitigation measures for improvement of water quality in Denmark. Ecol. Eng. 2020, 155, 105863. [Google Scholar] [CrossRef]
- Christianson, L.; Helmers, M.; Bhandari, A.A. Practice-oriented review of woodchip bioreactors for subsurface agricultural drainage. Appl. Eng. Agric. 2012, 28, 861–874. [Google Scholar] [CrossRef]
- Schipper, L.A.; Robertson, W.D.; Gold, A.J.; Jaynes, D.B.; Cameron, S.C. Denitrifying bioreactors: An approach for reducing nitrate loads to receiving waters. Ecol. Eng. 2010, 36, 1532–1543. [Google Scholar] [CrossRef]
- Grießmeier, V.; Wienhöfer, J.; Horn, H.; Gescher, J. Assessing and modeling biocatalysis in field denitrification beds reveals key influencing factors for future constructions. Water Res. 2021, 188, 116467. [Google Scholar] [CrossRef]
- Christianson, L.; Bhandari, A.; Helmers, M.; Kult, K.; Sutphin, T.; Wolf, R. Performance evaluation of four field-scale agricultural drain-age denitrification bioreactors in Iowa. Trans. ASABE 2012, 55, 2163–2174. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, C.C.; Larsen, S.E.; Kjaergaard, C. Nitrogen removal in woodchip-based biofilters of variable designs treating agricultural drainage discharges. J. Environ. Qual. 2019, 48, 1881–1889. [Google Scholar] [CrossRef] [Green Version]
- Feyereisen, G.; Rosen, C.; Ishii, S.; Wang, P.; Ghane, E.; Sadowsky, M. Optimizing Woodchip Bioreactors to Treat Nitrogen and Phosphorus in Subsurface Drainage Water. Final Report for Minnesota Department of Agriculture, USA, Project #108837. 2018, pp. 1–46. Available online: https://wrl.mnpals.net/islandora/object/WRLrepository:3447 (accessed on 10 October 2020).
- Maxwell, B.M.; Díaz-García, C.; Martínez-Sánchez, J.J.; Birgand, F.; Álvarez-Rogel, J. Temperature sensitivity of nitrate removal in woodchip bioreactors increases with woodchip age and following drying–rewetting cycles. Environ. Sci. Water Res. Technol. 2020, 6, 2752. [Google Scholar] [CrossRef]
- Roser, M.B.; Feyereisen, G.W.; Spokas, K.A.; Mulla, D.J.; Strock, J.S.; Gutknecht, J. Carbon dosing increases nitrate removal rates in denitrifying bioreactors at low-temperature high-flow conditions. J. Environ. Qual. 2018, 47, 856–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zumft, W.G. Cell biology and molecular basis of denitrification. Microbiol. Mol. Biol. Rev. 1997, 61, 533–616. [Google Scholar] [CrossRef]
- Shapleigh, J.P. Denitrifying prokaryotes. In The Prokaryotes: Prokaryotic Physiology and Biochemistry; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 405–425. [Google Scholar]
- Lazcano, C.; Zhu-Barker, X.; Decock, C. Effects of organic fertilizers on the soil microorganisms responsible for N2O emissions: A review. Microorganisms 2021, 9, 983. [Google Scholar] [CrossRef]
- Schipper, L.A.; Vojvodić-Vuković, M. Five years of nitrate removal, denitrification and carbon dynamics in a denitrification wall. Water Res. 2001, 35, 3473–3477. [Google Scholar] [CrossRef]
- Jang, J.; Anderson, E.L.; Venterea, R.T.; Sadowsky, M.J.; Rosen, C.J.; Feyereisen, G.W.; Ishii, S. Denitrifying bacteria active in woodchip bioreactors at low-temperature conditions. Front. Microbiol. 2019, 10, 635. [Google Scholar] [CrossRef]
- Nordström, A.; Hellman, M.; Hallin, S.; Herbert, R.B. Microbial controls on net production of nitrous oxide in a denitrifying woodchip bioreactor. J. Environ. Qual. 2021, 50, 228–240. [Google Scholar] [CrossRef]
- Canion, A.; Prakash, O.; Green, S.J.; Jahnke, L.; Kuypers, M.M.; Kostka, J.E. Isolation and physiological characterization of psychrophilic denitrifying bacteria from permanently cold Arctic fjord sediments (Svalbard, Norway). Environ. Microbiol. 2013, 15, 1606–1618. [Google Scholar] [CrossRef]
- Anderson, E.L.; Jang, J.; Venterea, R.T.; Feyereisen, G.W.; Ishii, S. Isolation and characterization of denitrifiers from woodchip bioreactors for bioaugmentation application. J. Appl. Microbiol. 2020, 129, 590–600. [Google Scholar] [CrossRef]
- King, D.; Nedwell, D. Changes in the nitrate-reducing community of an anaerobic saltmarsh sediment in response to seasonal selection by temperature. Microbiology 1984, 130, 2935–2941. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.; Dong, L.; Wilson, J.; Stott, A.; Osborn, A.; Nedwell, D. Seasonal variation in denitrification and dissimilatory nitrate reduction to ammonia process rates and corresponding key functional genes along an estuarine nitrate gradient. Front. Microbiol. 2015, 6, 542. [Google Scholar] [CrossRef]
- El Fantroussi, S.; Agathos, S.N. Is bioaugmentation a feasible strategy for pollutant removal and site remediation? Curr. Opin. Microbiol. 2005, 8, 268–275. [Google Scholar] [CrossRef]
- Andersen, H.E.; Kronvang, B.; Larsen, S.E.; Hoffmann, C.C.; Jensen, T.S.; Rasmussen, E.K. Climate-change impacts on hydrology and nutrients in a Danish lowland river basin. Sci. Total Environ. 2006, 365, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Butler, I.B.; Schoonen, M.A.A.; Rickard, D.T. Removal of dissolved oxygen from water: A comparison of four common techniques. Talanta 1994, 41, 211–215. [Google Scholar] [CrossRef]
- Wolfe, R.S. Microbial formation of methane. Adv. Microb. Physiol. 1971, 6, 107–146. [Google Scholar]
- ISO7890-1:1986. Water Quality—Determination of Nitrate—Part 1: 2,6-Dimethylphenol Spectrometric Method; ISO: Geneva, Switzerland, 1986; p. 5. [Google Scholar]
- Yoshinari, T.; Knowles, R. Acetylene inhibition of nitrous oxide reduction by denitrifying bacteria. Biochem. Biophys. Res. Commun. 1976, 69, 705–710. [Google Scholar] [CrossRef]
- Widdel, F. Theory and measurement of bacterial growth. In Grundpraktikum Mikrobiologie; Universität Bremen: Bremen, Germany, 2010; p. 11. [Google Scholar]
- Petersen, S.O.; Hoffmann, C.; Schäfer, C.-M.; Blicher-Mathiesen, G.; Elsgaard, L.; Kristensen, K.; Larsen, S.E.; Torp, S.B.; Greve, M.H. Annual emissions of CH4 and N2O, and ecosystem respiration, from eight organic soils in Western Denmark managed by agriculture. Biogeosciences 2012, 9, 403–422. [Google Scholar] [CrossRef] [Green Version]
- Meyer, F.; Paarmann, D.; D’Souza, M.; Olson, R.; Glass, E.M.; Kubal, M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; et al. The metagenomics RAST server—A public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC Bioinform. 2018, 9, 386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S.; Kawashima, S.; Okuno, Y.; Hattori, M. The KEGG resource for deciphering the genome. Nucleic Acids Res. 2004, 32, D277–D280. [Google Scholar] [CrossRef] [Green Version]
- Randle-Boggis, R.J.; Helgason, T.; Sapp, M.; Ashton, P.D. Evaluating techniques for metagenome annotation using simulated sequence data. FEMS Microbiol. Ecol. 2016, 92, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilling, M.J. Reaction Kinetics; Clarendon Press: Oxford, UK, 1975. [Google Scholar]
- Myrold, D.D. Quantification of nitrogen transformations. In Manual of Environmental Microbiology; Hurst, C.J., Knudsen, G.R., McInernay, M.J., Stetzenbach, L.D., Walter, M.V., Eds.; American Society for Microbiology Press: Washington, DC, USA, 1997; pp. 445–452. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing. In R Foundation for Statistical Computing; Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 12 November 2020).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R package Version 3.1-152. 2021. Available online: https://cran.R-project.org/package=nlme (accessed on 12 November 2020).
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmke, E.; Weyland, H. Psychrophilic versus psychrotolerant bacteria—Occurrence and significance in polar and temperate marine habitats. Cell. Mol. Biol. 2004, 50, 553–561. [Google Scholar] [PubMed]
- Moyer, C.L.; Eric Collins, R.; Morita, R.Y. Psychrophiles and Psychrotrophs. In Reference Module in Life Sciences; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar] [CrossRef]
- Yu, K.; Struwe, S.; Kjøller, A.; Chen, G. Denitrification rate determined by nitrate disappearance is higher than determined by nitrous oxide production with acetylene blockage. Ecol. Eng. 2008, 32, 90–96. [Google Scholar] [CrossRef]
- Holtan-Hartwig, L.; Dörsch, P.; Bakken, L.R. Low temperature control of soil denitrifying communities: Kinetics of N2O production and reduction. Soil Biol. Biochem. 2002, 34, 1797–1806. [Google Scholar] [CrossRef]
- Addy, K.; Gold, A.J.; Christianson, L.E.; David, M.B.; Schipper, L.A.; Ratigan, N.A. Denitrifying bioreactors for nitrate removal: A meta-analysis. J. Environ. Qual. 2016, 45, 873–881. [Google Scholar] [CrossRef]
- Hellman, M.; Hubalek, V.; Juhanson, J.; Almstrand, R.; Peura, S.; Hallin, S. Substrate type determines microbial activity and community composition in bioreactors for nitrate removal by denitrification at low temperature. Sci. Total Environ. 2021, 755, 143023. [Google Scholar] [CrossRef]
- Kouanda, A.; Hua, G. Determination of nitrate removal kinetics model parameters in woodchip bioreactors. Water Res. 2021, 195, 116974. [Google Scholar] [CrossRef]
- Bellini, M.I.; Kumaresan, D.; Tarlera, S.; Murrell, J.C.; Fernandez-Scavino, A. Identification of active denitrifiers by DNA-stable isotope probing and amplicon sequencing reveals Betaproteobacteria as responsible for attenuation of nitrate contamination in a low impacted aquifer. FEMS Microbiol. Ecol. 2018, 94, fix181. [Google Scholar] [CrossRef]
- Grießmeier, V.; Gescher, J. Influence of the potential carbon sources for field denitrification beds on their microbial diversity and the fate of carbon and nitrate. Front. Microbiol. 2018, 9, 1313. [Google Scholar] [CrossRef]
- Woodard, L.M.; Bielkie, A.R.; Eisses, J.F.; Ketchum, P.A. Occurrence of nitrate reductase and molybdopterin in Xanthomonas maltophilia. Appl. Environ. Microbiol. 1990, 56, 3766–3771. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, L.; Han, W.; Wang, X.; Guo, Z.; Peng, F.; Yang, F.; Kong, M.; Gao, Y.; Chao, J.; et al. Nitrate removal, spatiotemporal communities of denitrifiers and the importance of their genetic potential for denitrification in novel denitrifying bioreactors. Bioresour. Technol. 2017, 241, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Moore, E.; Tindall, B.; Santos, V.; Pieper, D.; Ramos, J.-L.; Palleroni, N. The Prokaryotes. In Nonmedical: Pseudomonas; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2005. [Google Scholar]
- Gamble, T.N.; Betlach, M.R.; Tiedje, J.M. Numerically dominant denitrifying bacteria from world soils. Appl. Environ. Microbiol. 1977, 33, 926–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grießmeier, V.; Bremges, A.; McHardy, A.C.; Gescher, J. Investigation of different nitrogen reduction routes and their key microbial players in wood chip-driven denitrification beds. Sci. Rep. 2017, 7, 17028. [Google Scholar] [CrossRef] [PubMed]
- Shannon, K.E.M.; Saleh-Lakha, S.; Burton, D.L.; Zebarth, B.J.; Goyer, C.; Trevors, J.T. Effect of nitrate and glucose addition on denitrification and nitric oxide reductase (cnorB) gene abundance and mRNA levels in Pseudomonas mandelii inoculated into anoxic soil. Antonie Leeuwenhoek 2011, 100, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, K.; Krieger, R.; Benkert, B.; Eschbach, M.; Arai, H.; Schobert, M.; Jahn, D. The anaerobic regulatory network required for Pseudomonas aeruginosa nitrate respiration. J. Bacteriol. 2007, 189, 4310–4314. [Google Scholar] [CrossRef] [Green Version]
- Heylen, K.; Lebbe, L.; De Vos, P. Acidovorax caeni sp. nov.; a denitrifying species with genetically diverse isolates from activated sludge. Int. J. Syst. Evol. Microbiol. 2008, 58, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Petersen, R.J.; Liang, Z.; Prinds, C.; Jéglot, A.; Thamdrup, B.; Kjaergaard, C.; Elsgaard, L. Nitrate reduction pathways and interactions with iron in the drainage water infiltration zone of a riparian wetland soil. Biogeochemistry 2020, 150, 235–255. [Google Scholar] [CrossRef]
- Ciok, A.; Budzik, K.; Zdanowski, M.K.; Gawor, J.; Grzesiak, J.; Decewicz, P.; Gromadka, R.; Bartosik, D.; Dziewit, L. Plasmids of psychrotolerant Polaromonas spp. isolated from Arctic and Antarctic glaciers—Diversity and role in adaptation to polar environments. Front. Microbiol. 2018, 9, 1285. [Google Scholar] [CrossRef]
- Irgens, R.; Gosink, J.; Staley, J. Polaromonas vacuolata gen. nov.; sp. nov.; a psychrophilic, marine, gas vacuolate bacterium from Antarctica. Int. J. Syst. Bacteriol. 1996, 46, 822–826. [Google Scholar] [CrossRef] [Green Version]
- Bernardet, J.-F.; Bowman, J.P. The Genus Flavobacterium. In The Prokaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.-H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 7, pp. 481–531. [Google Scholar]
- Pishgar, R.; Dominic, J.A.; Sheng, Z.; Tay, J.H. Denitrification performance and microbial versatility in response to different selection pressures. Bioresour. Technol. 2019, 281, 72–83. [Google Scholar] [CrossRef]
- Abdelhamed, H.; Nho, S.W.; Karsi, A.; Lawrence, M.L. The role of denitrification genes in anaerobic growth and virulence of Flavobacterium columnare. J. Appl. Microbiol. 2021, 130, 1062–1074. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Li, L.; Dai, Z.; Senbati, Y.; Song, K.; He, X. Aerobic denitrification affects gaseous nitrogen loss in biofloc-based recirculating aquaculture system. Aquaculture 2020, 529, 735686. [Google Scholar] [CrossRef]
- Yao, R.; Yuan, Q.; Wang, K. Enrichment of denitrifying bacterial community using nitrite as an electron acceptor for nitrogen removal from wastewater. Water 2020, 12, 48. [Google Scholar] [CrossRef] [Green Version]
- Lycus, P.; Lovise Bothun, K.; Bergaust, L.; Peele Shapleigh, J.; Bakken, L.R.; Frostegård, Å. Phenotypic and genotypic richness of denitrifiers revealed by a novel isolation strategy. ISME J. 2017, 11, 2219–2232. [Google Scholar] [CrossRef] [Green Version]
- Rocco, C.A.; Bergaust, L.L.; Bakken, L.R.; Yavitt, J.B.; Shapleigh, J.P. Modularity of nitrogen-oxide reducing soil bacteria: Linking phenotype to genotype. Environ. Microbiol. 2017, 19, 2507–2519. [Google Scholar] [CrossRef] [PubMed]
- Kraft, B.; Strous, M.; Tegetmeyer, H.E. Microbial nitrate respiration--genes, enzymes and environmental distribution. J. Biotechnol. 2011, 155, 104–117. [Google Scholar] [CrossRef] [PubMed]
- Hallin, S.; Philippot, L.; Löffler, F.E.; Sanford, R.A.; Jones, C.M. Genomics and ecology of novel N2O-reducing microorganisms. Trends Microbiol. 2018, 26, 43–55. [Google Scholar] [CrossRef]
- Frostegård, Å.; Vick, S.H.W.; Lim, N.Y.M.; Bakken, L.R.; Shapleigh, J.P. Linking meta-omics to the kinetics of denitrification intermediates reveals pH-dependent causes of N2O emissions and nitrite accumulation in soil. bioRxiv 2020. [Google Scholar] [CrossRef]
- Chon, K.; Chang, J.-S.; Lee, E.; Lee, J.; Ryu, J.; Cho, J. Abundance of denitrifying genes coding for nitrate (narG), nitrite (nirS), and nitrous oxide (nosZ) reductases in estuarine versus wastewater effluent-fed constructed wetlands. Ecol. Eng. 2011, 37, 64–69. [Google Scholar] [CrossRef]
- Liao, R.; Miao, Y.; Li, J.; Li, Y.; Wang, Z.; Du, J.; Li, Y.; Li, A.; Shen, H. Temperature dependence of denitrification microbial communities and functional genes in an expanded granular sludge bed reactor treating nitrate-rich wastewater. RSC Adv. 2018, 8, 42087–42094. [Google Scholar] [CrossRef] [Green Version]
- Venterea, R.T. Nitrite-driven nitrous oxide production under aerobic soil conditions: Kinetics and biochemical controls. Glob. Chang. Biol. 2007, 13, 1798–1809. [Google Scholar] [CrossRef]
- Warneke, S.; Schipper, L.A.; Bruesewitz, D.A.; Baisden, W.T. A comparison of different approaches for measuring denitrification rates in a nitrate removing bioreactor. Water Res. 2011, 45, 4141–4151. [Google Scholar] [CrossRef] [PubMed]
- Warneke, S.; Schipper, L.A.; Bruesewitz, D.A.; McDonald, I.; Cameron, S. Rates, controls and potential adverse effects of nitrate removal in a denitrification bed. Ecol. Eng. 2011, 37, 511–522. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jéglot, A.; Sørensen, S.R.; Schnorr, K.M.; Plauborg, F.; Elsgaard, L. Temperature Sensitivity and Composition of Nitrate-Reducing Microbiomes from a Full-Scale Woodchip Bioreactor Treating Agricultural Drainage Water. Microorganisms 2021, 9, 1331. https://doi.org/10.3390/microorganisms9061331
Jéglot A, Sørensen SR, Schnorr KM, Plauborg F, Elsgaard L. Temperature Sensitivity and Composition of Nitrate-Reducing Microbiomes from a Full-Scale Woodchip Bioreactor Treating Agricultural Drainage Water. Microorganisms. 2021; 9(6):1331. https://doi.org/10.3390/microorganisms9061331
Chicago/Turabian StyleJéglot, Arnaud, Sebastian Reinhold Sørensen, Kirk M. Schnorr, Finn Plauborg, and Lars Elsgaard. 2021. "Temperature Sensitivity and Composition of Nitrate-Reducing Microbiomes from a Full-Scale Woodchip Bioreactor Treating Agricultural Drainage Water" Microorganisms 9, no. 6: 1331. https://doi.org/10.3390/microorganisms9061331
APA StyleJéglot, A., Sørensen, S. R., Schnorr, K. M., Plauborg, F., & Elsgaard, L. (2021). Temperature Sensitivity and Composition of Nitrate-Reducing Microbiomes from a Full-Scale Woodchip Bioreactor Treating Agricultural Drainage Water. Microorganisms, 9(6), 1331. https://doi.org/10.3390/microorganisms9061331