
Plasmodium matutinum Causing Avian Malaria in Lovebirds (Agapornis roseicollis) Hosted in an Italian Zoo

 , , , and
, , , and 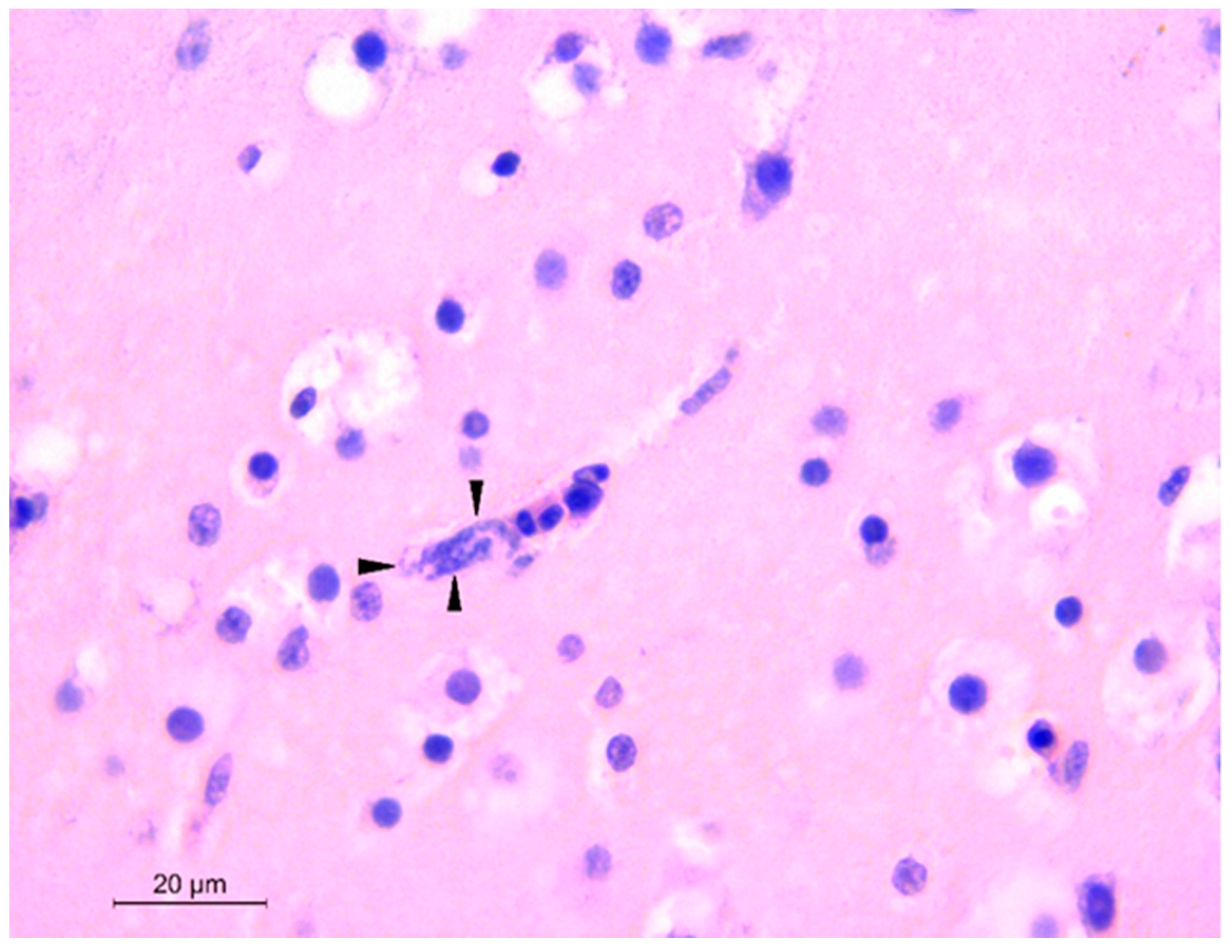{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Collection
2.2. Pathology, Histopathology, and Bacteriology
2.3. Haemosporidians Molecular Identification and Characterization
3. Results
3.1. Pathology, Histopathology, and Bacteriology
3.2. Haemosporidians Identification and Characterization
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olias, P.; Wegelin, M.; Zenker, W.; Freter, S.; Gruber, A.D.; Klopfleisch, R. Avian malaria deaths in parrots, Europe. Emerg. Infect. Dis. 2011, 17, 950–952. [Google Scholar] [CrossRef]
- Nourani, L.; Zakeri, S.; Dinparast Djadid, N. Dynamics of prevalence and distribution pattern of avian Plasmodium species and its vectors in diverse zoogeographical areas—A review. Infect. Genet. Evol. 2020, 8, 1042–1044. [Google Scholar] [CrossRef] [PubMed]
- Harl, J.; Himmel, T.; Valkiūnas, G.; Ilgūnas, M.; Bakonyi, T.; Weissenböck, H. Geographic and host distribution of haemosporidian parasite lineages from birds of the family Turdidae. Malar. J. 2020, 19, 335. [Google Scholar] [CrossRef]
- Valkiūnas, G. Avian Malaria Parasites and Other Haemosporidia; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Glaizot, O.; Fumagalli, L.; Iritano, K.; Lalubin, F.; Van Rooyen, J.; Christe, P. High prevalence and lineage diversity of avian malaria in wild populations of great tits (Parus major) and mosquitoes (Culex pipiens). PLoS ONE 2012, 7, e34964. [Google Scholar] [CrossRef] [PubMed]
- Lalubin, F.; Delédevant, A.; Glaizot, O.; Christe, P. Natural malaria infection reduces starvation resistance of nutritionally stressed mosquitoes. J. Anim. Ecol. 2014, 83, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Martínez-de la Puente, J.M.; Muñoz, J.; Capelli, G.; Montarsi, F.; Soriguer, R.; Arnoldi, D.; Rizzoli, A.; Figuerola, J. Avian malaria parasites in the last supper: Identifying encounters between parasites and the invasive Asian mosquito tiger and native mosquito species in Italy. Malar. J. 2015, 14, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Valkiūnas, G.; Iezhova, T.A. Keys to the avian malaria parasites. Malar. J. 2018, 29, 212. [Google Scholar] [CrossRef] [PubMed]
- Waldenström, J.; Bensch, S.; Hasselquist, D.; Ostman, O. A new nested polymerase chain reaction method very efficient in detecting Plasmodium and Haemoproteus infections from avian blood. J. Parasitol. 2004, 90, 191–194. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Zehtindjiev, P.; Dimitrov, D.; Križanauskienė, A.; Iezhova, T.A.; Bensch, S. Polymerase chain reaction-based identification of Plasmodium (Huffia) elongatum, with remarks on species identity of haemosporidian lineages deposited in GenBank. Parasitol. Res. 2008, 102, 1185–1193. [Google Scholar] [CrossRef]
- Vanstreels, R.E.T.; Dutra, D.A.; Ferreira-Junior, F.C.; Hurtado, R.; Egert, L.; Mayorga, L.F.S.P.; Catão-Dias, J.L. Epidemiology, hematology, and unusual morphological characteristics of Plasmodium during an avian malaria outbreak in penguins in Brazil. Parasitol. Res. 2019, 118, 3497–3508. [Google Scholar] [CrossRef]
- Spottiswoode, N.; Bartlett, S.L.; Conley, K.J.; Seimon, T.A.; Griffin, D.O.; Sykes, J.M., 4th. Analysis of Plasmodium lineages identified in captive penguins (Sphenisciformes spp.), eiders (Somateria spp.), and inca terns (Larosterna inca) in a North American zoological collection. J. Zoo. Wildl. Med. 2020, 51, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Braga, E.M.; Silveira, P.; Belo, N.O.; Valkiūnas, G. Recent advances in the study of avian malaria: An overview with an emphasis on the distribution of Plasmodium spp in Brazil. Mem. Inst. Oswaldo Cruz. 2011, 106 (Suppl. 1), 3–11. [Google Scholar] [CrossRef]
- Hoque, M.A.; Burgess, G.W.; Karo-Karo, D.; Cheam, A.L.; Skerratt, L.F. Monitoring of wild birds for Newcastle disease virus in north Queensland, Australia. Prev. Vet. Med. 2012, 103, 49–62. [Google Scholar] [CrossRef] [PubMed]
- LaPointe, D.A.; Atkinson, C.T.; Samuel, M.D. Ecology and conservation biology of avian malaria. Ann. N. Y. Acad. Sci. 2012, 1249, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Asghar, M.; Hasselquist, D.; Hansson, B.; Zehtindjiev, P.; Westerdahl, H.; Bensch, S. Chronic infection. Hidden costs of infection: Chronic malaria accelerates telomere degradation and senescence in wild birds. Science 2015, 23, 436–438. [Google Scholar] [CrossRef] [PubMed]
- Grilo, M.L.; Vanstreels, R.E.; Wallace, R.; García-Párraga, D.; Braga, É.M.; Chitty, J.; Catão-Dias, J.L.; Madeira de Carvalho, L.M. Malaria in penguins—Current perceptions. Avian. Pathol. 2016, 45, 393–407. [Google Scholar] [CrossRef] [Green Version]
- Vanstreels, R.E.; Braga, É.M.; Catão-Dias, J.L. Blood parasites of penguins: A critical review. Parasitology 2016, 143, 931–956. [Google Scholar] [CrossRef]
- Magallanes, S.; García-Longoria, L.; López-Calderón, C.; Reviriego, M.; de Lope, F.; Møller, A.P.; Marzal, A. Uropygial gland volume and malaria infection are related to survival in migratory house martins. J. Avian. Biol. 2017, 48, 1355–1359. [Google Scholar] [CrossRef]
- Muriel, J.; Graves, J.A.; Gil, D.; Magallanes, S.; Salaberria, C.; Casal-López, M.; Marzal, A. Molecular characterization of avian malaria in the spotless starling (Sturnus unicolor). Parasitol. Res. 2018, 117, 919–928. [Google Scholar] [CrossRef]
- Bennett, G.F.; Bishop, M.A.; Peirce, M.A. Checklist of the avian species of Plasmodium Marchiafava & Celli, 1885 (Apicomplexa) and their distribution by avian family and Wallacean life zones. Syst. Parasitol. 1993, 26, 171–179. [Google Scholar]
- Verwey, J.K.; Peters, A.; Monks, D.; Raidal, S.R. Spillover of avian haemosporidian parasites (Haemosporidia: Plasmodium) and death of captive psittacine species. Aust. Vet. J. 2018, 96, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Masello, J.F.; Martínez, J.; Calderón, L.; Wink, M.; Quillfeldt, P.; Sanz, V.; Theuerkauf, J.; Ortiz-Catedral, L.; Berkunsky, I.; Brunton, D.; et al. Can the intake of antiparasitic secondary metabolites explain the low prevalence of hemoparasites among wild Psittaciformes? Parasites Vectors 2018, 11, 357. [Google Scholar] [CrossRef] [Green Version]
- Corradetti, A.; Morcos, W.M.; Neri, I. Sulle produzioni di forme endoistiocitare nei canarini infettati con il ceppo italiano di Plasmodium matutinum derivato da Turdus iliacus. Parassitologia 1962, 4, 105–108. [Google Scholar]
- Corradetti, A.; Verolini, F.; Neri, I. Plasmodium (Haemamoeba) giovannolai n. sp. parassita di Turdus merula. Parassitologia 1963, 5, 11–18. [Google Scholar]
- Zehtindjiev, P.; Križanauskienė, A.; Scebba, S.; Dimitrov, D.; Valkiūnas, G.; Hagemann, A.; Tieleman, I.; Bensch, S. Haemosporidian infections in skylarks (Alauda arvensis): A comparative PCR-based and microscopy study on the parasite diversity and prevalence in southern Italy and the Netherlands. Eur. J. Wildl. Res. 2012, 58, 335–344. [Google Scholar] [CrossRef]
- Ebani, V.V.; Nardoni, S.; Giani, M.; Rocchigiani, G.; Archin, T.; Altomonte, I.; Mancianti, F. Molecular survey on the occurrence of avian haemosporidia, Coxiella burnetii and Francisella tularensis in waterfowl from central Italy. Int. J. Parasitol. Parasites Wildl. 2019, 10, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Iurescia, M.; Romiti, F.; Cocumelli, C.; Diaconu, E.L.; Stravino, F.; Onorati, R.; Alba, P.; Friedrich, K.G.; Maggi, F.; Magliano, A.; et al. Plasmodium matutinum Transmitted by Culex pipiens as a Cause of Avian Malaria in Captive African Penguins (Spheniscus demersus) in Italy. Front. Vet. Sci. 2021, 8, 621974. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Ilgunas, M.; Bukauskaitė, D.; Palinauskas, V.; Bernotienė, R.; Iezhova, T.A. Molecular characterization and distribution of Plasmodium matutinum, a common avian malaria parasite. Parasitology 2017, 144, 1726–1735. [Google Scholar] [CrossRef] [Green Version]
- García-Del-Río, M.; Sancho, R.; Martínez, J.; Merino, S. Blood parasite infections in strigiformes and psittaciformes species in captivity with a new record of potential fatal blood parasite transmission to parrots. J. Zoo. Wildl. Med. 2021, 51, 799–813. [Google Scholar] [CrossRef]
- Bennett, G.F.; Earlé, R.A.; Du Toit, H.; Huchzermeyer, F.W. A host-parasite catalogue of the haematozoa of the sub-Saharan birds. Onderstepoort J. Vet. Res. 1992, 59, 1–73. [Google Scholar] [PubMed]
- Tarello, W. Fatal Haemoproteus psittaci infection in an African grey parrot. Vet. Rec. 2005, 157, 32. [Google Scholar] [CrossRef] [PubMed]
- Belo, N.O.; Passos, L.F.; Júnior, L.M.; Goulart, C.E.; Sherlock, T.M.; Braga, E.M. Avian malaria in captive psittacine birds: Detection by microscopy and 18S rRNA gene amplification. Prev. Vet. Med. 2009, 88, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Baron, H.R.; Howe, L.; Varsani, A.; Doneley, R.J. Disease screening of three breeding populations of adult exhibition budgerigars (Melopsittacus undulatus) in New Zealand reveals a high prevalence of a novel polyomavirus and avian malaria infection. Avian. Dis. 2014, 58, 111–117. [Google Scholar] [CrossRef]
- Chagas, C.R.; Valkiūnas, G.; de Oliveira Guimarães, L.; Monteiro, E.F.; Guida, F.J.; Simões, R.F.; Rodrigues, P.T.; de Albuquerque Luna, E.J.; Kirchgatter, K. Diversity and distribution of avian malaria and related haemosporidian parasites in captive birds from a Brazilian megalopolis. Malar. J. 2017, 16, 83. [Google Scholar] [CrossRef]
- Galosi, L.; Scaglione, F.E.; Magi, G.E.; Cork, S.C.; Peirce, M.A.; Ferraro, S.; Cucuzza, L.S.; Cannizzo, F.T.; Rossi, G. Fatal Leucocytozoon Infection in a Captive Grey-headed Parrot (Poicephalus robustus suahelicus). J. Avian. Med. Surg. 2019, 33, 179–183. [Google Scholar] [CrossRef]
- Ortiz-Catedral, L.; Brunton, D.; Stidworthy, M.F.; Elsheikha, H.M.; Pennycott, T.; Schulze, C.; Braun, M.; Wink, M.; Gerlach, H.; Pendl, H.; et al. Haemoproteus minutus is highly virulent for Australasian and South American parrots. Parasit. Vectors. 2019, 12, 40. [Google Scholar] [CrossRef]
- Hellgren, O.; Waldenström, J.; Bensch, S. A new PCR assay for simultaneous studies of Leucocytozoon, Plasmodium, and Haemoproteus from avian blood. J. Parasitol. 2004, 90, 797–802. [Google Scholar] [CrossRef]
- Kent, W.J. BLAT—the BLAST–like alignment tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef] [Green Version]
- Bensch, S.; Hellgren, O.; Perez-Tris, J. MalAvi: A public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol. Ecol. Res. 2009, 9, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Ilgūnas, M.; Bukauskaitė, D.; Palinauskas, V.; Iezhova, T.A.; Dinhopl, N.; Nedorost, N.; Weissenbacher-Lang, C.; Weissenböck, H.; Valkiūnas, G. Mortality and pathology in birds due to Plasmodium (Giovannolaia) homocircumflexum infection, with emphasis on the exoerythrocytic development of avian malaria parasites. Malar. J. 2016, 15, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maqsood, M. Haemoproteus handai (sp. nov.) occurring in Indian parakeet (Psittacula cyanocephala). Ind. Vet. J. 1943, 20, 109–111. [Google Scholar]
- Sijbranda, D.; Hunter, S.; Howe, L.; Lenting, B.; Argilla, L.; Gartrell, B.D. Cases of mortality in little penguins (Eudyptula minor) in New Zealand associated with avian malaria. N. Z. Vet. J. 2017, 65, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, A.; Chavatte, J.M.; Landau, I.; Snounou, G.; Petit, T. Morphologic and molecular study of hemoparasites in wild corvids and evidence of sequence identity with Plasmodium DNA detected in captive black-footed penguins (Spheniscus demersus). J. Zoo. Wildl. Med. 2014, 45, 577–588. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cocumelli, C.; Iurescia, M.; Diaconu, E.L.; Galietta, V.; Raso, C.; Buccella, C.; Stravino, F.; Grande, F.; Fiorucci, L.; De Liberato, C.; et al. Plasmodium matutinum Causing Avian Malaria in Lovebirds (Agapornis roseicollis) Hosted in an Italian Zoo. Microorganisms 2021, 9, 1356. https://doi.org/10.3390/microorganisms9071356
Cocumelli C, Iurescia M, Diaconu EL, Galietta V, Raso C, Buccella C, Stravino F, Grande F, Fiorucci L, De Liberato C, et al. Plasmodium matutinum Causing Avian Malaria in Lovebirds (Agapornis roseicollis) Hosted in an Italian Zoo. Microorganisms. 2021; 9(7):1356. https://doi.org/10.3390/microorganisms9071356
Chicago/Turabian StyleCocumelli, Cristiano, Manuela Iurescia, Elena Lavinia Diaconu, Valentina Galietta, Caterina Raso, Carmela Buccella, Fiorentino Stravino, Francesco Grande, Letizia Fiorucci, Claudio De Liberato, and et al. 2021. "Plasmodium matutinum Causing Avian Malaria in Lovebirds (Agapornis roseicollis) Hosted in an Italian Zoo" Microorganisms 9, no. 7: 1356. https://doi.org/10.3390/microorganisms9071356
APA StyleCocumelli, C., Iurescia, M., Diaconu, E. L., Galietta, V., Raso, C., Buccella, C., Stravino, F., Grande, F., Fiorucci, L., De Liberato, C., Caprioli, A., & Battisti, A. (2021). Plasmodium matutinum Causing Avian Malaria in Lovebirds (Agapornis roseicollis) Hosted in an Italian Zoo. Microorganisms, 9(7), 1356. https://doi.org/10.3390/microorganisms9071356