
The Abundance and Taxonomic Diversity of Filterable Forms of Bacteria during Succession in the Soils of Antarctica (Bunger Hills)

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. Physico-Chemical Analyses of Samples
2.2.2. Scheme of the Experiment to Initiate Succession
2.2.3. Obtaining the Fraction of Filterable Forms of Bacteria
2.2.4. Determination of the Total Number of Bacteria and FFB
2.2.5. Isolation of DNA
2.2.6. Next-Generation Sequencing of the 16S rRNA Gene from Total Soil DNA
2.2.7. Amplification and Analysis of the 16S rRNA Genes from Bacterial Isolates Grown on Nutrient Media
2.2.8. Statistical Data Processing Methods
2.2.9. Depositing the Received Data
3. Results
3.1. Morphology and Composition of Soil Samples
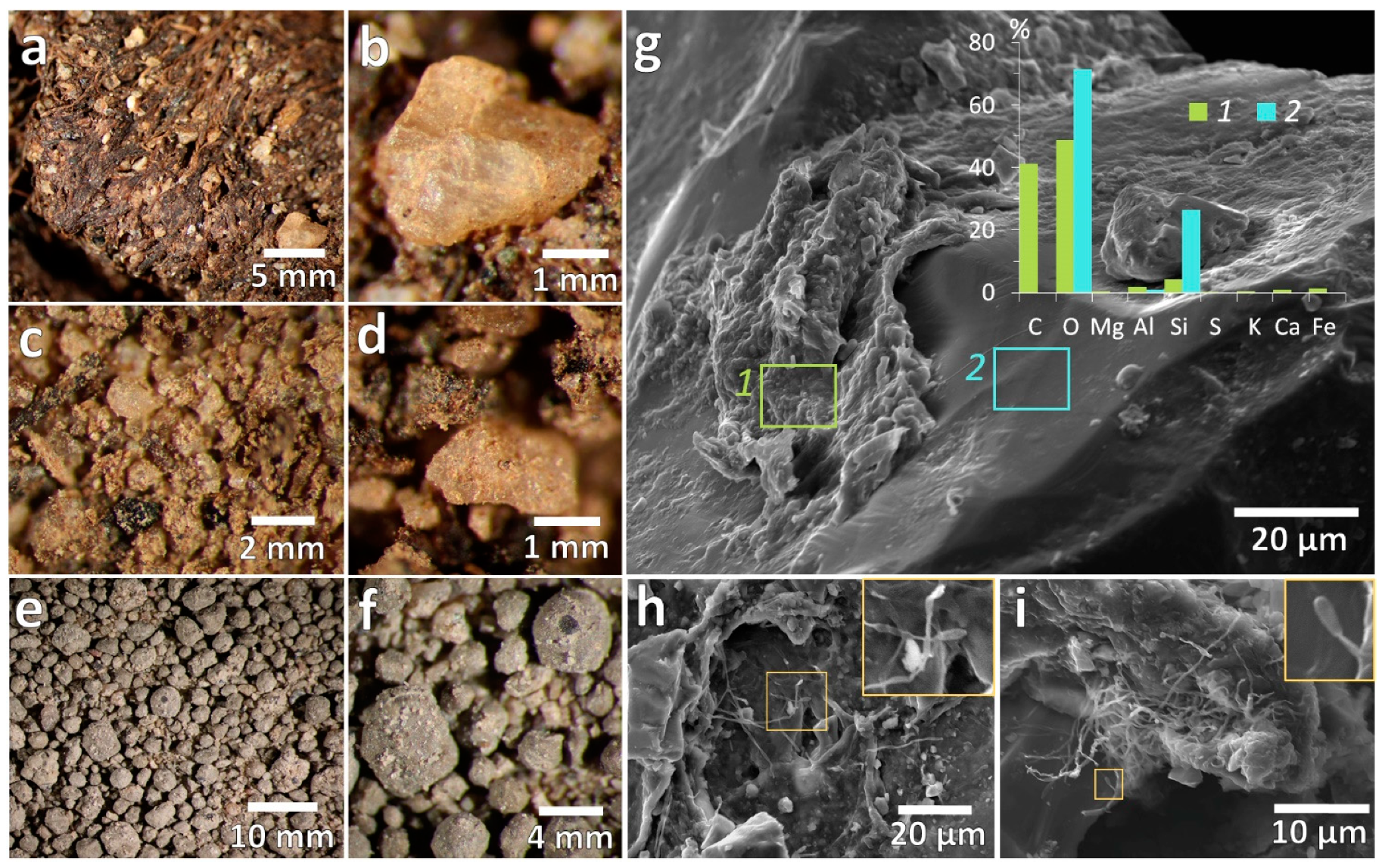
3.1.1. Mesomorphology
3.1.2. Micromorphology
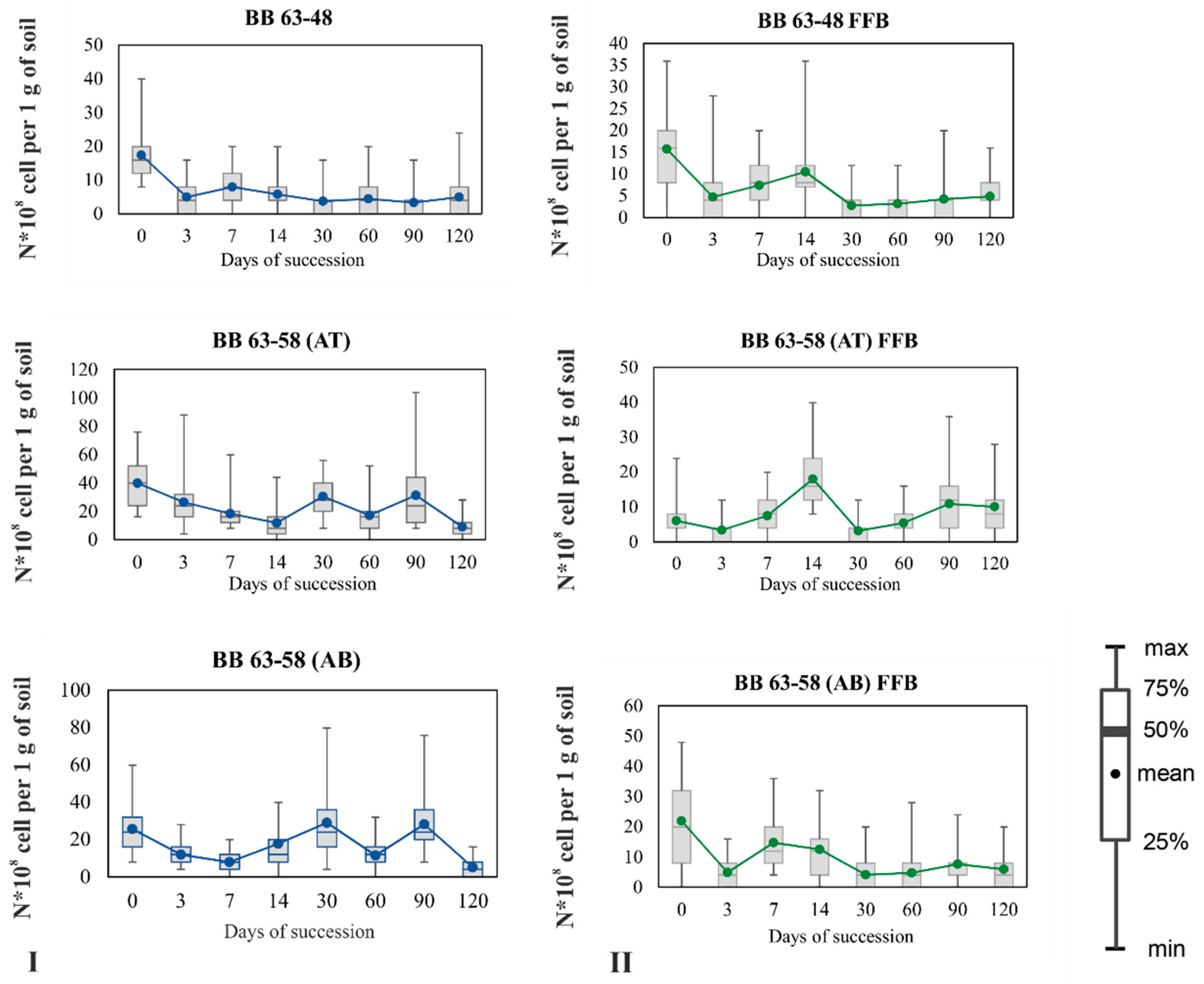
3.2. Dynamics of the Total Number of Bacteria and FFB in the Studied Soils
3.2.1. Dynamics of the Total Number of Bacteria in the Studied Soil Samples
3.2.2. Dynamics of FFB during Succession
3.3. Taxonomic Diversity of Communities in the Studied Samples
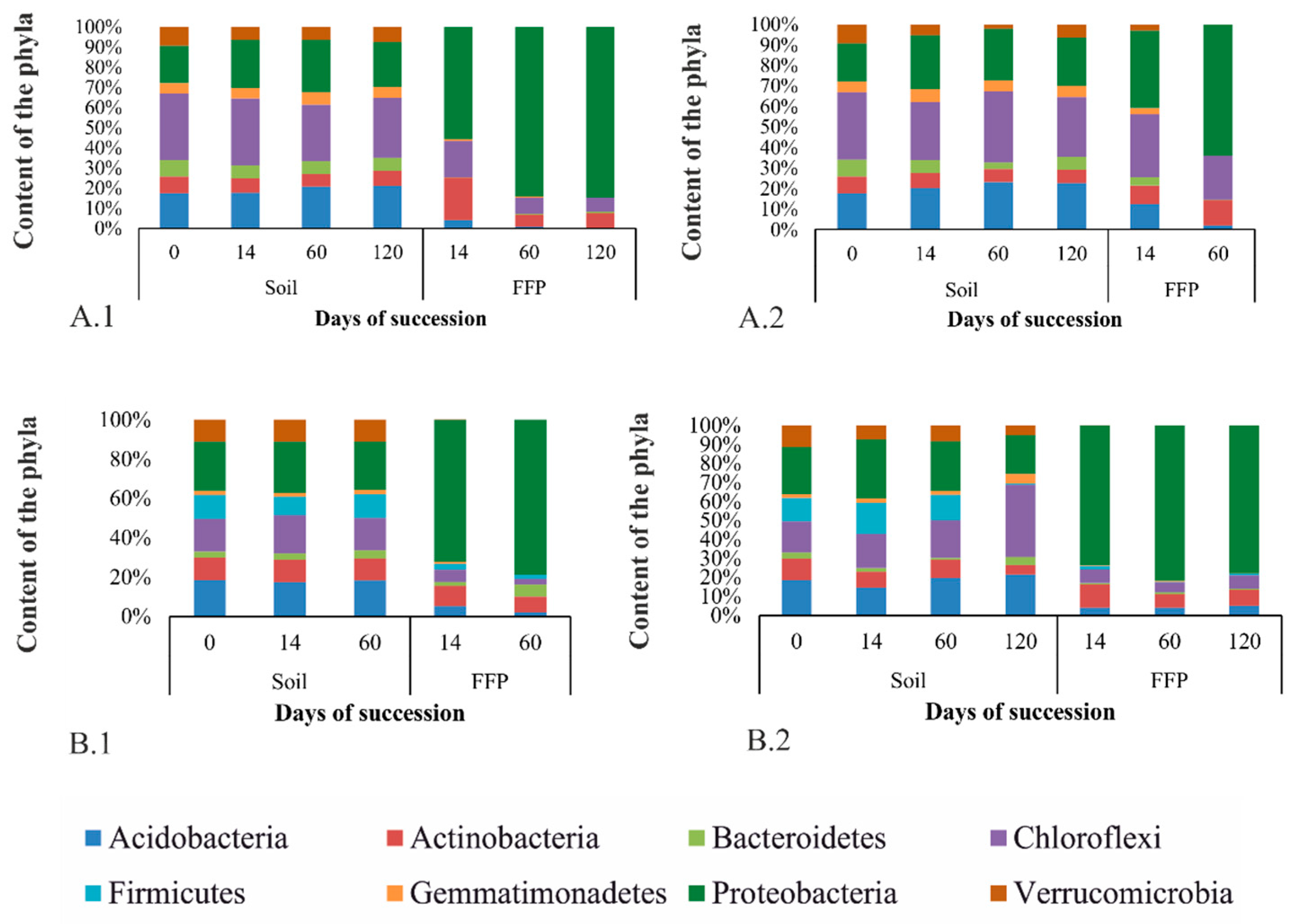
3.3.1. Taxonomic Diversity of Bacteria in the Studied Soils at the Phylum Level
3.3.2. Taxonomic Diversity of FFB in the Studied Soils at the Phylum Level
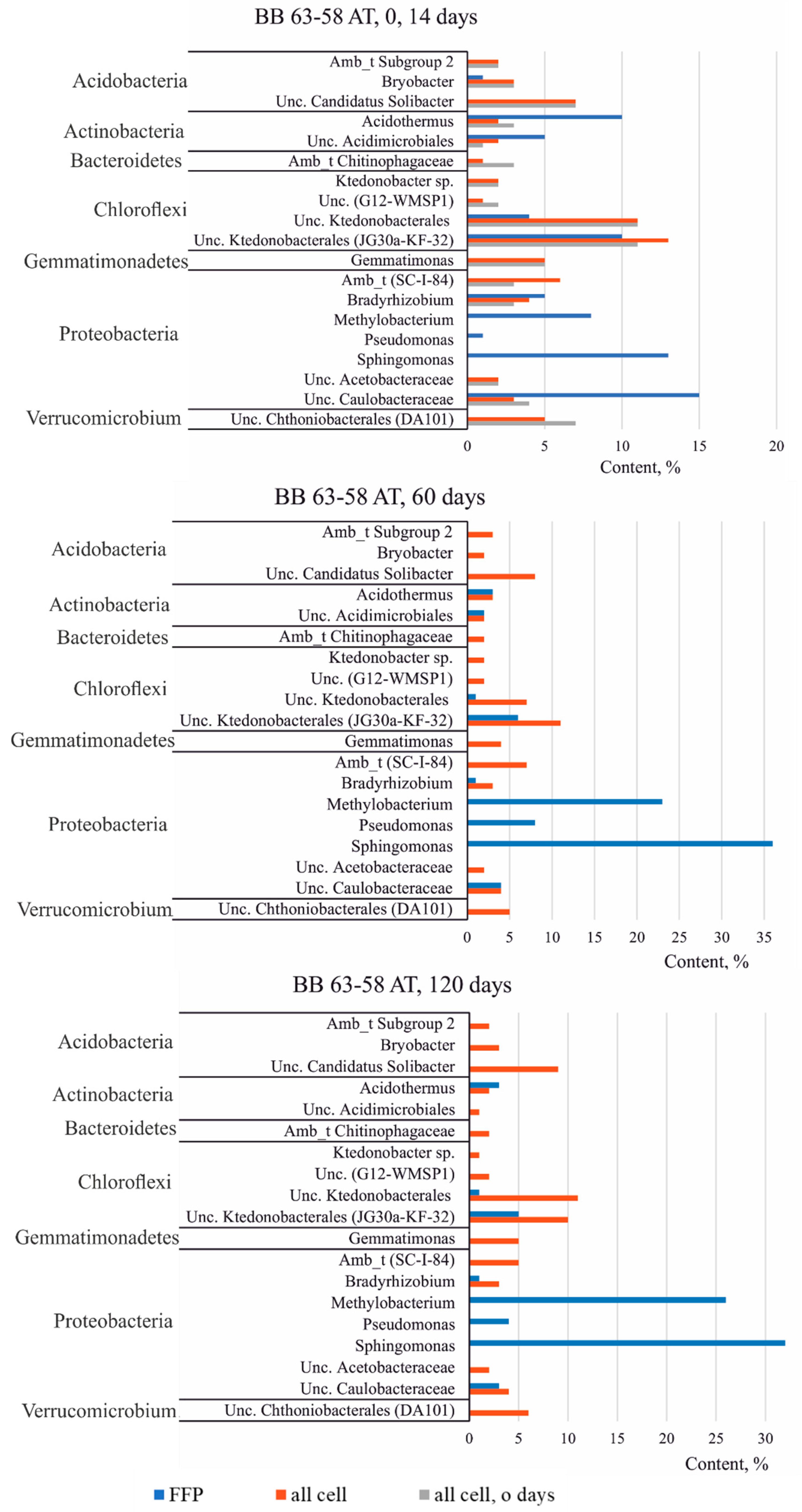
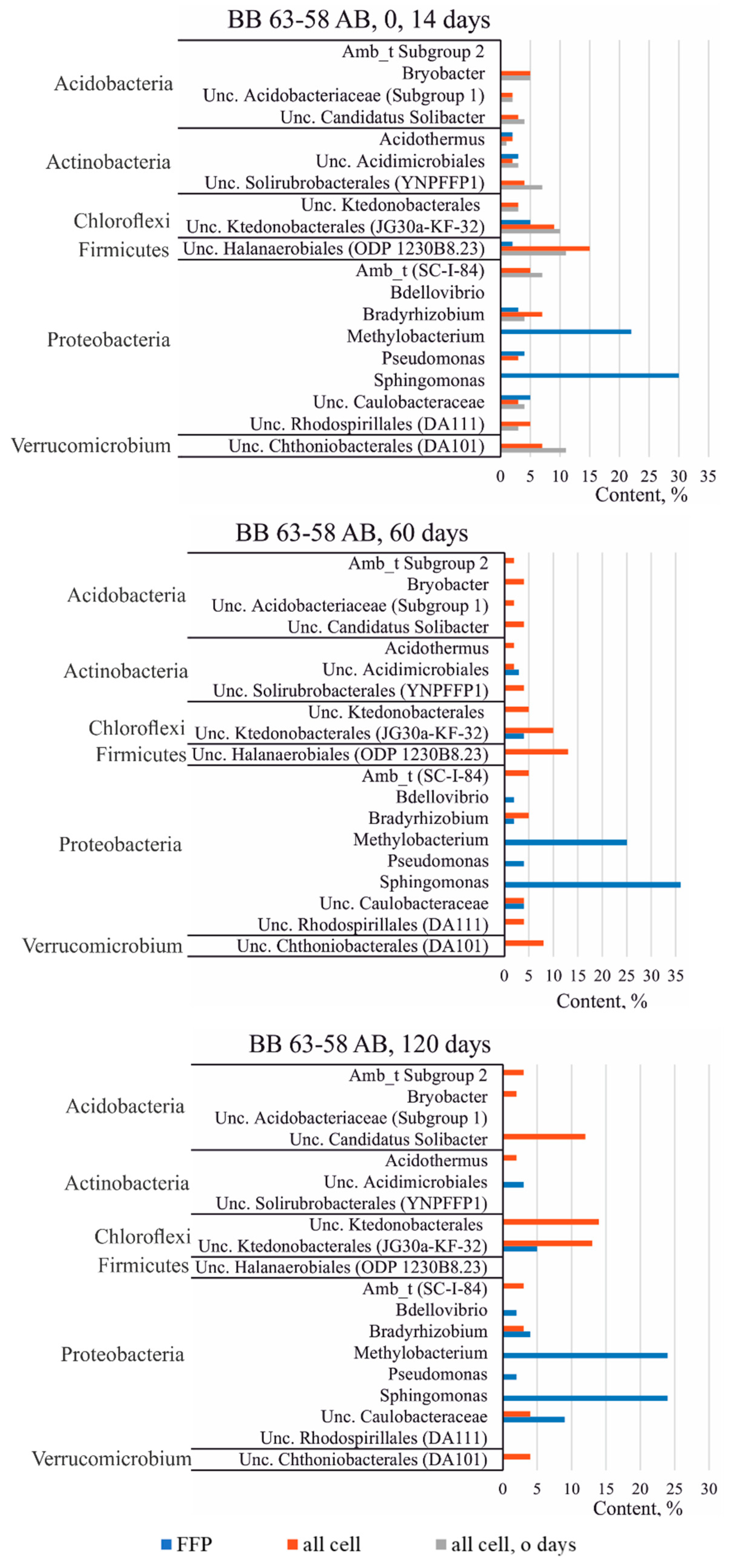
3.3.3. Taxonomic Diversity of Bacteria in the Studied Soils at the Genus Level
3.4. Analysis of Diversity Indices
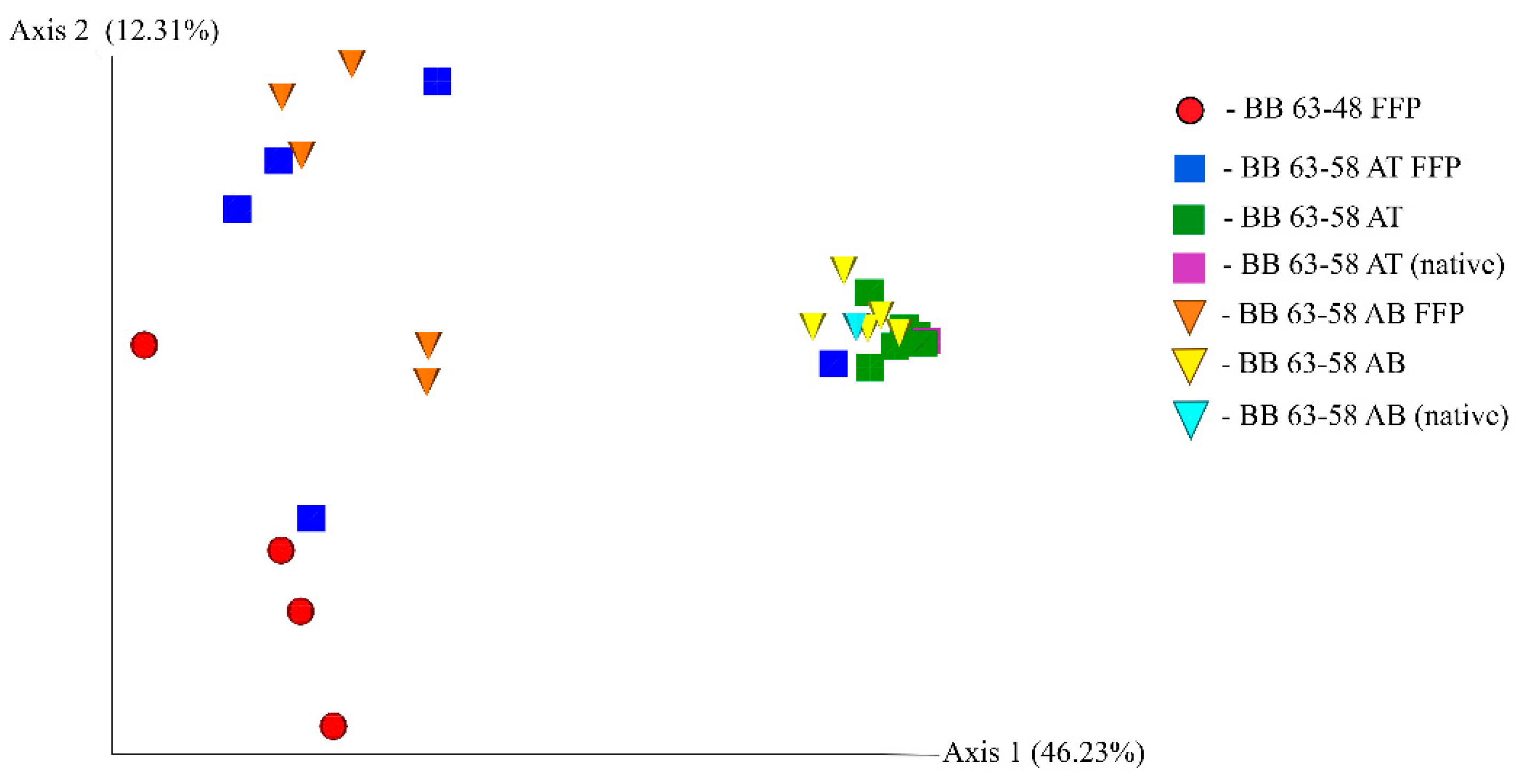
3.5. Analysis of Phylogenetic Similarity of Bacterial Communities
3.6. Alphaproteobacteria/Acidobacteria Ratio
3.7. Number and Taxonomic Diversity of Cultured Bacterial Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bej, A.K.; Mojib, N. Cold adaptation in Antarctic biodegradative microorganisms. In Polar Microbiology: The Ecology, Biodiversity and Bioremediation Potential of Microorganisms in Extremely Cold Environments; CRC Press (Taylor & Francis Group): Boca Raton, FL, USA, 2009; pp. 157–177. [Google Scholar]
- Chattopadhyay, M.K. Mechanism of bacterial adaptation to low temperature. J. Biosci. 2006, 31, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Demkina, E.V.; Soina, V.S.; Zvyagintsev, D.G.; El’-Registan, G.I. Reproductive resting forms of Arthrobacter globiformis. Mikrobiologiya 2000, 69, 377–382. [Google Scholar]
- Mulyukin, A.L.; Demkina, E.V.; Kryazhevskikh, N.A.; Galchenko, V.F.; El-Registan, G.I.; Suzina, N.E.; Duda, V.I.; Vorob’eva, L.I. Dormant forms of Micrococcus luteus and Arthrobacter globiformis not platable on standard media. Microbiology 2009, 78, 407–418. [Google Scholar] [CrossRef]
- Kriazhevskih, N.A.; Demkina, E.V.; Loĭko, N.G.; Baslerov, R.V.; Kolganova, T.V.; Soina, V.S.; Manucharova, N.A.; Gal’chenko, V.F.; Él’-Registan, G.I. Comparison of the adaptation potential of Arthrobacter oxydans and Acinetobacter lwoffii isolated from permafrost sediments with their collection analogues. Mikrobiologiia 2013, 82, 27–41. [Google Scholar] [CrossRef]
- Nakai, R. Size Matters: Ultra-small and Filterable Microorganisms in the Environment. Microbes Environ. 2020, 35, ME20025. [Google Scholar] [CrossRef] [PubMed]
- Duda, V.I.; Suzina, N.E.; Polivtseva, V.N.; Boronin, A.M. Ultramicrobacteria: Formation of the concept and contribution of ultramicrobacteria to biology. Mikrobiologiia 2012, 81, 379–390. [Google Scholar] [CrossRef]
- Zhang, Y. Persistent and dormant tubercle bacilli and latent tuberculosis. Front. Biosci. 2004, 9, 1136–1156. [Google Scholar] [CrossRef] [Green Version]
- Kudinova, A.G.; Lysak, L.V.; Soina, V.S.; Mergelov, N.S.; Dolgikh, A.V.; Shorkunov, I.G. Bacterial communities in the soils of cryptogamic barrens of East Antarctica (the Larsemann Hills and Thala Hills oases). Eurasian Soil Sci. 2015, 48, 276–287. [Google Scholar] [CrossRef]
- Lysak, L.V.; Lapygina, E.V.; Konova, I.A.; Zvyagintsev, D.G. Quantity and taxonomic composition of ultramicrobacteria in soils. Microbiology 2010, 79, 408–412. [Google Scholar] [CrossRef]
- Kudinova, A.G.; Petrova, M.A.; Dolgikh, A.V.; Soina, V.S.; Lysak, L.V.; Maslova, O.A. Taxonomic Diversity of Bacteria and Their Filterable Forms in the Soils of Eastern Antarctica (Larsemann Hills and Bunger Hills). Microbiology 2020, 89, 574–584. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of bacteria and archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; John Wiley and Sons: New York, NY, USA, 1991; pp. 125–175. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Li, Q.; Chen, X.; Jiang, Y.; Jiang, C. Morphological identification of actinobacteria. In Actinobacteria-Basics and Biotechnological Applications; InTech: Rijeka, Croatia, 2016; pp. 59–86. [Google Scholar]
- Yabe, S.; Sakai, Y.; Abe, K.; Yokota, A. Diversity of Ktedonobacteria with Actinomycetes-Like Morphology in Terrestrial Environments. Microbes Environ. 2017, 32, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Janssen, P.H. Identifying the dominant soil bacterial taxa in libraries of 16S rRNA and 16S rRNA genes. Appl. Environ. Microbiol. 2006, 72, 1719–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeBruyn, J.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global biogeography and quantitative seasonal dynamics of Gemmatimonadetes in soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [Green Version]
- Herlemann, D.P.; Lundin, D.; Labrenz, M.; Jürgens, K.; Zheng, Z.; Aspeborg, H.; Andersson, A.F. Metagenomic De novo Assembly of an aquatic representative of the verrucomicrobial class Spartobacteria. MBio 2013, 4, e0569-12. [Google Scholar] [CrossRef] [Green Version]
- Ji, M.; van Dorst, J.; Bissett, A.; Brown, M.V.; Palmer, A.S.; Snape, I.; Siciliano, S.D.; Ferrari, B.C. Microbial diversity at Mitchell Peninsula, Eastern Antarctica: Apotential biodiversity “hotspot”. Polar Biol. 2016, 39, 237–249. [Google Scholar] [CrossRef]
- Pudasaini, S.; Wilson, J.; Ji, M.; van Dorst, J.; Snape, I.; Palmer, A.S.; Burns, B.P.; Ferrari, B.C. Microbial diversity of browning Peninsula, Eastern Antarctica revealed using molecular and cultivation methods. Front. Microbiol. 2017, 8, 591. [Google Scholar] [CrossRef] [Green Version]
- Alekseev, I.; Zverev, A.; Abakumov, E. Microbial Communities in Permafrost Soils of Larsemann Hills, Eastern Antarctica: Environmental Controls and Effect of Human Impact. Microorganisms 2020, 8, 1202. [Google Scholar] [CrossRef]
- Bajerski, F.; Wagner, D. Bacterial succession in Antarctic soils of two glacier forefields on Larsemann Hills, East Antarctica. FEMS Microbiol. Ecol. 2013, 85, 128–142. [Google Scholar] [CrossRef]
- Smit, E.; Leeflang, P.; Gommans, S.; van den Broek, J.; van Mil, S.; Wernars, K. Diversity and seasonal fluctuations of the dominant members of the bacterial soil community in a wheat field as determined by cultivation and molecular methods. Appl. Environ. Microbiol. 2001, 67, 2284–2291. [Google Scholar] [CrossRef] [Green Version]
- Thomson, B.; Ostle, N.; McNamara, N.; Bailey, M.; Whiteley, A.; Griffiths, R. Vegetation affects the relative abundances of dominant soil bacterial taxa and soil respiration rates in an upland grassland soil. Microb. Ecol. 2010, 59, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Lünsdorf, H.; Erb, R.W.; Abraham, W.R.; Timmis, K.N. ‘Clay hutches’: A novel interaction between bacteria and clay minerals. Environ. Microbiol. 2000, 2, 161–168. [Google Scholar] [CrossRef]
- Fomina, M.; Skorochod, I. Microbial Interaction with Clay Minerals and Its Environmental and Biotechnological Implications. Minerals 2020, 10, 861. [Google Scholar] [CrossRef]
- Yergeau, E.; Bokhorst, S.; Kang, S.; Zhou, J.; Greer, C.W.; Aerts, R.; Kowalchuk, G.A. Shifts in soil microorganisms in response to warming are consistent across a range of Antarctic environments. ISME J. 2012, 6, 692–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newsham, K.K.; Tripathi, B.M.; Dong, K.; Yamamoto, N.; Adams, J.M.; Hopkins, D.W. Bacterial Community Composition and Diversity Respond to Nutrient Amendment but Not Warming in a Maritime Antarctic Soil. Microb. Ecol. 2019, 78, 974–984. [Google Scholar] [CrossRef] [PubMed]
- He, X.; McLean, J.S.; Edlund, A.; Yooseph, S.; Hall, A.P.; Liu, S.Y.; Dorrestein, P.C.; Esquenazi, E.; Hunter, R.C.; Cheng, G.; et al. Cultivation of a human-associated TM7 phylotype reveals a reduced genome and epibiotic parasitic lifestyle. Proc. Natl. Acad. Sci. USA 2015, 112, 244–249. [Google Scholar] [CrossRef] [Green Version]
- Hahn, M.W.; Lünsdorf, H.; Wu, Q.; Schauer, M.; Höfle, M.G.; Boenigk, J.; Stadler, P. Isolation of novel ultramicrobacteria classified as actinobacteria from five freshwater habitats in Europe and Asia. Appl. Environ. Microbiol. 2003, 69, 1442–1451. [Google Scholar] [CrossRef] [Green Version]
- Janssen, P.H.; Schuhmann, A.; Mörschel, E.; Rainey, F.A. Novel anaerobic ultramicrobacteria belonging to the Verrucomicrobiales lineage of bacterial descent isolated by dilution culture from anoxic rice paddy soil. Appl. Environ. Microbiol. 1997, 63, 1382–1388. [Google Scholar] [CrossRef] [Green Version]
- Nakai, R.; Shibuya, E.; Justel, A.; Rico, E.; Quesada, A.; Kobayashi, F.; Iwasaka, Y.; Yu, G.; Amano, Y.; Iwatsuki, T.; et al. Phylogeographic analysis of filterable bacteria with special reference to Rhizobiales strains that occur in cryospheric habitats. Antarct. Sci. 2013, 25, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Nakai, R.; Nishijima, M.; Tazato, N.; Handa, Y.; Karray, F.; Sayadi, S.; Isoda, H.; Naganuma, T. Oligoflexus tunisiensis gen. nov., sp. nov., a Gram-negative, aerobic, filamentous bacterium of a novel proteobacterial lineage, and description of Oligoflexaceae fam. nov., Oligoflexales ord. nov. and Oligoflexia classis nov. Int. J. Syst. Evol. Microbiol. 2014, 64, 3353–3359. [Google Scholar] [CrossRef]
- Hahn, M.W.; Schmidt, J.; Koll, U.; Rohde, M.; Verbarg, S.; Pitt, A.; Nakai, R.; Takeshi Naganuma, T.; Lang, E. Silvanigrella aquatica gen. nov., sp. nov., isolated from a freshwater lake, description of Silvanigrellaceae fam. nov. and Silvanigrellales ord. nov., reclassification of the order Bdellovibrionales in the class Oligoflexia, reclassification of the families Bacteriovoracaceae and Halobacteriovoraceae in the new order Bacteriovoracales ord. nov., and reclassification of the family Pseudobacteriovoracaceae in the order Oligoflexales. Int. J. Syst. Evol. Microbiol. 2017, 67, 2555–2568. [Google Scholar]
- Ryan, M.P.; Pembroke, J.T.; Adley, C.C. Ralstonia pickettii: A persistent gram-negative nosocomial infectious organism. J. Hosp. Infect. 2006, 62, 278–284. [Google Scholar] [CrossRef]
- Salter, S.J.; Cox, M.J.; Turek, E.M.; Calus, S.T.; Cookson, W.O.; Moffatt, M.F.; Turner, P.; Parkhill, J.; Loman, N.J.; Walker, A.W. Reagent and laboratory contamination can critically impact sequence-based microbiome analyses. BMC Biol. 2014, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Haller, C.M.; Rölleke, S.; Vybiral, D.; Witte, A.; Velimirov, B. Investigation of 0.2 µm filterable bacteria from the Western Mediterranean Sea using a molecular approach: Dominance of potential starvation forms. FEMS Microbiol. Ecol. 2000, 31, 153–161. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Location | Soil Profile, № | Horizon, Depth in [cm] | C | N | W |
|---|---|---|---|---|---|
| [%] | |||||
| The central part of the Hills, flat valley, fragmentary surface sodium bicarbonate crusts, Typic Haploturbel without an organogenic horizon | BB 63–48 | 10–15 | 0.13 ± 0.00 | 0.03 ± 0.00 | 4.17 ± 0.15 |
| Southern part of the Hills, wind shelter, Typic Haplorthel with an organogenic horizon | BB 63–58 | AT, 0–2 | 6.62 ± 0.01 | 0.32 ± 0.01 | 10.02 ± 0.11 |
| AB, 10–15 | 3.0 ± 0.01 | 0.40 ± 0.01 | 6.49 ± 0.20 | ||
| Sample | C [%] | N [%] | ||||
|---|---|---|---|---|---|---|
| Native | At the End of Succession (120 Days) | Native | At the End of Succession (120 Days) | |||
| 5 °C | 20 °C | 5 °C | 20 °C | |||
| BB 63–48 | 0.13 | 0.12 | 0.18 | 0.03 | 0.08 | 0.06 |
| BB 63–58 (AT) | 6.63 | 4.97 | 6.38 | 0.66 | 0.50 | 0.71 |
| BB 63–58 (AB) | 3.03 | 4.12 | 2.90 | 0.39 | 0.44 | 0.39 |
| Sample | Succession Days | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 14 | 60 | 120 | ||||||
| 5 °C | 20 °C | 5 °C | 20 °C | 5 °C | 20 °C | ||||
| BB 63–48 | Soil | - | - | - | - | - | - | - | |
| FFB | - | - | - | 0.913 | 0.909 | 0.891 | 0.912 | ||
| BB 63–58 | AT | Soil | 0.966 | 0.962 | 0.973 | 0.972 | 0.961 | 0.969 | 0.970 |
| FFB | - | 0.949 | 0.972 | 0.919 | 0.939 | 0.923 | - | ||
| AB | Soil | 0.972 | 0.970 | 0.969 | 0.972 | 0.969 | - | 0.954 | |
| FFB | - | 0.957 | 0.945 | 0.932 | 0.918 | - | 0.947 | ||
| Sample | Succession Days | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 14 | 60 | 120 | ||||||
| 5 °C | 20 °C | 5 °C | 20 °C | 5 °C | 20 °C | ||||
| BB 63–48 | Soil | - | - | - | - | - | - | - | |
| FFB | - | - | - | 5.4 | 5.2 | 4.6 | 4.8 | ||
| BB 63–58 | AT | Soil | 6.5 | 6.4 | 6.6 | 6.5 | 6.2 | 6.7 | 6.7 |
| FFB | - | 5.7 | 6.8 | 5.1 | 5.3 | 5.1 | - | ||
| AB | Soil | 6.4 | 6.4 | 6.3 | 6.4 | 6.3 | - | 6.0 | |
| FFB | - | 6.3 | 5.8 | 5.5 | 5.4 | - | 5.6 | ||
| Temperature | 5 °C | 20 °C | ||||||
|---|---|---|---|---|---|---|---|---|
| Days of Succession | 0 | 14 | 60 | 120 | 0 | 14 | 60 | 120 |
| BB 63–58 (AT) | ||||||||
| all bacteria | 0.71 | 0.82 | 0.70 | 0.60 | 0.71 | 0.84 | 0.73 | 0.62 |
| FFB fraction | - | 12.5 | 85.0 | 130.0 | - | 2.25 | 28.5 | - |
| BB 63–58 (AB) | ||||||||
| all bacteria | 0.83 | 0.76 | 0.78 | - | 0.83 | 1.43 | 0.89 | 0.71 |
| FFB fraction | - | 12.6 | 33.5 | - | - | 16.5 | 18.0 | 13.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kudinova, A.G.; Dolgih, A.V.; Mergelov, N.S.; Shorkunov, I.G.; Maslova, O.A.; Petrova, M.A. The Abundance and Taxonomic Diversity of Filterable Forms of Bacteria during Succession in the Soils of Antarctica (Bunger Hills). Microorganisms 2021, 9, 1728. https://doi.org/10.3390/microorganisms9081728
Kudinova AG, Dolgih AV, Mergelov NS, Shorkunov IG, Maslova OA, Petrova MA. The Abundance and Taxonomic Diversity of Filterable Forms of Bacteria during Succession in the Soils of Antarctica (Bunger Hills). Microorganisms. 2021; 9(8):1728. https://doi.org/10.3390/microorganisms9081728
Chicago/Turabian StyleKudinova, Alina G., Andrey V. Dolgih, Nikita S. Mergelov, Ilya G. Shorkunov, Olga A. Maslova, and Mayya A. Petrova. 2021. "The Abundance and Taxonomic Diversity of Filterable Forms of Bacteria during Succession in the Soils of Antarctica (Bunger Hills)" Microorganisms 9, no. 8: 1728. https://doi.org/10.3390/microorganisms9081728
APA StyleKudinova, A. G., Dolgih, A. V., Mergelov, N. S., Shorkunov, I. G., Maslova, O. A., & Petrova, M. A. (2021). The Abundance and Taxonomic Diversity of Filterable Forms of Bacteria during Succession in the Soils of Antarctica (Bunger Hills). Microorganisms, 9(8), 1728. https://doi.org/10.3390/microorganisms9081728