
Two Lysine Sites That Can Be Malonylated Are Important for LuxS Regulatory Roles in Bacillus velezensis



Abstract
:1. Introduction
2. Materials and Methods
3. Results
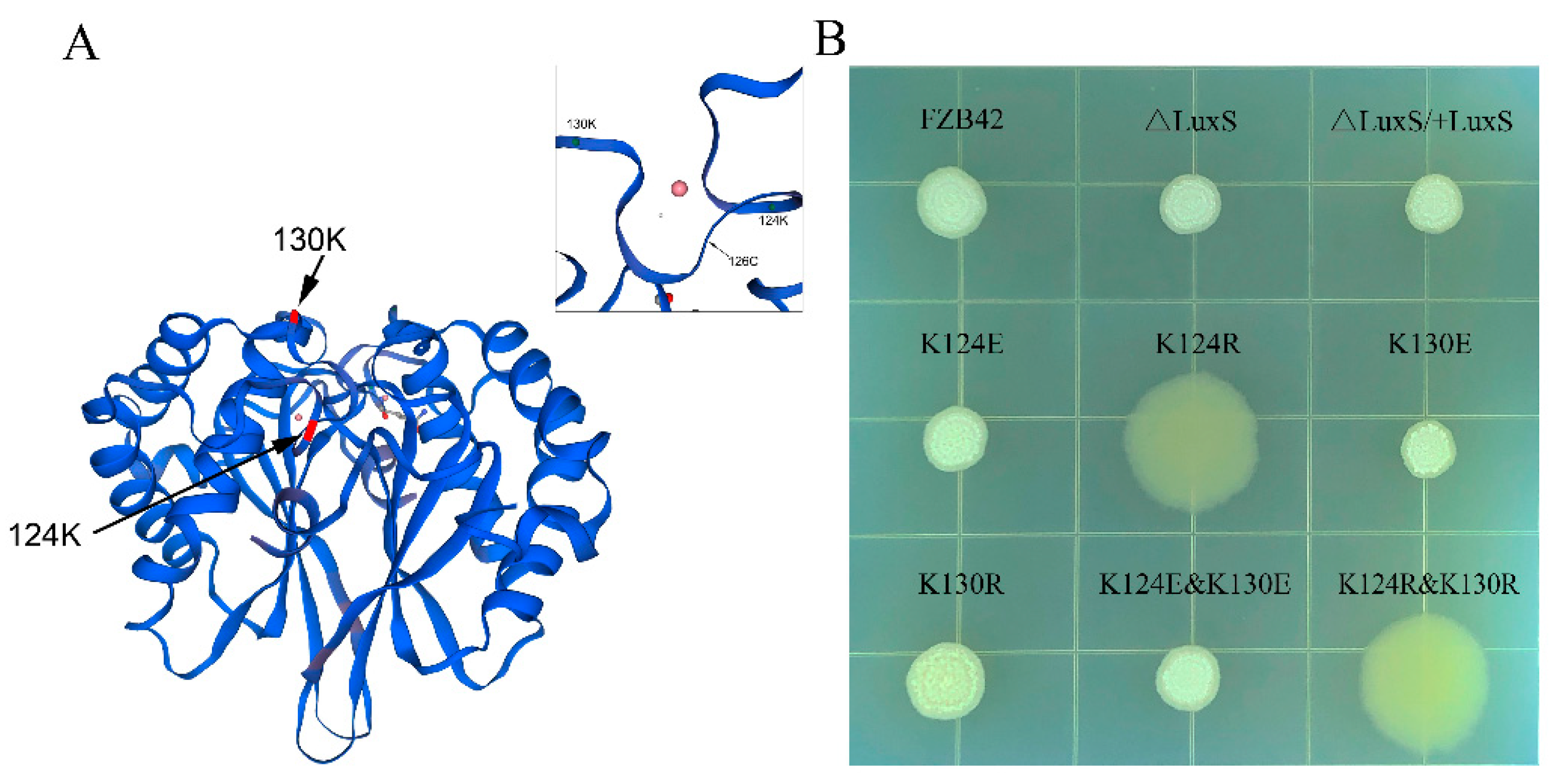
3.1. Construction of Mutants
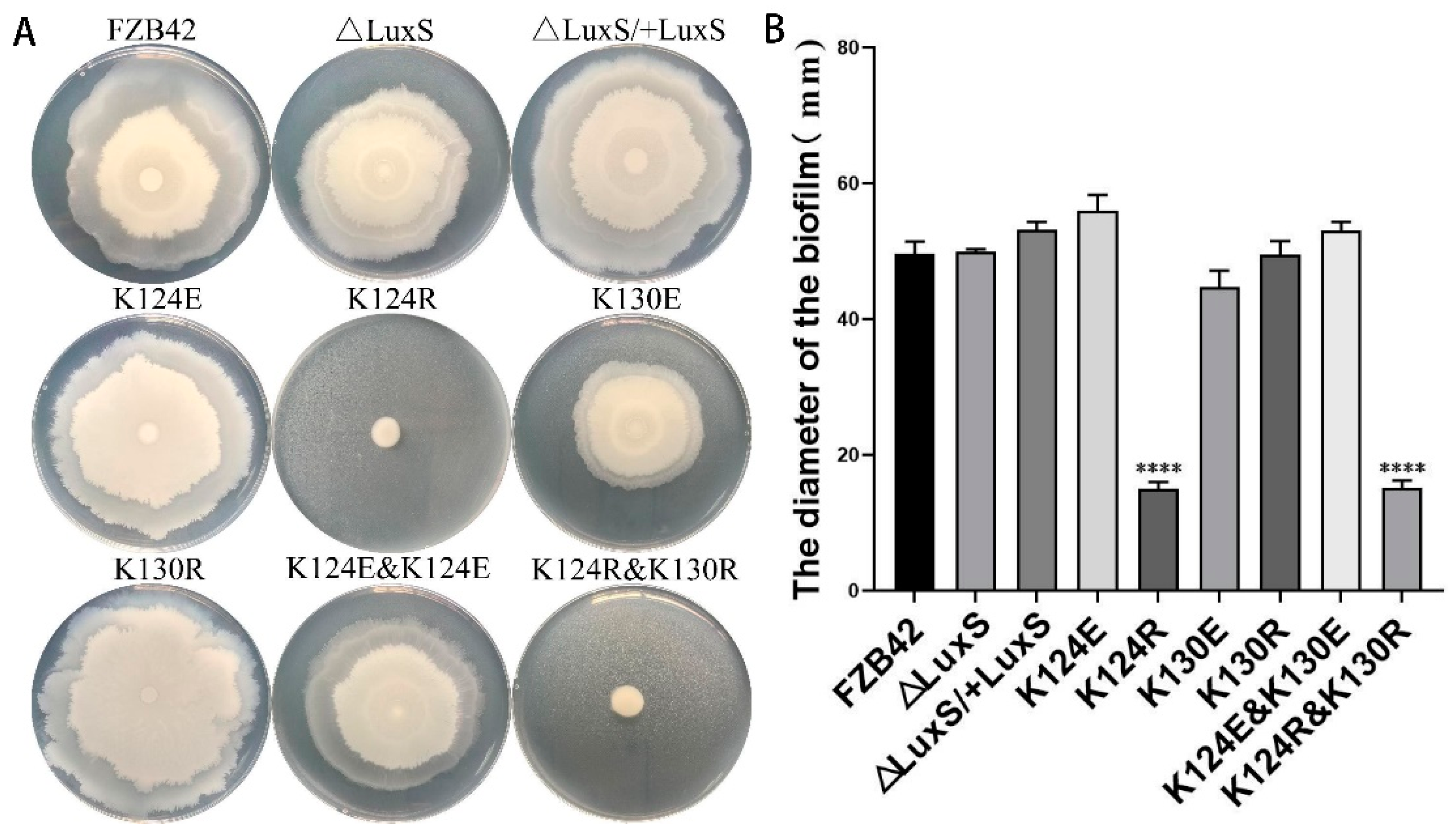
3.2. K124 of LuxS Was Critical for Biofilm Formation of FZB42
3.3. Mutations Compromised FZB42 Swarming and Sporulation
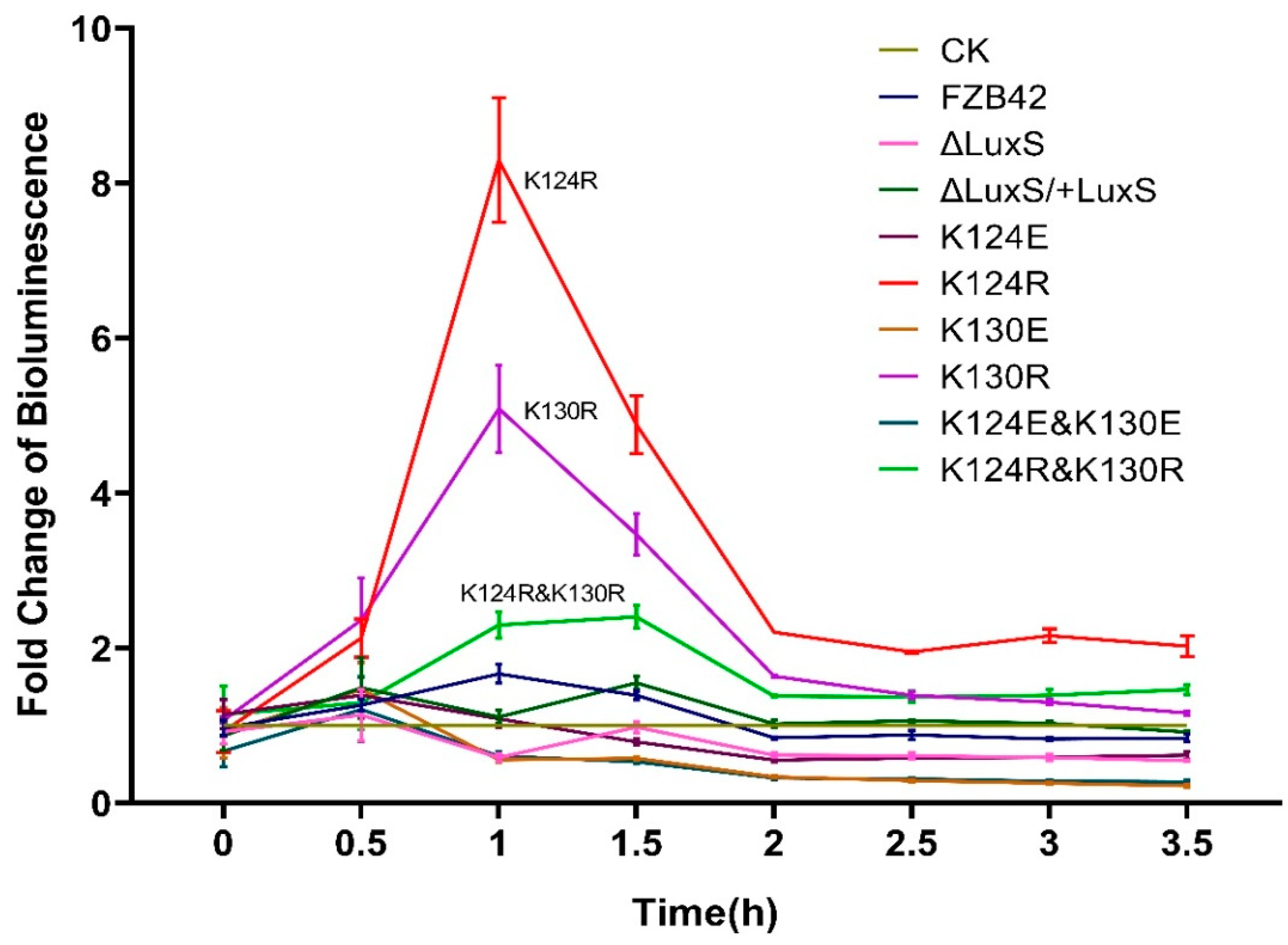
3.4. K124R and K130R Induced AI-2 Production
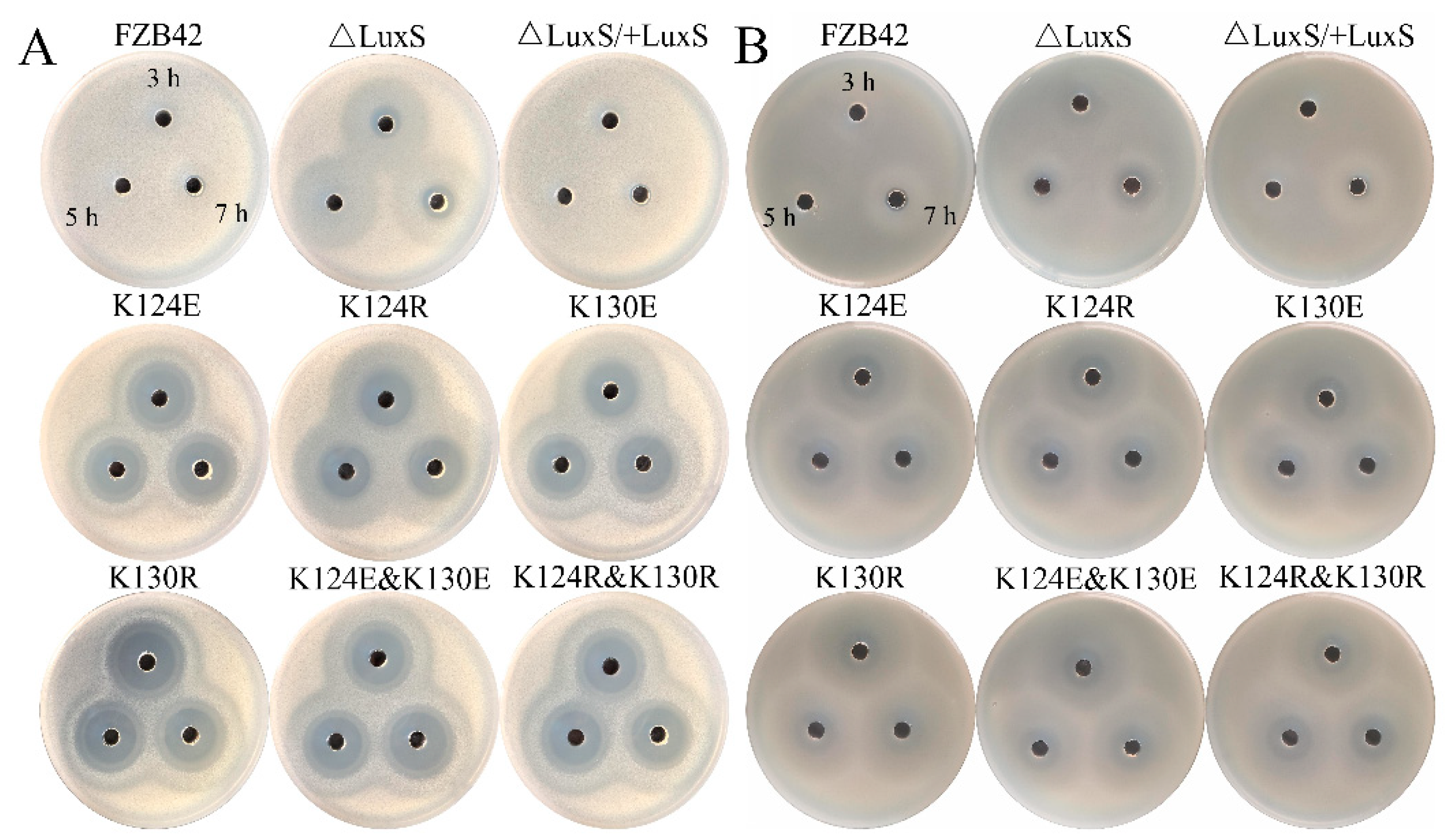
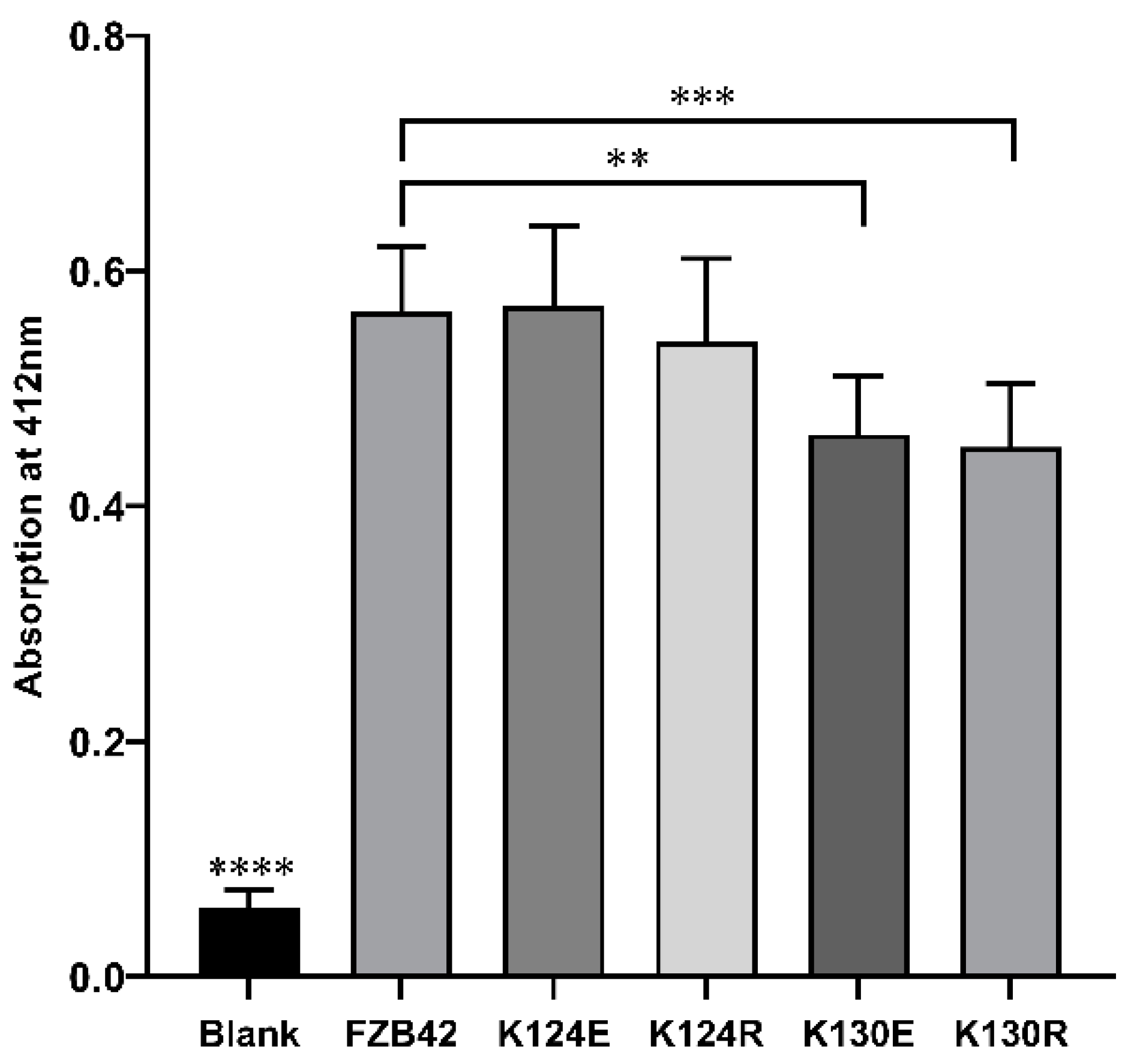
3.5. Mutations Enhanced Antibiotic Production
3.6. K130 Mutations Affected in vitro Enzymatic Activity of LuxS
3.7. K130 Is Highly Conserved in Bacillus Species
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fan, B.; Li, Y.L.; Li, L.; Peng, X.J.; Bu, C.; Wu, X.Q.; Borriss, R. Malonylome analysis of rhizobacterium Bacillus amyloliquefaciens FZB42 reveals involvement of lysine malonylation in polyketide synthesis and plant-bacteria interactions. J. Proteom. 2017, 154, 1–12. [Google Scholar]
- Park, J.; Chen, Y.; Tishkoff, D.X.; Peng, C.; Tan, M.; Dai, L.; Xie, Z.; Zhang, Y.; Zwaans, B.M.; Skinner, M.E. SIRT5-mediated lysine desuccinylation impacts diverse metabolic pathways. Mol. Cell 2013, 50, 919–930. [Google Scholar]
- Chao, P.; Lu, Z.; Xie, Z.; Cheng, Z.; Zhao, Y. The first identification of lysine malonylation substrates and its regulatory enzyme. Mol. Cell Proteom. 2011, 10, M111-012658. [Google Scholar]
- Nishida, Y.; Rardin, M.; Carrico, C.; He, W.; Sahu, A.; Gut, P.; Najjar, R.; Fitch, M.; Hellerstein, M.; Gibson, B. SIRT5 Regulates both Cytosolic and Mitochondrial Protein Malonylation with Glycolysis as a Major Target. Mol. Cell 2015, 59, 321–332. [Google Scholar]
- Colak, G.; Pougovkina, O.; Dai, L.; Tan, M.; Brinke, H.T.; Huang, H.; Cheng, Z.; Park, J.; Wan, X.; Liu, X. Proteomic and Biochemical Studies of Lysine Malonylation Suggest Its Malonic Aciduria-associated Regulatory Role in Mitochondrial Function and Fatty Acid Oxidation. Mol. Cell Proteom. 2015, 14, 3056–3071. [Google Scholar]
- Fan, B. Bacillus velezensis FZB42 in 2018: The Gram-Positive Model Strain for Plant Growth Promotion and Biocontrol. Front. Microbiol. 2018, 9, 2491. [Google Scholar]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Eisenreich, A.; Schneider, K.; Heinemeyer, I.; Morgenstern, B.; Voss, B.; Hess, W.R.; Reva, O. Comparative analysis of the complete genome sequence of the plant growth-promoting bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 2007, 25, 1007–1014. [Google Scholar]
- Fan, B.; Chen, X.H.; Budiharjo, A.; Bleiss, W.; Vater, J.; Borriss, R. Efficient colonization of plant roots by the plant growth promoting bacterium Bacillus amyloliquefaciens FZB42, engineered to express green fluorescent protein. J. Biotechnol 2011, 151, 303–311. [Google Scholar]
- Pei, D.; Zhu, J. Mechanism of action of S-ribosylhomocysteinase (LuxS). Curr. Opin. Chem. Biol. 2004, 8, 492–497. [Google Scholar]
- Zhu, J.; Patel, R.; Pei, D. Catalytic Mechanism of S -Ribosylhomocysteinase (LuxS): Stereochemical Course and Kinetic Isotope Effect of Proton Transfer Reactions. Biochemistry 2004, 43, 10166–10172. [Google Scholar]
- Palmer, J.L.; Abeles, R.H. The mechanism of action of S-adenosylhomocysteinase. J. Biol. Chem. 1979, 254, 1217–1226. [Google Scholar]
- Winans, S.C. Bacterial Esperanto. Nat. Struct. Biol. 2002, 9, 83–84. [Google Scholar]
- Xavier, K.B.; Bassler, B.L. LuxS quorum sensing: More than just a numbers game. Curr. Opin. Microbiol. 2003, 6, 191–197. [Google Scholar]
- Vendeville, A.; Winzer, K.; Heurlier, K.; Tang, C.; Hardie, K. Making ‘sense’ of metabolism: Autoinducer-2, LuxS and pathogenic bacteria. Nat. Rev. Microbiol. 2005, 3, 383–396. [Google Scholar]
- Schauder, S.; Bassler, B.L. The languages of bacteria. Genes Dev. 2001, 15, 1468–1480. [Google Scholar]
- Mcnab, R.; Lamont, R.J. Microbial dinner-party conversations: The role of LuxS in interspecies communication. J. Med. Microbiol. 2003, 52, 541–545. [Google Scholar]
- Stevenson, B.; Babb, K. LuxS-mediated quorum sensing in Borrelia burgdorferi, the Lyme disease spirochete. Infect. Immun. 2002, 70, 4099–4105. [Google Scholar]
- Miller, S.T.; Xavier, K.B.; Campagna, S.R.; Taga, M.E.; Hughson, F.M. Salmonella typhimurium recognizes a chemically distinct form of the bacterial quorum-sensing signal AI-2. Mol. Cell 2004, 15, 677–687. [Google Scholar]
- Miranda, V.; Torcato, I.M.; Xavier, K.B.; Ventura, M.R. Synthesis of d-desthiobiotin-AI-2 as a novel chemical probe for autoinducer-2 quorum sensing receptors. Bioorg Chem. 2019, 92, 103200. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.S.; Regt, A.; Brito, P.H.; Miller, S.T.; Xavier, K.B. Identification of functional LsrB-like autoinducer-2 receptors. J. Bacteriol. 2009, 191, 6975. [Google Scholar]
- Torcato, I.M.; Kasal, M.R.; Brito, P.H.; Miller, S.T.; Xavier, K.B. Identification of novel autoinducer-2 receptors in Clostridia reveals plasticity in the binding site of the LsrB receptor family. J. Biol. Chem. 2019, 294, 4450–4463. [Google Scholar]
- Pereira, C.S.; Mcauley, J.R.; Taga, M.E.; Xavier, K.B.; Miller, S.T. Sinorhizobium meliloti, a bacterium lacking the autoinducer-2 (AI-2) synthase, responds to AI-2 supplied by other bacteria. Mol. Microbiol. 2010, 70, 1223–1235. [Google Scholar]
- Kavanaugh, J.S.; Gakhar, L.; Horswill, A.R. The structure of LsrB from Yersinia pestis complexed with autoinducer-2. Acta Crystallogr. 2011, 67. [Google Scholar]
- Chen, X.; Schauder, S.; Potier, N.; Van Dorsselaer, A.; Pelczer, I.; Bassler, B.L.; Hughson, F.M. Structural identification of a bacterial quorum-sensing signal containing boron. Nature 2002, 415, 545–549. [Google Scholar]
- Zhang, L.; Li, S.; Liu, X.; Wang, Z.; Shen, X. Sensing of autoinducer-2 by functionally distinct receptors in prokaryotes. Nat. Commun. 2020, 11, 1–13. [Google Scholar]
- Lombardia, E.; Rovetto, A.J.; Arabolaza, A.L.; Grau, R.R. A LuxS-dependent cell-to-cell language regulates social behavior and development in Bacillus subtilis. J. Bacteriol. 2006, 188, 4442–4452. [Google Scholar]
- Lebeer, S. Functional analysis of luxS in the probiotic strain Lactobacillus rhamnosus GG reveals a central metabolic role important for growth and Biofilm formation. J. Bacteriol. 2007, 189, 860–871. [Google Scholar]
- Duanis-Assaf, D.; Steinberg, D.; Chai, Y.; Shemesh, M. The LuxS Based Quorum Sensing Governs Lactose Induced Biofilm Formation by Bacillus subtilis. Front. Microbiol. 2016, 6, 1517. [Google Scholar]
- Pereira, C.S.; Thompson, J.A.; Xavier, K.B. AI-2-mediated signalling in bacteria. Fems Microbiol. Rev. 2013, 2, 156–181. [Google Scholar]
- Hilgers, M.T.; Ludwig, M.L. Crystal structure of the quorum-sensing protein LuxS reveals a catalytic metal site. Proc. Natl. Acad. Sci. USA 2001, 98, 11169–11174. [Google Scholar]
- Andrew, W.; Martino, B.; Stefan, B.; Gabriel, S.; Gerardo, T.; Rafal, G.; Heer, F.T.; de Beer, A.P.; Chriatine, R.; Lorenza, B. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, W1, W296–W303. [Google Scholar]
- Idris, E.; Iglesias, D.J.; Talon, M.; Borriss, R. Tryptophan-Dependent Production of Indole-3-Acetic Acid (IAA) Affects Level of Plant Growth Promotion by Bacillus amyloliquefaciens FZB42. Mol. Plant Microbe Interact. MPMI 2007, 20, 619. [Google Scholar]
- Pascale, J.; Jean-Raphael, F.; Marie-Laure, H.; François, D. Rapid orientated cloning in a shuttle vector allowing modulated gene expression in Bacillus subtilis. Fems Microbiol. Lett. 2001, 205, 91–97. [Google Scholar]
- Schauder, S.; Shokat, K.; Surette, M.G.; Bassler, B.L. The LuxS family of bacterial autoinducers: Biosynthesis of a novel quorum-sensing signal molecule. Mol. Microbiol. 2001, 41, 463–476. [Google Scholar]
- Wu, L.; Wu, H.; Chen, L.; Yu, X.; Borriss, R.; Gao, X. Difficidin and bacilysin from Bacillus amyloliquefaciens FZB42 have antibacterial activity against Xanthomonas oryzae rice pathogens. Sci. Rep. 2015, 5, 12975. [Google Scholar]
- Wu, L.; Wu, H.; Chen, L.; Xie, S.; Zang, H.; Borriss, R.; Gao, X. Bacilysin from Bacillus amyloliquefaciens FZB42 has specific bactericidal activity against harmful algal bloom species. Appl. Environ. Microbiol. 2014, 80, 7512–7520. [Google Scholar]
- Chen, X.H.; Scholz, R.; Borriss, M.; Junge, H.; M?Gel, G.; Kunz, S.; Borriss, R. Difficidin and bacilysin produced by plant-associated Bacillus amyloliquefaciens are efficient in controlling fire blight disease. J. Biotechnol. 2009, 140, 38–44. [Google Scholar]
- Chen, X.H.; Vater, J.; Piel, J.; Franke, P.; Scholz, R.; Schneider, K.; Koumoutsi, A.; Hitzeroth, G.; Grammel, N.; Strittmatter, M.A.W. Structural and functional characterization of three polyketide synthase gene clusters in Bacillus amyloliquefaciens FZB42. J. Bacteriol. 2006, 188, 4024–4036. [Google Scholar]
- Mariappan, A.; Makarewicz, O.; Chen, X.H.; Borriss, R. Two-component response regulator DegU controls the expression of bacilysin in plant-growth-promoting bacterium Bacillus amyloliquefaciens FZB42. J. Mol. Microbiol. Biotechnol. 2012, 22, 114–125. [Google Scholar]
- Vlamakis, H.; Chai, Y.; Beauregard, P.; Losick, R.; Kolter, R. Sticking together: Building a biofilm the Bacillus subtilis way. Nature reviews. Microbiology 2013, 11, 157–168. [Google Scholar]
- Xiong, Q.; Liu, D.; Zhang, H.; Dong, X.; Zhang, G.; Liu, Y.; Zhang, R. Quorum sensing signal autoinducer-2 promotes root colonization of Bacillus velezensis SQR9 by affecting biofilm formation and motility. Appl. Microbiol. Biotechnol. 2020, 104, 7177–7185. [Google Scholar] [CrossRef] [PubMed]
- Ju, X.Y.; Li, J.K.; Zhu, M.J.; Lv, F.X. Effect of the luxS gene on biofilm formation and antibiotic resistance by Salmonella serovar Dublin. Food Res. Int. 2018, 107, 385–393. [Google Scholar]
- Hammer, B.K.; Bassler, B.L. Quorum sensing controls biofilm formation in Vibrio cholerae. Mol. Microbiol. 2003, 50, 101–114. [Google Scholar]
- Ren, D.; Sims, J.J.; Wood, T.K. Inhibition of biofilm formation and swarming of Bacillus subtilis by (5Z)-4-bromo-5-(bromomethylene)-3-butyl-2(5H)-furanone. Lett. Appl. Microbiol. 2002, 34, 293–299. [Google Scholar]
- Auger, S.; Krin, E.; Aymerich, S.; Gohar, M. Autoinducer 2 Affects Biofilm Formation by Bacillus cereus. Appl. Environ. Microbiol. 2006, 72, 937–941. [Google Scholar]
- Sun, J.; Daniel, R.; Wagner-D Bler, I.; Zeng, A.P. Is autoinducer-2 a universal signal for interspecies communication: A comparative genomic and phylogenetic analysis of the synthesis and signal transduction pathways. BMC Evol. Biol. 2004, 4, 1–11. [Google Scholar]
- Rezzonico, F.; Duffy, B. Lack of genomic evidence of AI-2 receptors suggests a non-quorum sensing role for luxS in most bacteria. BMC Microbiol. 2008, 8, 1–9. [Google Scholar]
- Min, J.; Shao, W.; Perego, M.; Hoch, J.A. Multiple histidine kinases regulate entry into stationary phase and sporulation in Bacillus subtilis. Mol. Microbiol. 2000, 38, 535–542. [Google Scholar]
- Aguilar, C.; Vlamakis, H.; Guzman, A.; Losick, R.; Kolter, R. KinD Is a Checkpoint Protein Linking Spore Formation to Extracellular-Matrix Production in Bacillus subtilis Biofilms. Mbio 2010, 1, e00035-10. [Google Scholar]
- Fan, B.; Li, Y.L.; Mariappan, A.; Becker, A.; Wu, X.Q. New SigD-regulated genes identified in the rhizobacterium Bacillus amyloliquefaciens FZB42. Biol. Open 2016, 5, 1776–1783. [Google Scholar]
- Paul, C.S.; Anton, H.; Gao, X.W.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42—A review. Front. Microbiol. 2015, 6, 780. [Google Scholar]
- Koumoutsi, A.; Chen, X.H.; Vater, J.; Borriss, R. DegU and YczE Positively Regulate the Synthesis of Bacillomycin D by Bacillus amyloliquefaciens Strain FZB42. Appl. Environ. Microbiol. 2007, 73, 6953–6964. [Google Scholar]
- Liu, Y.; Nan, Z.; Qiu, M.; Feng, H.; Vivanco, J.M.; Shen, Q.; Zhang, R. Enhanced rhizosphere colonization of beneficial Bacillus amyloliquefaciens SQR9 by pathogen infection. Fems Microbiol. Lett. 2014, 353, 49–56. [Google Scholar]
- Fazle Rabbee, M.; Baek, K.-H. Antimicrobial Activities of Lipopeptides and Polyketides of Bacillus velezensis for Agricultural Applications. Molecules 2020, 25, 4973. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Materials | Description or Sequence (5′–3′) | Source |
|---|---|---|
| Strain | ||
| FZB42 | Bacillus velezensis FZB42 wild type | Lab stock |
| FBS119 | FZB42, luxS::speR | This study |
| FBS121 | FZB42, amyE::luxS + emR, luxS::speR | This study |
| FBS122 | FZB42, amyE::luxS (K124R) + emR, luxS::speR | This study |
| FBS123 | FZB42, amyE::luxS (K124E) + emR, luxS::speR | This study |
| FBS124 | FZB42, amyE::luxS (K130R) + emR, luxS::speR | This study |
| FBS125 | FZB42, amyE::luxS (K130E) + emR, luxS::speR | This study |
| FBS126 | FZB42, amyE::luxS (K124R,K130R) + emR, luxS::speR | This study |
| FBS127 | FZB42, amyE::luxS (K124E,K130E) + emR, luxS::speR | This study |
| V. harveyi BB170 | luxN::Tn5 | BNCC |
| B. megaterium | Lab stock | |
| S. aureus ATCC9144 | bacilysin sensitive | ATCC |
| B. subtilis 168 | trpC2 | BGSC |
| FBS385 | B. subtilis 168 with pDG148-Stu-luxS (K130E) | This study |
| FBS386 | B. subtilis 168 with pDG148-Stu-luxS (K130R) | This study |
| FBS387 | B. subtilis 168 with pDG148-Stu-luxS | This study |
| FBS388 | B. subtilis 168 with pDG148-Stu-luxS (K124E) | This study |
| FBS389 | B. subtilis 168 with pDG148-Stu-luxS (K124R) | This study |
| Plasmids | ||
| pFB01 | amyE::gfp + emR (Ampr Emr) | [8] |
| pMD-19 | Commercial T-Vector (Ampr) | Takara |
| pFB68 | pDG148-Stu (Ampr Kmr) | BGSC |
| pFB106 | pMD-19-luxS-speR (Ampr Spcr) | This study |
| pFB108 | pFB01-luxS (Ampr Emr) | This study |
| pFB109 | pFB01-luxS(K124R) (Ampr Emr) | This study |
| pFB110 | pFB01-luxS(K124E) (Ampr Emr) | This study |
| pFB144 | pFB01-luxS(K130R) (Ampr Emr) | This study |
| pFB145 | pFB01-luxS(K130E) (Ampr Emr) | This study |
| pFB487 | pDG148-Stu-luxS-K130E (Ampr Kmr) | This study |
| pFB488 | pDG148-Stu-luxS-K130R (Ampr Kmr) | This study |
| pFB489 | pDG148-Stu-luxS (Ampr Kmr) | This study |
| pFB490 | pDG148-Stu-luxS-K124E (Ampr Kmr) | This study |
| pFB491 | pDG148-Stu-luxS-K124R (Ampr Kmr) | This study |
| Primers | ||
| FBO-335 | ATAATCCACAGCAGGTA | |
| FBO-336 | TTGAACAATCACGAAAC | |
| FBO-445 | ATACCAAACATCTAAATTCCCGG | |
| FBO-449 | CGATCACTTCGACATCATAGATA | |
| FBO-450 | GCAGAAGCGAATGTCAAACTTAT | |
| FBO-453 | TTGTTCTGCGCTCTCATTGC | |
| FBO-465 | ATAATCGATCTTCCGCCACAATTCTTA (ClaI restriction site) | |
| FBO-495 | ATACTCGAGAGCATACCGCACATACCT (AvaI restriction site) | |
| FBO-516 | ATCATGAAGTCTCGCCTGGCC | |
| FBO-517 | ATCATGAAGTTCCGCCTGGCC | |
| FBO-518 | TTAGAAGGCGCGAAACGTCTGAT | |
| FBO-545 | GCGGCCAACGAAAGACAGTGC | |
| FBO-546 | GCGGCCAACGAAGAACAGTGC | |
| FBO-547 | CGGAATCTCCGTAATATCGAT | |
| FBO-1371 | AAGGAGGAAGCAGGTATGCATCACCATCACCATCACGGATCAATGCCTTCATAGAAAGTTTTGAG (His-tag) | |
| FBO-1372 | GACACGCACGAGGTTTATCCGAACACTTTCAGCAAATC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, X.; Li, Y.; Fan, J.; Zhao, Y.; Borriss, R.; Fan, B. Two Lysine Sites That Can Be Malonylated Are Important for LuxS Regulatory Roles in Bacillus velezensis. Microorganisms 2021, 9, 1338. https://doi.org/10.3390/microorganisms9061338
Cao X, Li Y, Fan J, Zhao Y, Borriss R, Fan B. Two Lysine Sites That Can Be Malonylated Are Important for LuxS Regulatory Roles in Bacillus velezensis. Microorganisms. 2021; 9(6):1338. https://doi.org/10.3390/microorganisms9061338
Chicago/Turabian StyleCao, Xianming, Yulong Li, Jialu Fan, Yinjuan Zhao, Rainer Borriss, and Ben Fan. 2021. "Two Lysine Sites That Can Be Malonylated Are Important for LuxS Regulatory Roles in Bacillus velezensis" Microorganisms 9, no. 6: 1338. https://doi.org/10.3390/microorganisms9061338
APA StyleCao, X., Li, Y., Fan, J., Zhao, Y., Borriss, R., & Fan, B. (2021). Two Lysine Sites That Can Be Malonylated Are Important for LuxS Regulatory Roles in Bacillus velezensis. Microorganisms, 9(6), 1338. https://doi.org/10.3390/microorganisms9061338