
Methanogenic Biodegradation of iso-Alkanes by Indigenous Microbes from Two Different Oil Sands Tailings Ponds



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Materials
2.2. Preparation of Microcosms
2.3. Chemical Analyses
2.4. Stoichiometry of iso-Alkane Mineralization under Methanogenic Conditions
2.5. Nucleic Acid Extraction and Purification
2.6. PCR Amplification and Bioinformatics
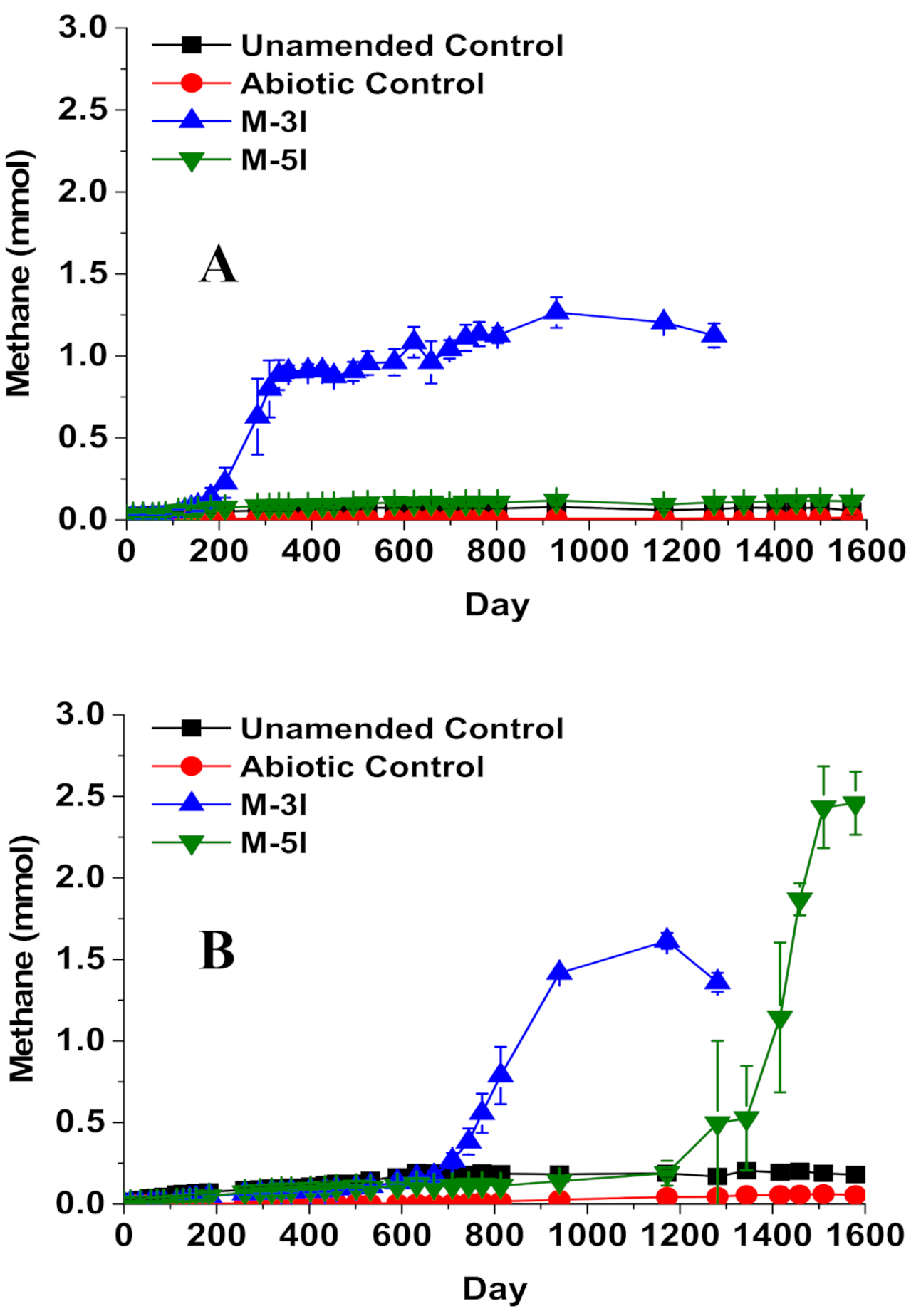
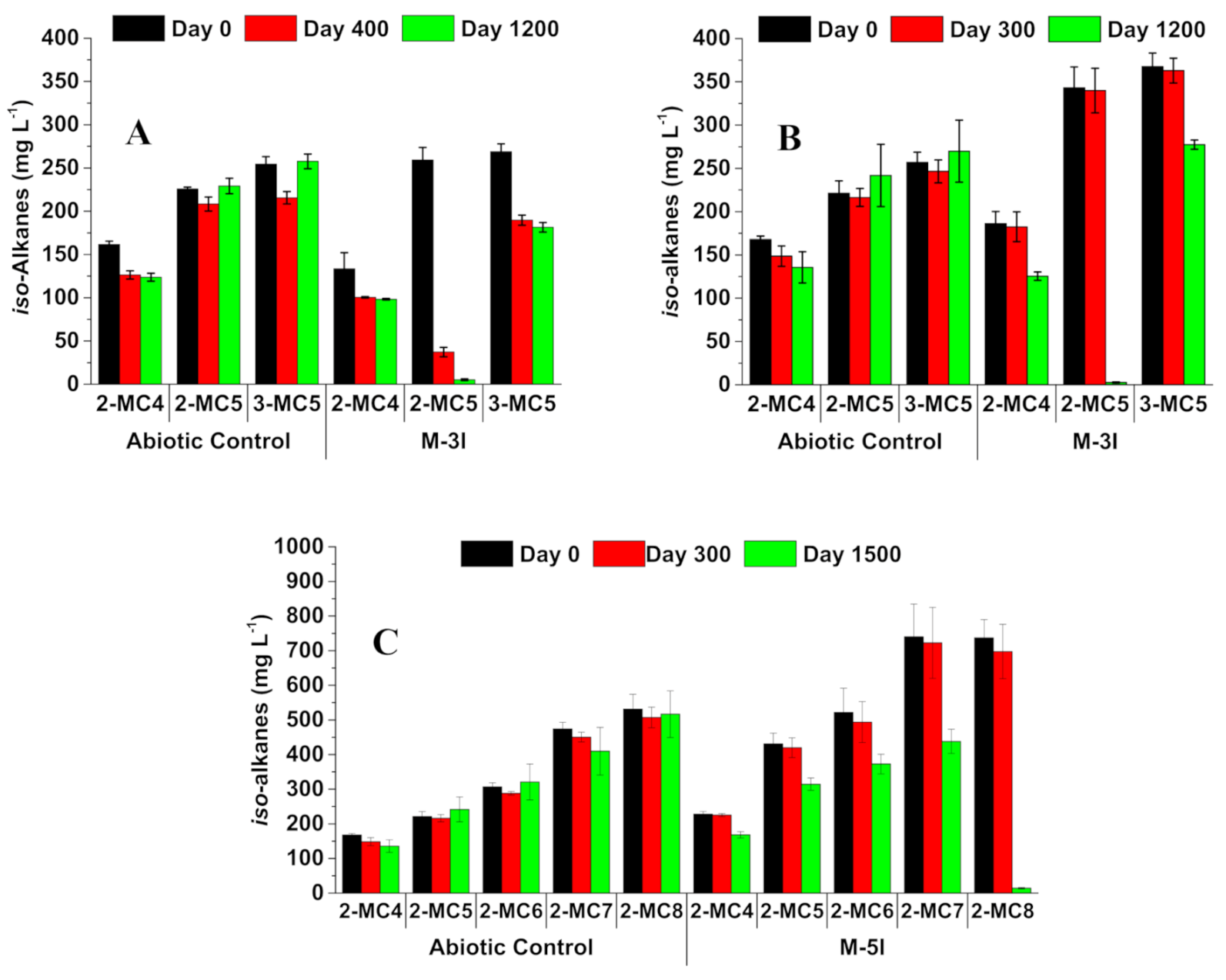
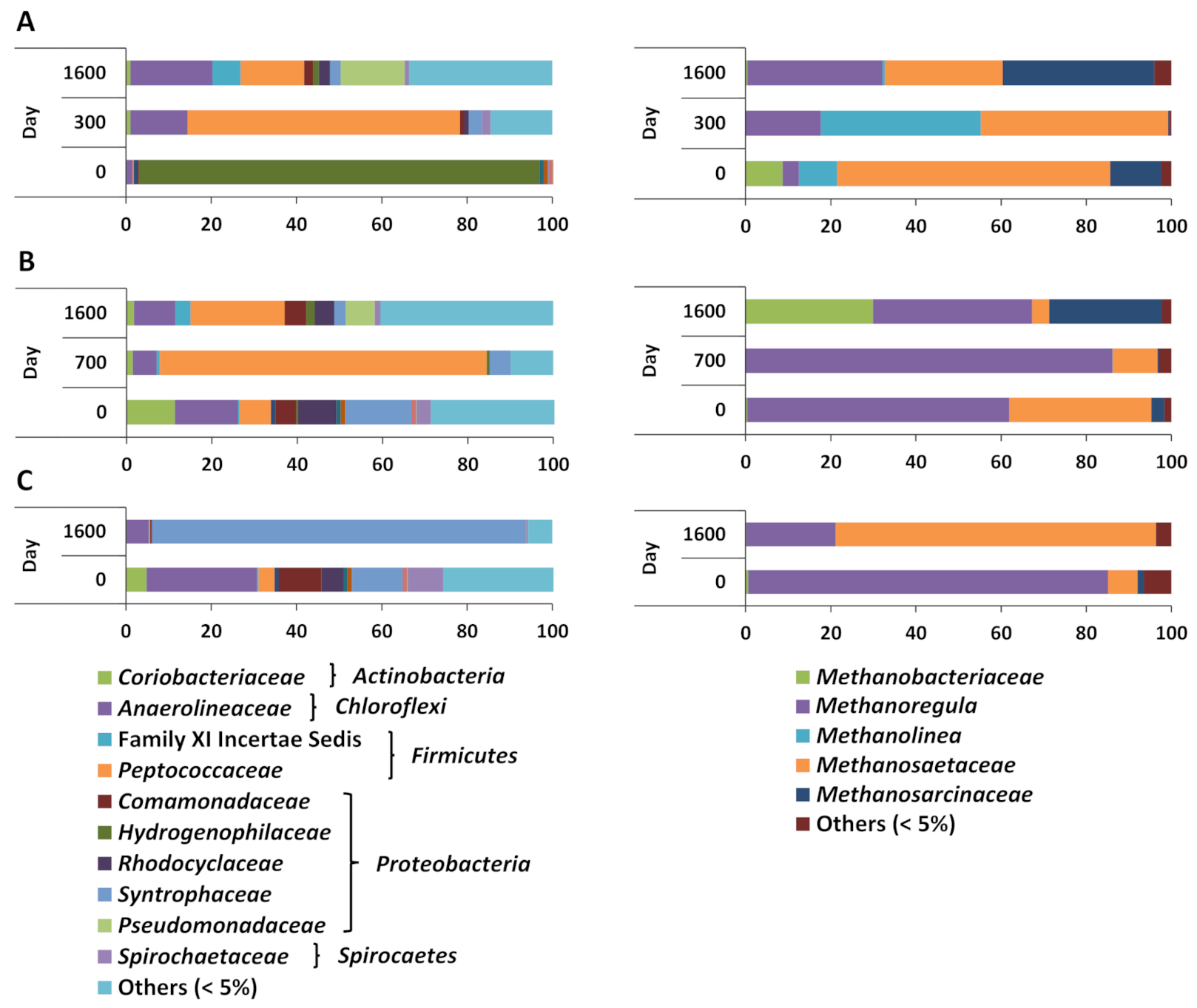
3. Results
3.1. Methanogenic Biodegradation of iso-Alkanes
3.2. Microbial Communities Involved in Biodegradation of iso-Alkane Mixtures
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rabus, R.; Hansen, T.A.; Widdel, F. Dissimilatory sulfate- and sulfur-reducing prokaryotes. In The Prokaryotes-Prokaryotic Physiology and Biochemistry; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 310–404. ISBN 978-3-642-30140-7. [Google Scholar]
- Sierra-garcia, I.N.; Oliveira, V.M. De Microbial hydrocarbon degradation: Efforts to understand biodegradation in petroleum reservoirs. In Biodegradation—Engineering and Technology; INTECH: London, UK, 2013; pp. 47–72. [Google Scholar]
- Anderson, R.T.; Lovley, D.R. Hexadecane decay by methanogenesis. Nature 2000, 404, 722–723. [Google Scholar] [CrossRef] [PubMed]
- So, C.M.; Young, L.Y. Anaerobic biodegradation of alkanes by enriched consortia under four different reducing conditions. Environ. Toxicol. Chem. 2001, 20, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.M.; Head, I.M.; Gray, N.D.; Adams, J.J.; Rowan, A.K.; Aitken, C.M.; Bennett, B.; Huang, H.; Brown, A.; Bowler, B.F.J.; et al. Crude-oil biodegradation via methanogenesis in subsurface petroleum reservoirs. Nature 2008, 451, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Callaghan, A.V. Enzymes involved in the anaerobic oxidation of n-alkanes: From methane to long-chain paraffins. Front. Microbiol. 2013, 4, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zengler, K.; Richnow, H.H.; Rosselló-Mora, R.; Michaelis, W.; Widdel, F. Methane formation from long-chain alkanes by anaerobic microorganisms. Nature 1999, 401, 266–269. [Google Scholar] [CrossRef]
- Siddique, T.; Mohamad Shahimin, M.F.M.F.; Zamir, S.; Semple, K.; Li, C.; Foght, J.M.J.M. Long-term incubation reveals methanogenic biodegradation of C5 and C6 iso-alkanes in oil sands tailings. Environ. Sci. Technol. 2015, 49, 14732–14739. [Google Scholar] [CrossRef] [PubMed]
- Siddique, T.; Semple, K.; Li, C.; Foght, J.M. Methanogenic biodegradation of iso-alkanes and cycloalkanes during long-term incubation with oil sands tailings. Environ. Pollut. 2020, 258. [Google Scholar] [CrossRef]
- Abu Laban, N.; Dao, A.; Semple, K.; Foght, J.M. Biodegradation of C7 and C8 iso-alkanes under methanogenic conditions. Environ. Microbiol. 2015, 17, 4898–4915. [Google Scholar] [CrossRef]
- Mohamad Shahimin, M.F.; Siddique, T. Methanogenic biodegradation of paraffinic solvent hydrocarbons in two different oil sands tailings. Sci. Total Environ. 2017, 583, 115–122. [Google Scholar] [CrossRef]
- Rocha, C.A.; Pedregosa, A.M.; Laborda, F. Biosurfactant-mediated biodegradation of straight and methyl-branched alkanes by pseudomonas aeruginosa ATCC 55925. AMB Express 2011, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hara, A.; Syutsubo, K.; Harayama, S. Alcanivorax which prevails in oil-contaminated seawater exhibits broad substrate specificity for alkane degradation. Environ. Microbiol. 2003, 5, 746–753. [Google Scholar] [CrossRef] [PubMed]
- Gjini, L.A. Biodegradation of Cycloalkanes under Different Redox Conditions; University of Alberta: Edmonton, AB, Canada, 2019. [Google Scholar]
- Kong, J.D.; Wang, H.; Siddique, T.; Foght, J.; Semple, K.; Burkus, Z.; Lewis, M.A. Second-generation stoichiometric mathematical model to predict methane emissions from oil sands tailings. Sci. Total Environ. 2019, 694, 133645. [Google Scholar] [CrossRef] [Green Version]
- Foght, J.M.; Gieg, L.M.; Siddique, T. The microbiology of oil sands tailings: Past, present, future. FEMS Microbiol. Ecol. 2017, 93, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Burkus, Z.; Wheler, J.; Pletcher, S. GHG Emissions from Oil Sands Tailings Ponds: Overview and Modelling Based on Fermentable Substrates; Alberta Environment and Sustainable Resource Development: Edmonton, AB, Canada, 2014; Available online: https://era.library.ualberta.ca/items/411947ce-1a33-42d5-a2cf-172fba9a2553 (accessed on 18 July 2021).
- An, D.; Caffrey, S.M.; Soh, J.; Agrawal, A.; Brown, D.; Budwill, K.; Dong, X.; Dunfield, P.F.; Foght, J.; Gieg, L.M.; et al. Metagenomics of hydrocarbon resource environments indicates aerobic taxa and genes to be unexpectedly common. Environ. Sci. Technol. 2013, 47, 10708–10717. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.L.L.; Li, C.; Ramos-Padrón, E.; Nesbø, C.; Soh, J.; Sensen, C.W.W.; Voordouw, G.; Foght, J.; Gieg, L.M.M. Oil sands tailings ponds harbour a small core prokaryotic microbiome and diverse accessory communities. J. Biotechnol. 2016, 235, 187–196. [Google Scholar] [CrossRef]
- Siddique, T.; Fedorak, P.M.; Foght, J.M. Biodegradation of short-chain n-alkanes in oil sands tailings under methanogenic conditions. Environ. Sci. Technol. 2006, 40, 5459–5464. [Google Scholar] [CrossRef]
- Mohamad Shahimin, M.F.; Foght, J.M.; Siddique, T. Preferential methanogenic biodegradation of short-chain n-alkanes by microbial communities from two different oil sands tailings ponds. Sci. Total Environ. 2016, 553, 250–257. [Google Scholar] [CrossRef]
- Siddique, T.; Penner, T.; Klassen, J.; Nesbø, C.; Foght, J.M. Microbial communities involved in methane production from hydrocarbons in oil sands tailings. Environ. Sci. Technol. 2012, 46, 9802–9810. [Google Scholar] [CrossRef]
- OSRIN. Glossary of Terms and Acronyms Used in oil Sands Mining, Processing and Environmental Management—December 2014 Update; OSRIN Rep. No. SR-1; The Oil Sands Research and Information Network: Edmonton, AB, Canada, 2010; 125p, Available online: https://era.library.ualberta.ca/items/8d620167-921d-4955-ac6c-b9d06d5b6325 (accessed on 18 July 2021).
- Siddique, T.; Fedorak, P.M.; MacKinnon, M.D.; Foght, J.M. Metabolism of BTEX and naphtha compounds to methane in oil sands tailings. Environ. Sci. Technol. 2007, 41, 2350–2356. [Google Scholar] [CrossRef] [PubMed]
- Mohamad Shahimin, M.F.; Siddique, T. Sequential biodegradation of complex naphtha hydrocarbons under methanogenic conditions in two different oil sands tailings. Environ. Pollut. 2017, 221, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Siddique, T.; Kuznetsov, P.; Kuznetsova, A.; Arkell, N.; Young, R.; Li, C.; Guigard, S.; Underwood, E.; Foght, J.M. Microbially-accelerated consolidation of oil sands tailings. Pathway I: Changes in porewater chemistry. Front. Microbiol. 2014, 5, 106. [Google Scholar] [CrossRef] [Green Version]
- Siddique, T.; Kuznetsov, P.; Kuznetsova, A.; Li, C.; Young, R.; Arocena, J.M.; Foght, J.M. Microbially-accelerated consolidation of oil sands tailings. Pathway II: Solid phase biogeochemistry. Front. Microbiol. 2014, 5, 107. [Google Scholar] [CrossRef] [PubMed]
- Siddique, T.; Gupta, R.; Fedorak, P.M.; MacKinnon, M.D.; Foght, J.M. A first approximation kinetic model to predict methane generation from an oil sands tailings settling basin. Chemosphere 2008, 72, 1573–1580. [Google Scholar] [CrossRef] [PubMed]
- Fedorak, P.M.; Hrudey, S.E. The effects of phenol and some alkyl phenolics on batch anaerobic methanogenesis. Water Res. 1984, 18, 361–367. [Google Scholar] [CrossRef]
- Holowenko, F.M.; MacKinnon, M.D.; Fedorak, P.M. Methanogens and sulfate-reducing bacteria in oil sands fine tailings waste. Can. J. Microbiol. 2000, 46, 927–937. [Google Scholar] [CrossRef]
- Symons, G.E.; Buswell, A.M. The methane fermentation of carbohydrates. J. Am. Chem. Soc. 1933, 55, 2028–2036. [Google Scholar] [CrossRef]
- Golby, S.; Ceri, H.; Marques, L.L.R.; Turner, R.J. Mixed-species biofilms cultured from an oil sand tailings pond can biomineralize metals. Microb. Ecol. 2013, 68, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Berdugo-Clavijo, C.; Dong, X.; Soh, J.; Sensen, C.W.; Gieg, L.M. Methanogenic biodegradation of two-ringed polycyclic aromatic hydrocarbons. FEMS Microbiol. Ecol. 2012, 81, 124–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowler, S.J.; Dong, X.; Sensen, C.W.; Suflita, J.M.; Gieg, L.M. Methanogenic toluene metabolism: Community structure and intermediates. Environ. Microbiol. 2012, 14, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Soh, J.; Dong, X.; Caffrey, S.M.; Voordouw, G.; Sensen, C.W. Phoenix 2: A locally installable large-scale 16S rRNA gene sequence analysis pipeline with Web interface. J. Biotechnol. 2013, 167, 393–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fedorak, P.M.; Coy, D.L.; Dudas, M.J.; Simpson, M.J.; Renneberg, J.; Mackinnon, M.D. Microbially-mediated fugitive gas production from oil sands tailings and increased tailings densification rates. J. Environ. Eng. Sci. 2003, 2, 199–211. [Google Scholar] [CrossRef]
- Arkell, N.; Kuznetsov, P.; Kuznetsova, A.; Foght, J.M.; Siddique, T. Microbial metabolism alters pore water chemistry and increases consolidation of oil sands tailings. J. Environ. Qual. 2015, 44, 145–153. [Google Scholar] [CrossRef]
- Zamanpour, M.K.; Kaliappan, R.S.; Rockne, K.J. Gas ebullition from petroleum hydrocarbons in aquatic sediments: A review. J. Environ. Manag. 2020, 271, 110997. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Semple, K.; Foght, J.M. Anaerobic alkane biodegradation by cultures enriched from oil sands tailings ponds involves multiple species capable of fumarate addition. FEMS Microbiol. Ecol. 2015, 91, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heipieper, H.J.; Martinez, P.M. Toxicity of hydrocarbons to microorganisms. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin, Germany, 2010; pp. 1566–1573. [Google Scholar]
- Chen, J.; Liu, Y.; Zhou, L.; Mbadinga, S.M.; Yang, T.; Zhou, J.; Liu, J. Methanogenic degradation of branched alkanes in enrichment cultures of production water from a high-temperature petroleum reservoir. Appl. Microbiol. Biotechnol. 2019, 103, 2391–2401. [Google Scholar] [CrossRef] [PubMed]
- Siddique, T.; Penner, T.; Semple, K.; Foght, J.M. Anaerobic biodegradation of longer-chain n-alkanes coupled to methane production in oil sands tailings. Environ. Sci. Technol. 2011, 45, 5892–5899. [Google Scholar] [CrossRef] [PubMed]
- Straub, K.L.; Benz, M.; Schink, B.; Widdel, F. Anaerobic, nitrate-dependent microbial oxidation of ferrous iron. Appl. Environ. Microbiol. 1996, 62, 1458–1460. [Google Scholar] [CrossRef] [Green Version]
- Bosch, J.; Lee, K.Y.; Jordan, G.; Kim, K.W.; Meckenstock, R.U. Anaerobic, nitrate-dependent oxidation of pyrite nanoparticles by thiobacillus denitrificans. Environ. Sci. Technol. 2012, 46, 2095–2101. [Google Scholar] [CrossRef]
- Tan, B.; Charchuk, R.; Li, C.; Laban, N.A.; Foght, J.M. Draft genome sequence of uncultivated firmicutes (Peptococcaceae SCADC) single cells sorted from methanogenic alkane-degrading cultures. Genome Announc. 2014, 2, e00909–e00914. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.; Jane Fowler, S.; Laban, N.A.; Dong, X.; Sensen, C.W.; Foght, J.; Gieg, L.M. Comparative analysis of metagenomes from three methanogenic hydrocarbon-degrading enrichment cultures with 41 environmental samples. ISME J. 2015, 9, 2028–2045. [Google Scholar] [CrossRef] [Green Version]
- Tan, B.; Nesbø, C.; Foght, J.M. Re-analysis of omics data indicates Smithella may degrade alkanes by addition to fumarate under methanogenic conditions. ISME J. 2014, 8, 2353–2356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, X.-Y.; Maurice Mbadinga, S.; Liu, Y.-F.; Yang, S.-Z.; Liu, J.-F.; Ye, R.-Q.; Gu, J.-D.; Mu, B.-Z. Insights into the anaerobic biodegradation pathway of n-alkanes in oil reservoirs by detection of signature metabolites. Sci. Rep. 2015, 5, 9801. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Wang, L.-Y.; Mbadinga, S.M.; Liu, J.-F.; Yang, S.-Z.; Gu, J.-D.; Mu, B.-Z. Anaerolineaceae and Methanosaeta turned to be the dominant microorganisms in alkanes-dependent methanogenic culture after long-term of incubation. AMB Express 2015, 5, 37. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Substrate | Incubation Time (d) | Substrate Added at Day 0 (mmol) | Substrate Consumed (mmol) ǂ | Theoretical Methane Yield * (mmol) | Total Predicted Methane Yield * (mmol) | Total Measured Methane Yield (mmol) | Percent of Theoretical Production (%) | |
|---|---|---|---|---|---|---|---|---|
| CNUL M-3I | 2-MC4 | 392 | 0.26 (±0.02) | 0.11 (±0.02) | 0.46 (±0.09) | 1.99 (±0.15) | 0.91 (±0.04) | 45.7 |
| 2-MC5 | 0.30 (±0.20) | 0.22 (±0.01) | 1.05 (±0.05) | |||||
| 3-MC5 | 0.31 (±0.01) | 0.10 (±0.00) | 0.48 (±0.02) | |||||
| CNRL M-3I | 2-MC4 | 1172 | 0.26 (±0.02) | 0.08 (±0.01) | 0.34 (±0.04) | 2.93 (±0.23) | 1.61 (±0.05) | 55.0 |
| 2-MC5 | 0.40 (±0.03) | 0.39 (±0.03) | 1.85 (±0.13) | |||||
| 3-MC5 | 0.43 (±0.02) | 0.16 (±0.01) | 0.75 (±0.06) | |||||
| CNRL M-5I | 2-MC4 | 1459 | 0.32 (±0.01) | 0.09 (±0.00) | 0.30 (±0.00) | 7.57 (±0.86) | 2.43 (±0.25) | 32.2 |
| 2-MC5 | 0.50 (±0.03) | 0.20 (±0.02) | 0.79 (±0.09) | |||||
| 2-MC6 | 0.53 (±0.05) | 0.21 (±0.04) | 0.94 (±0.24) | |||||
| 2-MC7 | 0.64 (±0.06) | 0.26 (±0.05) | 1.45 (±0.28) | |||||
| 2-MC8 | 0.57 (±0.03) | 0.50 (±0.03) | 3.46 (±0.24) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamad Shahimin, M.F.; Foght, J.M.; Siddique, T. Methanogenic Biodegradation of iso-Alkanes by Indigenous Microbes from Two Different Oil Sands Tailings Ponds. Microorganisms 2021, 9, 1569. https://doi.org/10.3390/microorganisms9081569
Mohamad Shahimin MF, Foght JM, Siddique T. Methanogenic Biodegradation of iso-Alkanes by Indigenous Microbes from Two Different Oil Sands Tailings Ponds. Microorganisms. 2021; 9(8):1569. https://doi.org/10.3390/microorganisms9081569
Chicago/Turabian StyleMohamad Shahimin, Mohd Faidz, Julia M. Foght, and Tariq Siddique. 2021. "Methanogenic Biodegradation of iso-Alkanes by Indigenous Microbes from Two Different Oil Sands Tailings Ponds" Microorganisms 9, no. 8: 1569. https://doi.org/10.3390/microorganisms9081569
APA StyleMohamad Shahimin, M. F., Foght, J. M., & Siddique, T. (2021). Methanogenic Biodegradation of iso-Alkanes by Indigenous Microbes from Two Different Oil Sands Tailings Ponds. Microorganisms, 9(8), 1569. https://doi.org/10.3390/microorganisms9081569