
Biodiversity of Actinomycetes from Heavy Metal Contaminated Technosols


Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Characterization of Isolates
2.2. Molecular Identification and Diversity Estimation
2.3. Analysis of Heavy Metal Resistance
3. Results
3.1. Isolation and Characterization of Isolates
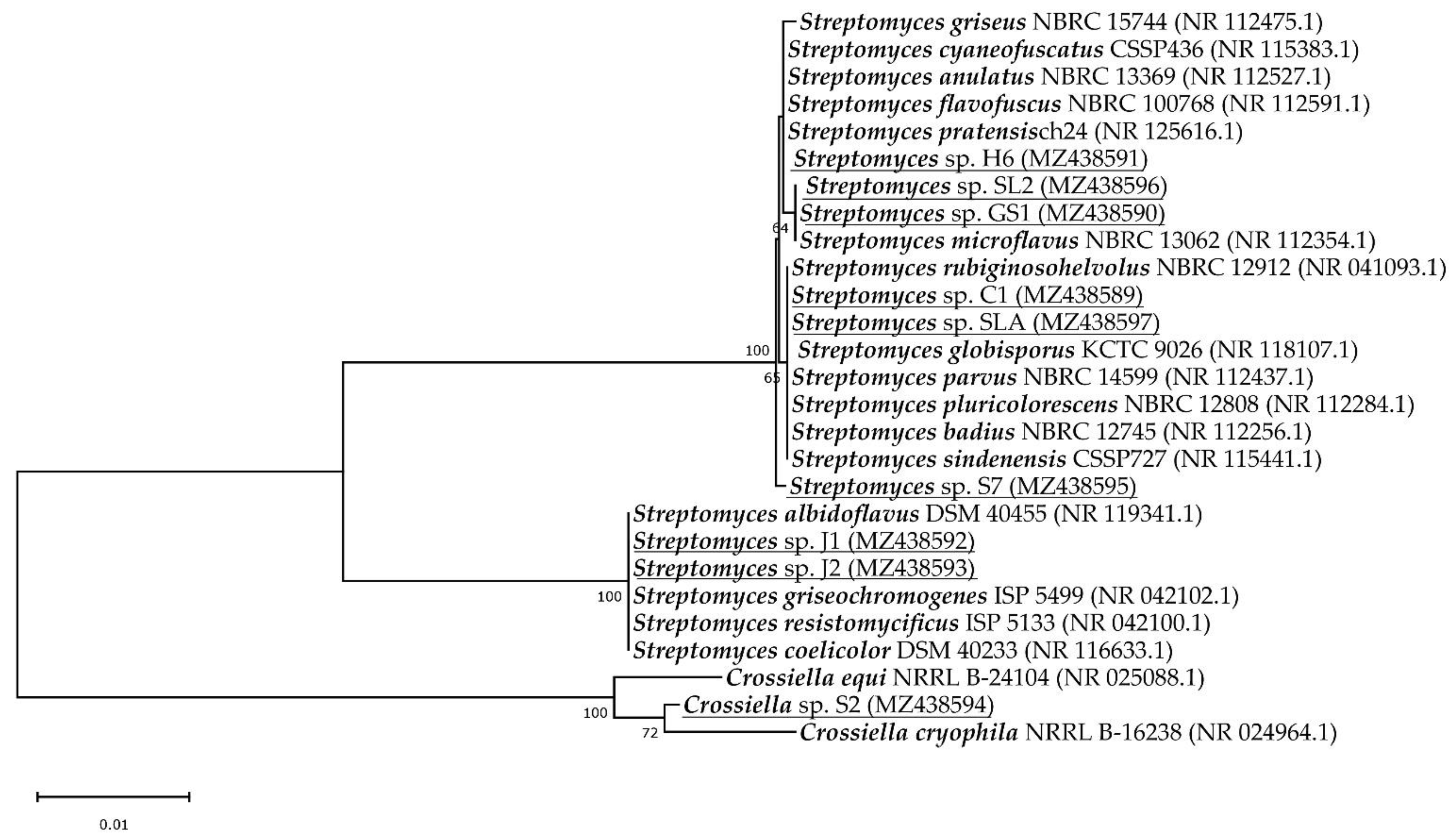
3.2. Identification of Isolates
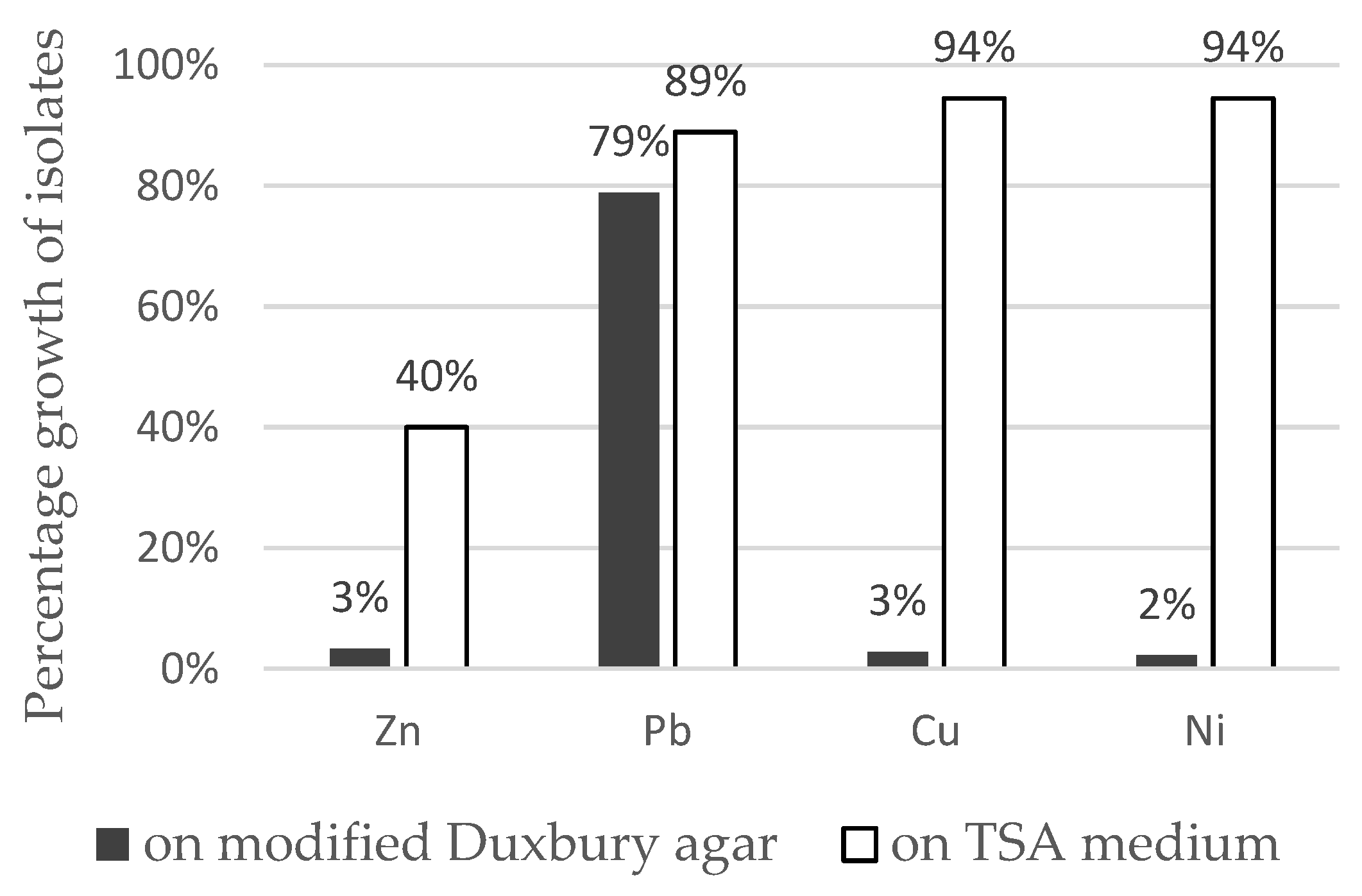
3.3. Heavy Metal Resistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Uzarowicz, Ł.; Charzyński, P.; Greinert, A.; Hulisz, P.; Kabała, C.; Kusza, G.; Kwasowski, W.; Pędziwiatr, A. Studies of Technogenic Soils in Poland: Past, Present, and Future Perspectives. Soil Sci. Ann. 2020, 71, 281–299. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy Metal Toxicity and the Environment. Exp. Suppl. 2012, 101, 133–164. [Google Scholar] [CrossRef] [Green Version]
- Ahluwalia, S.S.; Goyal, D. Microbial and Plant Derived Biomass for Removal of Heavy Metals from Wastewater. Bioresour. Technol. 2007, 98, 2243–2257. [Google Scholar] [CrossRef] [PubMed]
- Giller, K.E.; Witter, E.; Mcgrath, S.P. Toxicity of Heavy Metals to Microorganisms and Microbial Processes in Agricultural Soils: A Review. Soil Biol. Biochem. 1998, 30, 1389–1414. [Google Scholar] [CrossRef]
- Torsvik, V.; Øvreås, L. Chapter 2 Microbial Diversity, Life Strategies, and Adaptation to Life in Extreme Soils; Dion, P., Nautiyal, C.S., Eds.; Springer: Berlin/Heidelberg, Germany, 2008; Volume 13. [Google Scholar]
- Hur, M.; Park, S.-J. Identification of Microbial Profiles in Heavy-Metal-Contaminated Soil from Full-Length 16S RRNA Reads Sequenced by a PacBio System. Microorganisms 2019, 7, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez, A.; Catalano, S.A.; Amoroso, M.J. Heavy Metal Resistant Strains Are Widespread along Streptomyces Phylogeny. Mol. Phylogenet. 2013, 66, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Frankovská, J.; Slaninka, I.; Kordík, J.; Jurkovič, Ľ.; Greif, V.; Šottník, P.; Dananaj, I.; Mikita, S.; Dercová, K.; Jánová, V. Atlas Sanačných Metód Environmentálnych Záťaží; Štátny Geologický Ústav Dionýza Štúra: Bratislava, Slovakia, 2010. [Google Scholar]
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic Names with Standing in Nomenclature (LPSN) Moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef]
- Kämpfer, P.; Glaeser, S.P.; Parkes, L.; van Keulen, G.; Dyson, P. The Family Streptomycetaceae. In The Prokaryotes: Actinobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 889–1010. [Google Scholar] [CrossRef]
- Law, J.W.-F.; Chan, K.-G.; He, Y.-W.; Khan, T.M.; Ab Mutalib, N.-S.; Goh, B.-H.; Lee, L.-H. Diversity of Streptomyces Spp. from Mangrove Forest of Sarawak (Malaysia) and Screening of Their Antioxidant and Cytotoxic Activities. Sci. Rep. 2019, 9, 15262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayed, A.M.; Hassan, M.H.A.; Alhadrami, H.A.; Hassan, H.M.; Goodfellow, M.; Rateb, M.E. Extreme Environments: Microbiology Leading to Specialized Metabolites. J. Appl. Microbiol. 2019, 128, 630–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slovenská Agentúra Životného Prostredia. Správa o Stave Životného Prostredia Slovenskej Republiky v Roku 2019. 2019. Available online: https://www.enviroportal.sk/spravy/detail/10661 (accessed on 10 June 2021).
- Fargašová, A. Znečistenie Kovmi na Slovensku. Štátny Geologický Ústav Dionýza Štúra, Bratislava. 2010. Available online: https://docplayer.cz/39517320-Znecistenie-kovmi-na-slovensku-2009.html (accessed on 10 June 2021).
- Kučerová, G.; Majzlan, J.; Lalinská-Voleková, B.; Radková, A.; Bačík, P.; Michňová, J.; Šottník, P.; Jurkovič, L.; Klimko, T.; Steininger, R.; et al. Mineralogy of Neutral Mine Drainage in the Tailings of Siderite-Cu Ores in Eastern Slovakia. Can. Miner. 2014, 52, 779–798. [Google Scholar] [CrossRef]
- Tóth, R.; Škultétyová, S.; Hiller, E.; Jurkovič, Ľ.; Šottník, P.; Jaroslav, V.; Tatarková, V. Mobility of Potentially Toxic Elements in Sediments of the Tailings Impoundment Slovinky and Their Transport into the Surrounding Environment; Burkhard, J., Černík, M., Halousková, O., Eds.; Vodní Zdroje Ekomonitor Spol. s r.o: Chrudim, Czech Republic, 2013. [Google Scholar]
- Takac, P.; Kozakova, L.; Valkova, M.; Zelenak, F. Heavy Metals in Soils in Middle Spiš. Acta Montan. Slovaca 2008, 13, 82–86. [Google Scholar]
- Belaňová, E. Monitoring of Qualitative State of Soils in Hnúšťa. ACTA Fac. Ecol. 2015, 33, 5–13. [Google Scholar]
- Baroniaková, K. Streptomycéty z Prostredí Kontaminovaných Ťažkými Kovmi-Genetická a Biologická Charakterizácia, Košice. 2018. Available online: https://opac.crzp.sk/?fn=detailBiblioForm&sid=BCB1C6D618A1774C85DFCD5E7FBF (accessed on 10 June 2021).
- Porter, J.N.; Wilhelm, J.J.; Tresner, H.D. Method for the Preferential Isolation of Actinomycetes from Soils. Appl. Microbiol. 1960, 8, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, M.D.; Kharel, M.K.; Bosserman, M.A.; Rohr, J. Laboratory Maintenance of Streptomyces Species. Curr. Protoc. Microbiol. 2010, 18, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tresner, H.D.; Backus, E.J. System of Color Wheels for Streptomycete Taxonomy. Appl. Microbiol. 1963, 11, 335–338. [Google Scholar] [CrossRef]
- Hasan, F.; Shah, A.A.; Hameed, A. Methods for Detection and Characterization of Lipases: A Comprehensive Review. Biotechnol. Adv. 2009, 27, 782–798. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Banu, R.; Moni, R.; Islam, N.; Ruma, M.K.; Zohora, U.S. Investigation on a Bangladeshi Isolate Bacillus Aryabhattai for Promising Biotechnological Applications. Jahangirnagar Univ. J. Biol. Sci. 2018, 7, 33–45. [Google Scholar] [CrossRef]
- Sakthivel, M.; Karthikeyan, N.; Jayaveny, R.; Palani, P. Optimization of Culture Conditions for the Production of Extracellular Cellulase from Corynebacterium Lipophiloflavum. J. Ecobiotechnol. 2010, 2, 6–13. [Google Scholar]
- Nybo, S.E.; Shepherd, M.D.; Bosserman, M.A.; Rohr, J. Genetic Manipulation of Streptomyces Species. Curr. Protoc. Microbiol. 2010, 19, 1–26. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S Ribosomal DNA Amplification for Phylogenetic Study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Vandžurová, A.; Bačkor, P.; Javorský, P.; Pristaš, P. Staphylococcus Nepalensis in the Guano of Bats (Mammalia). Vet. Microbiol. 2013, 164, 116–121. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A Greedy Algorithm for Aligning DNA Sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Rong, X.; Huang, Y. Taxonomic Evaluation of the Streptomyces Griseus Clade Using Multilocus Sequence Analysis and DNA–DNA Hybridization, with Proposal to Combine 29 Species and Three Subspecies as 11 Genomic Species. Int. J. Syst. Evol. Microbiol. 2010, 60, 696–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Baz, S.; Baz, M.; Barakate, M.; Hassani, L.; El Gharmali, A.; Imziln, B. Resistance to and Accumulation of Heavy Metals by Actinobacteria Isolated from Abandoned Mining Areas. Sci. World J. 2015, 2015, 761834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Updating the 97% Identity Threshold for 16S Ribosomal RNA OTUs. Bioinformatics 2018, 34, 2371–2375. [Google Scholar] [CrossRef]
- Mohammadipanah, F.; Hamedi, J.; Spröer, C.; Rohde, M.; del Carmen Montero-Calasanz, M.; Klenk, H.-P. Streptomyces Zagrosensis Sp. Nov., Isolated from Soil. Int. J. Syst. Evol. Microbiol. 2014, 64, 3434–3440. [Google Scholar] [CrossRef]
- Rong, X.; Guo, Y.; Huang, Y. Proposal to Reclassify the Streptomyces Albidoflavus Clade on the Basis of Multilocus Sequence Analysis and DNA–DNA Hybridization, and Taxonomic Elucidation of Streptomyces Griseus Subsp. Solvifaciens. Syst. Appl. Microbiol. 2009, 32, 314–322. [Google Scholar] [CrossRef]
- Kim, K.-O.; Shin, K.-S.; Kim, M.N.; Shin, K.-S.; Labeda, D.P.; Han, J.-H.; Kim, S.B. Reassessment of the Status of Streptomyces Setonii and Reclassification of Streptomyces Fimicarius as a Later Synonym of Streptomyces Setonii and Streptomyces Albovinaceus as a Later Synonym of Streptomyces Globisporus Based on Combined 16S RRNA/GyrB Gene Sequence Analysis. Int. J. Syst. Evol. Microbiol. 2012, 62, 2978–2985. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Shi, Y.; Zhang, Y.; Zhou, Z.; Lu, Z.; Li, W.; Huang, Y.; Rodríguez, C.; Goodfellow, M. Classification of Streptomyces Griseus (Krainsky 1914) Waksman and Henrici 1948 and Related Species and the Transfer of ‘Microstreptospora Cinerea’ to the Genus Streptomyces as Streptomyces Yanii Sp. Nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 1605–1610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komaki, H.; Tamura, T. Reclassification of Streptomyces Fulvissimus as a Later Heterotypic Synonym of Streptomyces Microflavus. Int. J. Syst. Evol. Microbiol. 2020, 70, 5156–5162. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A Simple Method for Estimating Evolutionary Rate of Base Substitutions through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Duxbury, T. Toxicity of Heavy Metals to Soil Bacteria. FEMS Microbiol. Lett. 1981, 11, 217–220. [Google Scholar] [CrossRef]
- Pristas, P.; Stramova, Z.; Kvasnova, S.; Judova, J.; Perhacova, Z.; Vidova, B.; Sramkova, Z.; Godany, A. Non-Ferrous Metal Industry Waste Disposal Sites As A Source of Poly-Extremotolerant Bacteria. In Nova Biotechnologica et Chimica; Walter de Gruyter GmbH: Berlin, Germany, 2015; Volume 14, pp. 62–68. [Google Scholar] [CrossRef]
- Michaeli, E.; Boltižiar, M.; Solár, V.; Vilček, J.; Ivanová, M. Landfill Waste from the Production of Nickel and Cobalt in the Slovak Republic. Int. J. Metall. Mater. Eng. 2016, 2, 126. [Google Scholar] [CrossRef]
- Michaeli, E.; Solár, V.; Maxin, M.; Vilček, J.; Boltižiar, M. The Nature of the Technosols on the Waste from Nickel Production. Sustainability 2021, 13, 406. [Google Scholar] [CrossRef]
- Karelová, E.; Harichová, J.; Stojnev, T.; Pangallo, D.; Ferianc, P. The Isolation of Heavy-Metal Resistant Culturable Bacteria and Resistance Determinants from a Heavy-Metal-Contaminated Site. Biologia 2011, 66, 18–26. [Google Scholar] [CrossRef]
- Sedlakova-Kadukova, J.; Kopcakova, A.; Gresakova, L.; Godany, A.; Pristas, P. Bioaccumulation and Biosorption of Zinc by a Novel Streptomyces K11 Strain Isolated from Highly Alkaline Aluminium Brown Mud Disposal Site. Ecotoxicol. Environ. Saf. 2019, 167, 204–211. [Google Scholar] [CrossRef]
- Polti, M.A.; Amoroso, M.J.; Abate, C.M. Chromium(VI) Resistance and Removal by Actinomycete Strains Isolated from Sediments. Chemosphere 2007, 67, 660–667. [Google Scholar] [CrossRef]
- Albarracín, V.H.; Amoroso, M.J.; Abate, C.M. Isolation and Characterization of Indigenous Copper-Resistant Actinomycete Strains. Geochemistry 2005, 65, 145–156. [Google Scholar] [CrossRef]
- Albarracín, V.H.; Ávila, A.L.; Amoroso, M.J.; Abate, C.M. Copper Removal Ability by Streptomyces Strains with Dissimilar Growth Patterns and Endowed with Cupric Reductase Activity. FEMS Microbiol. Lett. 2008, 288, 141–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siñeriz, M.L.; Kothe, E.; Abate, C.M. Cadmium Biosorption by Streptomyces Sp. F4 Isolated from Former Uranium Mine. J. Basic Microbiol. 2009, 49, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Rho, J.-Y.; Kim, J.-H. Heavy Metal Biosorption and Its Significance to Metal Tolerance of Streptomycetes. J. Microbiol. 2002, 40, 51–54. [Google Scholar]
- Amoroso, M.J.; Castro, G.R.; Durán, A.; Peraud, O.; Oliver, G.; Hill, R.T. Chromium Accumulation by Two Streptomyces Spp. Isolated from Riverine Sediments. J. Ind. Microbiol. Biotechnol. 2001, 26, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Ravel, J.; Amoroso, M.J.; Colwell, R.R.; Hill, R.T. Mercury-Resistant Actinomycetes from the Chesapeake Bay. FEMS Microbiol. Lett. 1998, 162, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Van Nostrand, J.D.; Khijniak, T.V.; Gentry, T.J.; Novak, M.T.; Sowder, A.G.; Zhou, J.Z.; Bertsch, P.M.; Morris, P.J. Isolation and Characterization of Four Gram-Positive Nickel-Tolerant Microorganisms from Contaminated Sediments. Microb. Ecol. 2007, 53, 670–682. [Google Scholar] [CrossRef] [PubMed]
- Majzlik, P.; Strasky, A.; Adam, V.; Nemec, M.; Trnkova, L.; Zehnalek, J.; Hubalek, J.; Provaznik, I.; Kizek, R. Influence of Zinc(II) and Copper(II) Ions on Streptomyces Bacteria Revealed by Electrochemistry. Int. J. Electrochem. Sci. 2011, 6, 2171–2191. [Google Scholar]
- Schmidt, A.; Schmidt, A.; Haferburg, G.; Kothe, E. Superoxide Dismutases of Heavy Metal Resistant Streptomycetes. J. Basic Microbiol. 2007, 47, 56–62. [Google Scholar] [CrossRef]
- Schmidt, A.; Haferburg, G.; Sineriz, M.; Merten, D.; Büchel, G.; Kothe, E. Heavy Metal Resistance Mechanisms in Actinobacteria for Survival in AMD Contaminated Soils. Geochemistry 2005, 65, 131–144. [Google Scholar] [CrossRef]
- Schmidt, A.; Haferburg, G.; Schmidt, A.; Lischke, U.; Merten, D.; Ghergel, F.; Büchel, G.; Kothe, E. Heavy Metal Resistance to the Extreme: Streptomyces Strains from a Former Uranium Mining Area. Geochemistry 2009, 69, 35–44. [Google Scholar] [CrossRef]
- Guo, J.K.; Lin, Y.B.; Zhao, M.L.; Sun, R.; Wang, T.T.; Tang, M.; Wei, G.H. Streptomyces Plumbiresistens Sp. Nov., a Lead-Resistant Actinomycete Isolated from Lead-Polluted Soil in North-West China. Int. J. Syst. Evol. Microbiol. 2009, 59, 1326–1330. [Google Scholar] [CrossRef]
- Lin, Y.B.; Wang, X.Y.; Li, H.F.; Wang, N.N.; Wang, H.X.; Tang, M.; Wei, G.-H. Streptomyces Zinciresistens Sp. Nov., a Zinc-Resistant Actinomycete Isolated from Soil from a Copper and Zinc Mine. Int. J. Syst. Evol. Microbiol. 2011, 61, 616–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desjardin, V.; Bayard, R.; Huck, N.; Manceau, A.; Gourdon, R. Effect of Microbial Activity on the Mobility of Chromium in Soils. Waste Manag. 2002, 22, 195–200. [Google Scholar] [CrossRef]
- Dávila Costa, J.S.; Hoskisson, P.A.; Paterlini, P.; Romero, C.M.; Alvarez, A. Whole Genome Sequence of the Multi-Resistant Plant Growth-Promoting Bacteria Streptomyces Sp. Z38 with Potential Application in Agroindustry and Bio-Nanotechnology. Genomics 2020, 112, 4684–4689. [Google Scholar] [CrossRef] [PubMed]
- Simón Solá, M.Z.; Lovaisa, N.; Dávila Costa, J.S.; Benimeli, C.S.; Polti, M.A.; Alvarez, A. Multi-Resistant Plant Growth-Promoting Actinobacteria and Plant Root Exudates Influence Cr(VI) and Lindane Dissipation. Chemosphere 2019, 222, 679–687. [Google Scholar] [CrossRef]
- Undabarrena, A.; Ugalde, J.A.; Seeger, M.; Cámara, B. Genomic Data Mining of the Marine Actinobacteria Streptomyces Sp. H-KF8 Unveils Insights into Multi-Stress Related Genes and Metabolic Pathways Involved in Antimicrobial Synthesis. Peer J. 2017, 5, 1–35. [Google Scholar] [CrossRef] [Green Version]
- Undabarrena, A.; Beltrametti, F.; Claverías, F.P.; González, M.; Moore, E.R.B.; Seeger, M.; Cámara, B. Exploring the Diversity and Antimicrobial Potential of Marine Actinobacteria from the Comau Fjord in Northern Patagonia, Chile. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ding, C.; Liao, J.; Du, L.; Sun, Q.; Yang, J.; Yang, Y.; Zhang, D.; Tang, J.; Liu, N. Bioaccumulation Characterization of Uranium by a Novel Streptomyces Sporoverrucosus Dwc-3. J. Environ. Sci. 2016, 41, 162–171. [Google Scholar] [CrossRef]
- Tang, X.; Zhao, J.; Li, K.; Chen, Z.; Sun, Y.; Gao, J. Streptomyces Cyaneochromogenes Sp. Nov., a Blue Pigment-Producing Actinomycete from Manganese-Contaminated Soil. Int. J. Syst. Evol. Microbiol. 2019, 69, 2202–2207. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Yang, H.; Zhou, H.; Wang, C.; Fu, F.; Yu, Y.; Lu, X.; Tian, Y. Complete Genome Sequence of the Streptomyces Sp. Strain CdTB01, a Bacterium Tolerant to Cadmium. J. Biotechnol. 2016, 229, 42–43. [Google Scholar] [CrossRef] [PubMed]
- Mo, P.; Zhao, J.; Li, K.; Tang, X.; Gao, J. Streptomyces Manganisoli Sp. Nov., a Novel Actinomycete Isolated from Manganese-Contaminated Soil. Int. J. Syst. Evol. Microbiol. 2018, 68, 1890–1895. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Tang, X.; Zhao, J.; Guo, Y.; Tang, Y.; Gao, J. Streptomyces Cadmiisoli Sp. Nov., a Novel Actinomycete Isolated from Cadmium-Contaminated Soil. Int. J. Syst. Evol. Microbiol. 2019, 69, 1024–1029. [Google Scholar] [CrossRef]
- Mo, P.; Yu, Y.-Z.; Zhao, J.-R.; Gao, J. Streptomyces Xiangtanensis Sp. Nov., Isolated from a Manganese-Contaminated Soil. Antonie Leeuwenhoek 2017, 110, 297–304. [Google Scholar] [CrossRef]
- Li, H.; Lin, Y.; Guan, W.; Chang, J.; Xu, L.; Guo, J.; Wei, G. Biosorption of Zn(II) by Live and Dead Cells of Streptomyces Ciscaucasicus Strain CCNWHX 72-14. J. Hazard. Mater. 2010, 179, 151–159. [Google Scholar] [CrossRef]
- Lin, Y.; Wang, X.; Wang, B.; Mohamad, O.; Wei, G. Bioaccumulation Characterization of Zinc and Cadmium by Streptomyces Zinciresistens, a Novel Actinomycete. Ecotoxicol. Environ. Saf. 2012, 77, 7–17. [Google Scholar] [CrossRef]
- Sanjenbam, P.; Saurav, K.; Kannabiran, K. Biosorption of Mercury and Lead by Aqueous Streptomyces VITSVK9 Sp. Isolated from Marine Sediments from the Bay of Bengal, India. Front. Chem. Sci. Eng. 2012, 6, 198–202. [Google Scholar] [CrossRef]
- Qin, S.; Li, W.-J.; Dastager, S.G.; Hozzein, W.N. Editorial: Actinobacteria in Special and Extreme Habitats: Diversity, Function Roles, and Environmental Adaptations. Front. Microbiol. 2016, 7, 6–7. [Google Scholar] [CrossRef] [Green Version]
- Labeda, D.P.; Goodfellow, M.; Brown, R.; Ward, A.C.; Lanoot, B.; Vanncanneyt, M.; Swings, J.; Kim, S.-B.; Liu, Z.; Chun, J.; et al. Phylogenetic Study of the Species within the Family Streptomycetaceae. Antonie Leeuwenhoek 2012, 101, 73–104. [Google Scholar] [CrossRef]
- Antony-Babu, S.; Stien, D.; Eparvier, V.; Parrot, D.; Tomasi, S.; Suzuki, M.T. Multiple Streptomyces Species with Distinct Secondary Metabolomes Have Identical 16S RRNA Gene Sequences. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Labeda, D.P. Crossiella Gen. Nov., a New Genus Related to Streptoalloteichus. Int. J. Syst. Evol. Microbiol. 2001, 51, 1575–1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Sampling Site | Isolate | Substrate Mycelium | Aerial Mycelium | Pigment | Protease | Lipase | Amylase | Cellulase | 37 °C | pH 5 | pH 7 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Gelnica | C1 | O | W | - | + | ± | + | ± | + | + | + |
| GS1 | B | W | - | + | - | + | ± | - | + | + | |
| H6 | B | W | Br | + | + | + | ± | - | + | + | |
| J1 | B | W | - | + | + | + | - | + | + | + | |
| J2 | B | W | - | + | + | + | - | + | + | + | |
| Hnúšťa | S2 | B | W | - | - | ± | - | + | - | + | + |
| S7 | O | C | Br | - | - | + | - | - | + | + | |
| Slovinky | SL2 | B | W | - | + | ± | + | ± | + | + | + |
| SLA | Y-Br | W | - | + | ± | + | ± | + | + | + |
| Sampling Site | Isolate | MIC [mg/L] | |||
|---|---|---|---|---|---|
| Cu | Ni | Pb | Zn | ||
| Gelnica | C1 | 4 | 8 | 300 | 250 |
| GS1 | 300 | 300 | 1000 | 300 | |
| H6 | 300 | 8 | 1000 | 300 | |
| J1 | 2 | 100 | 300 | 100 | |
| J2 | 2 | 8 | 300 | 8 | |
| Hnúšťa | S2 | 64 | 100 | 250 | 100 |
| S7 | 300 | 150 | 125 | 64 | |
| Slovinky | SL2 | 300 | 250 | 1000 | 300 |
| SLA | 4 | 8 | 300 | 64 | |
| Isolate | Strain | Acc. Number | Place of Isolation | Country | Reference |
|---|---|---|---|---|---|
| S. cadmiisoli | ZFG47 | NR_171522.1 | soil in a cadmium-contaminated area in Xiangtan City | Hunan Province, China | [71] |
| S. cyaneochromogenes | MK-45 | NR_170501.1 | manganese-contaminated area | Xiangtan, China | [68] |
| S. manganisoli | MK 44 | KY911452.1 | manganese-polluted soil, Xiangtan Manganese Mine | South Central China | [70] |
| S. plumbiresistens | CCNWHX 13-160 | EU526954 | lead-polluted soil, Gansu province | Northwest China | [60] |
| S. sporoverrucosus | dwc-3 | KC508633.1 | disposal site for (ultra-)low uraniferous radioactive waste | Southwest China | [67] |
| S. xiangtanensis | LUSFXJ | NR_164877.1 | manganese-polluted soil, Xiangtan Manganese Mine | South Central China | [72] |
| S. zinciresistens | CCNWNQ 0016 | GU225938 | zinc and copper mine, Shaanxi province | Northwest China | [61] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cimermanova, M.; Pristas, P.; Piknova, M. Biodiversity of Actinomycetes from Heavy Metal Contaminated Technosols. Microorganisms 2021, 9, 1635. https://doi.org/10.3390/microorganisms9081635
Cimermanova M, Pristas P, Piknova M. Biodiversity of Actinomycetes from Heavy Metal Contaminated Technosols. Microorganisms. 2021; 9(8):1635. https://doi.org/10.3390/microorganisms9081635
Chicago/Turabian StyleCimermanova, Michaela, Peter Pristas, and Maria Piknova. 2021. "Biodiversity of Actinomycetes from Heavy Metal Contaminated Technosols" Microorganisms 9, no. 8: 1635. https://doi.org/10.3390/microorganisms9081635
APA StyleCimermanova, M., Pristas, P., & Piknova, M. (2021). Biodiversity of Actinomycetes from Heavy Metal Contaminated Technosols. Microorganisms, 9(8), 1635. https://doi.org/10.3390/microorganisms9081635