
Engagement of CEACAM1 by Helicobacter pylori HopQ Is Important for the Activation of Non-Canonical NF-κB in Gastric Epithelial Cells

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. H. pylori Strains
2.3. Infection Experiments
2.4. Binding Assay
2.5. Immunofluorescence (IF)
2.6. Immunohistochemistry (IHC)
2.7. Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. Western Blot
2.9. Quantitative Real-Time PCR (qPCR)
2.10. Statistics
3. Results
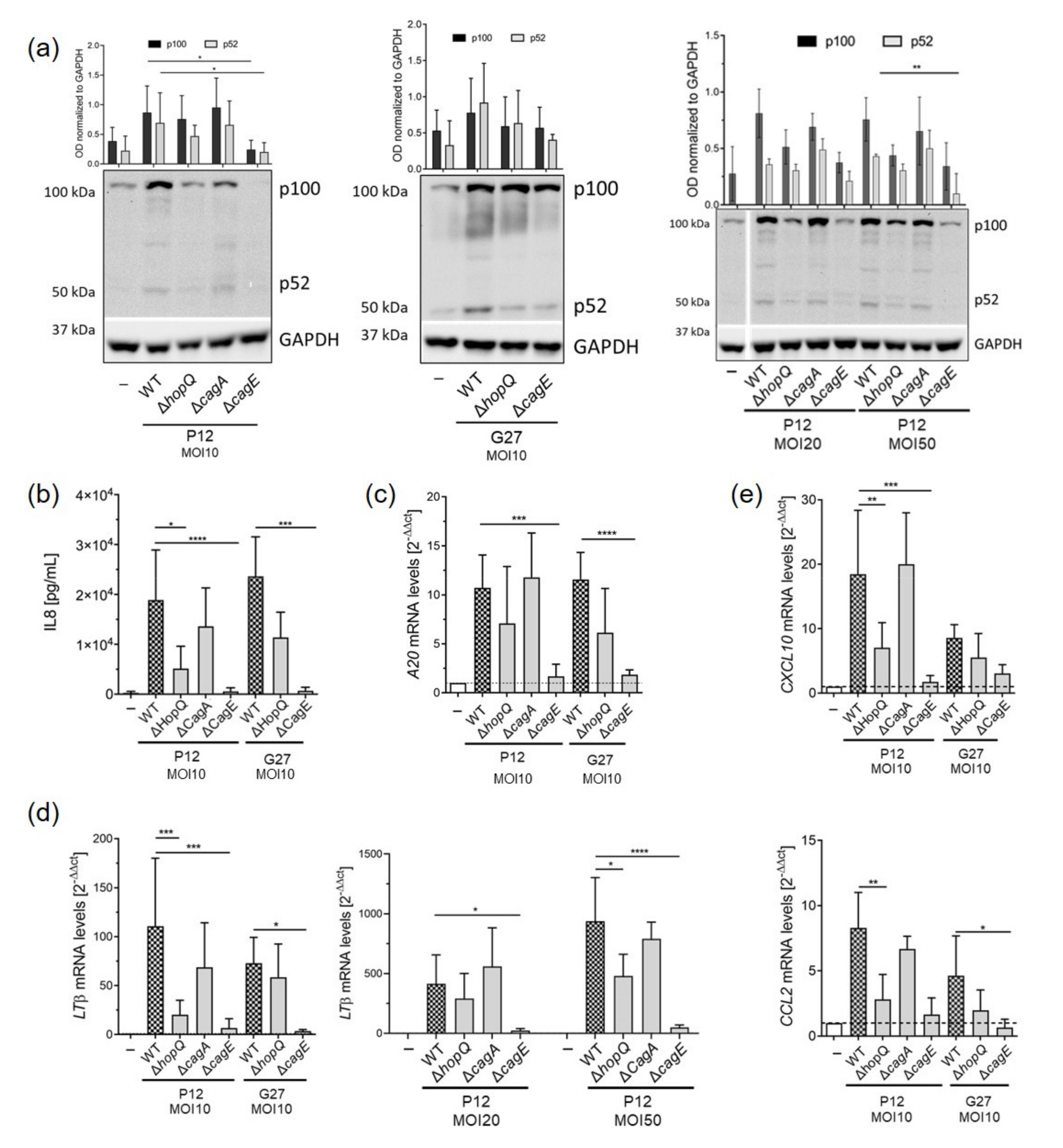
3.1. H. pylori HopQ Is Involved in the Activation of the Non-Canonical NF-κB Pathway
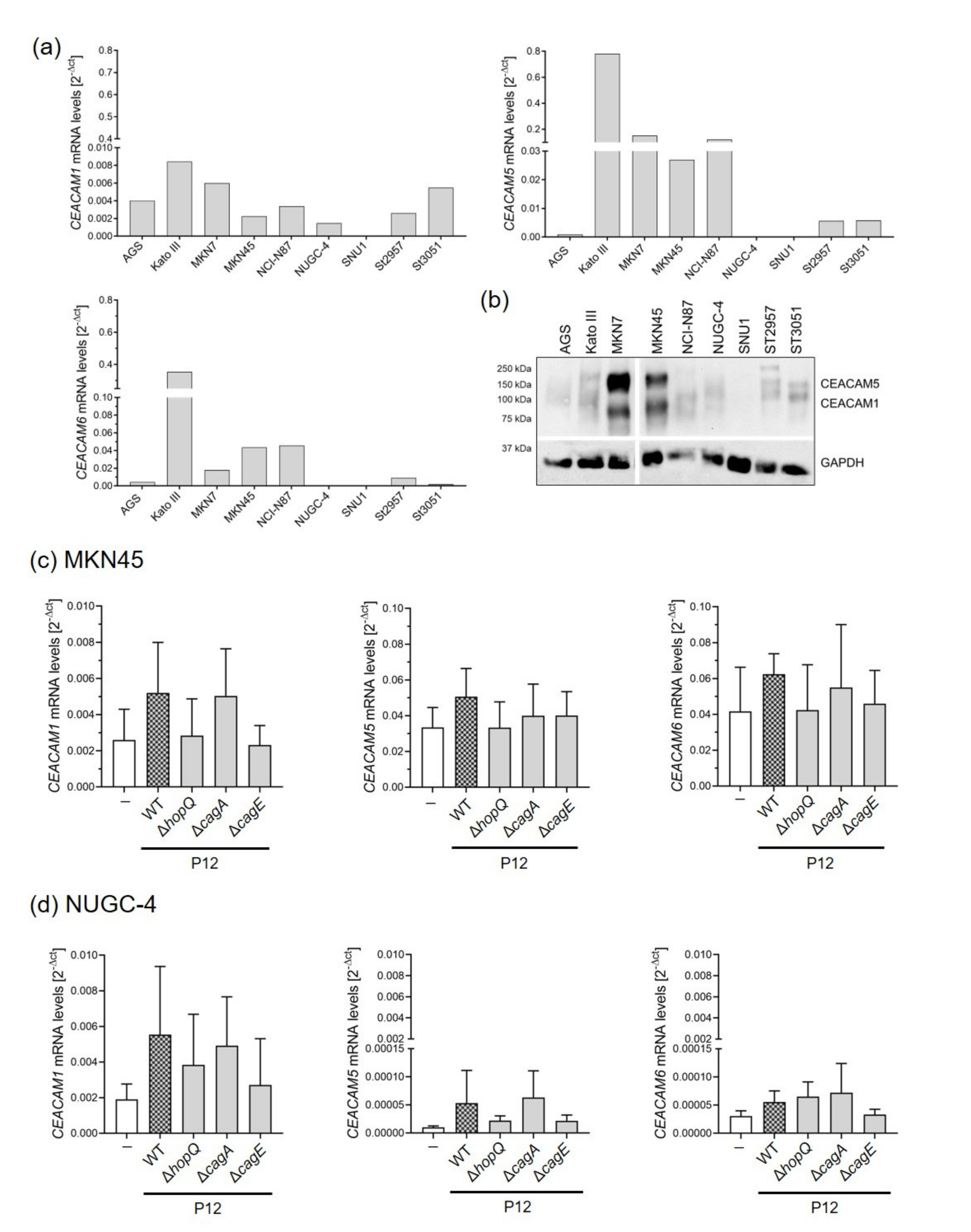
3.2. Gastric Cells Express Different Levels of CEACAMs That Are Altered upon H. pylori Infection
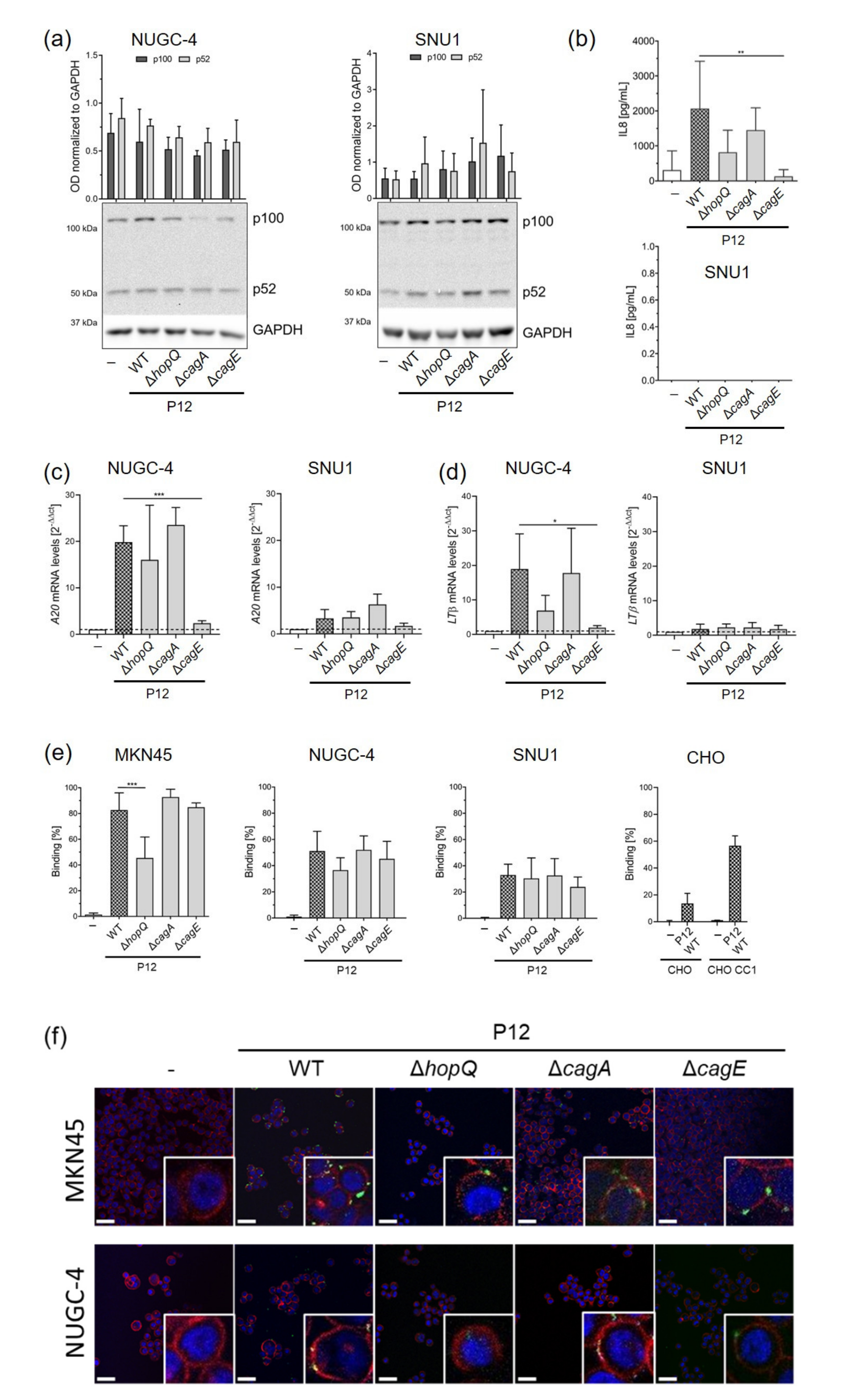
3.3. HopQ–CEACAM Interaction Is Important for the Activation of the Non-Canonical NF-κB Pathway
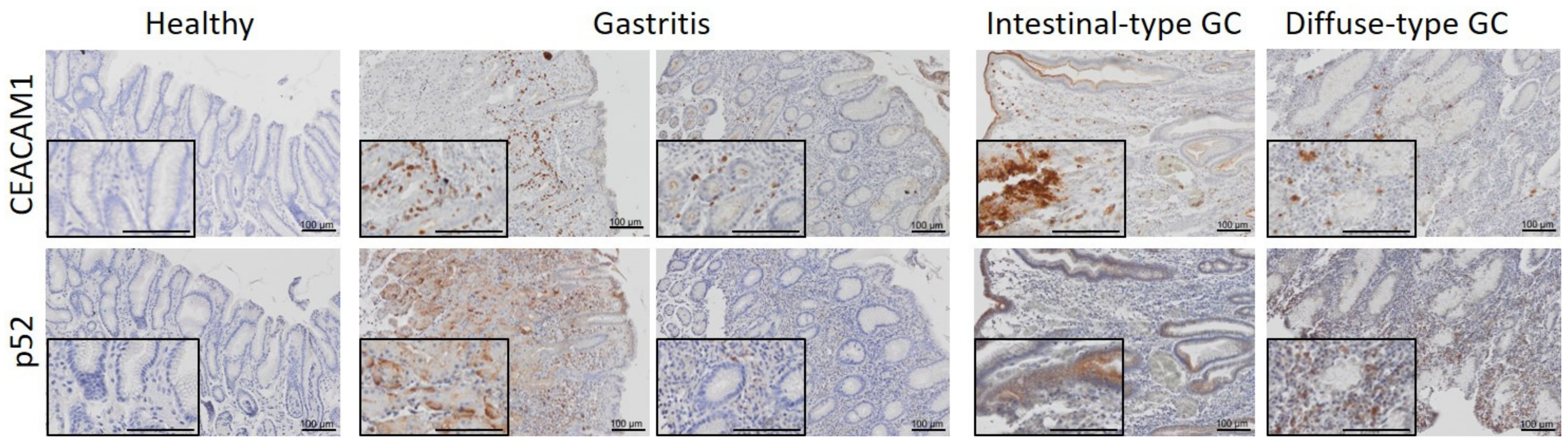
3.4. CEACAM1 Expression and Activation of the Non-Canonical NF-κB Pathway Positively Correlate in Human Gastritis and Gastric Cancer Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hooi, J.K.Y.; Lai, W.Y.; Ng, W.K.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; et al. Global Prevalence of Helicobacter pylori Infection: Systematic Review and Meta-Analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef] [Green Version]
- Takahashi-Kanemitsu, A.; Knight, C.T.; Hatakeyama, M. Molecular anatomy and pathogenic actions of Helicobacter pylori CagA that underpin gastric carcinogenesis. Cell Mol. Immunol. 2020, 17, 50–63. [Google Scholar] [CrossRef] [Green Version]
- Cover, T.L. Helicobacter pylori Diversity and Gastric Cancer Risk. MBio 2016, 7, e01869-15. [Google Scholar] [CrossRef] [Green Version]
- Santacroce, L.; Cagiano, R.; Del Prete, R.; Bottalico, L.; Sabatini, R.; Carlaio, R.G.; Prejbeanu, R.; Vermesan, H.; Dragulescu, S.I.; Vermesan, D.; et al. Helicobacter pylori infection and gastric MALTomas: An up-to-date and therapy highlight. Clin. Ter. 2008, 159, 457–462. [Google Scholar]
- Craig, V.J.; Arnold, I.; Gerke, C.; Huynh, M.Q.; Wundisch, T.; Neubauer, A.; Renner, C.; Falkow, S.; Muller, A. Gastric MALT lymphoma B cells express polyreactive, somatically mutated immunoglobulins. Blood 2010, 115, 581–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.P.; Zhu, Y.L.; Shao, W. Role of Helicobacter pylori virulence factor cytotoxin-associated gene A in gastric mucosa-associated lymphoid tissue lymphoma. World J. Gastroenterol. 2013, 19, 8219–8226. [Google Scholar] [CrossRef]
- Bailey, P.; Chang, D.K.; Nones, K.; Johns, A.L.; Patch, A.M.; Gingras, M.C.; Miller, D.K.; Christ, A.N.; Bruxner, T.J.; Quinn, M.C.; et al. Genomic analyses identify molecular subtypes of pancreatic cancer. Nature 2016, 531, 47–52. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer. Schistosomes, Liver Flukes and Helicobacter Pylori; IARC: Lyon, France, 1994; Volume 61, pp. 1–241. [Google Scholar]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Infection with Helicobacter Pylori; IARC: Lyon, France, 1994; Volume 61, pp. 177–240. [Google Scholar]
- Mahmoudian, R.A.; Gharaie, M.L.; Abbaszadegan, R.; Forghanifard, M.M.; Abbaszadegan, M.R. Interaction between LINC-ROR and Stemness State in Gastric Cancer Cells with Helicobacter pylori Infection. Iran. Biomed. J. 2021, 25, 157–168. [Google Scholar]
- Zhang, J.; Wei, J.; Wang, Z.; Feng, Y.; Wei, Z.; Hou, X.; Xu, J.; He, Y.; Yang, D. Transcriptome hallmarks in Helicobacter pylori infection influence gastric cancer and MALT lymphoma. Epigenomics 2020, 12, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Correa, P. Human gastric carcinogenesis: A multistep and multifactorial process--First American Cancer Society Award Lecture on Cancer Epidemiology and Prevention. Cancer Res. 1992, 52, 6735–6740. [Google Scholar]
- Duarte, H.O.; Gomes, J.; Machado, J.C.; Reis, C.A. Gastric cancer: Basic aspects. Helicobacter 2018, 23 (Suppl. 1), e12523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterbenc, A.; Jarc, E.; Poljak, M.; Homan, M. Helicobacter pylori virulence genes. World J. Gastroenterol. 2019, 25, 4870–4884. [Google Scholar] [CrossRef] [PubMed]
- Wroblewski, L.E.; Peek, R.M.; Wilson, K.T. Helicobacter pylori and gastric cancer: Factors that modulate disease risk. Clin. Microbiol. Rev. 2010, 23, 713–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polk, D.B.; Peek, R.M. Helicobacter pylori: Gastric cancer and beyond. Nat. Rev. Cancer 2010, 10, 403–414. [Google Scholar] [CrossRef] [Green Version]
- Backert, S.; Clyne, M.; Tegtmeyer, N. Molecular mechanisms of gastric epithelial cell adhesion and injection of CagA by Helicobacter pylori. Cell Commun. Signal. 2011, 9, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javaheri, A.; Kruse, T.; Moonens, K.; Mejias-Luque, R.; Debraekeleer, A.; Asche, C.I.; Tegtmeyer, N.; Kalali, B.; Bach, N.C.; Sieber, S.A.; et al. Helicobacter pylori adhesin HopQ engages in a virulence-enhancing interaction with human CEACAMs. Nat. Microbiol. 2016, 2, 16189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koniger, V.; Holsten, L.; Harrison, U.; Busch, B.; Loell, E.; Zhao, Q.; Bonsor, D.A.; Roth, A.; Kengmo-Tchoupa, A.; Smith, S.I.; et al. Erratum: Helicobacter pylori exploits human CEACAMs via HopQ for adherence and translocation of CagA. Nat. Microbiol. 2016, 2, 16233. [Google Scholar] [CrossRef] [Green Version]
- Moonens, K.; Hamway, Y.; Neddermann, M.; Reschke, M.; Tegtmeyer, N.; Kruse, T.; Kammerer, R.; Mejias-Luque, R.; Singer, B.B.; Backert, S.; et al. Helicobacter pylori adhesin HopQ disrupts trans dimerization in human CEACAMs. EMBO J. 2018, 37, e98665. [Google Scholar] [CrossRef]
- Mueller, D.; Tegtmeyer, N.; Brandt, S.; Yamaoka, Y.; De Poire, E.; Sgouras, D.; Wessler, S.; Torres, J.; Smolka, A.; Backert, S. c-Src and c-Abl kinases control hierarchic phosphorylation and function of the CagA effector protein in Western and East Asian Helicobacter pylori strains. J. Clin. Investig. 2012, 122, 1553–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tegtmeyer, N.; Backert, S. Role of Abl and Src family kinases in actin-cytoskeletal rearrangements induced by the Helicobacter pylori CagA protein. Eur. J. Cell Biol. 2011, 90, 880–890. [Google Scholar] [CrossRef] [PubMed]
- Fischer, W. Assembly and molecular mode of action of the Helicobacter pylori Cag type IV secretion apparatus. FEBS J. 2011, 278, 1203–1212. [Google Scholar] [CrossRef] [Green Version]
- Qu, C.K. The SHP-2 tyrosine phosphatase: Signaling mechanisms and biological functions. Cell Res. 2000, 10, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Lamb, A.; Chen, L.F. The many roads traveled by Helicobacter pylori to NFkappaB activation. Gut Microbes 2010, 1, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Lamb, A.; Chen, L.F. Role of the Helicobacter pylori-induced inflammatory response in the development of gastric cancer. J. Cell. Biochem. 2013, 114, 491–497. [Google Scholar] [CrossRef] [Green Version]
- Schweitzer, K.; Sokolova, O.; Bozko, P.M.; Naumann, M. Helicobacter pylori induces NF-kappaB independent of CagA. EMBO Rep. 2010, 11, 10–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feige, M.H.; Sokolova, O.; Pickenhahn, A.; Maubach, G.; Naumann, M. HopQ impacts the integrin alpha5beta1-independent NF-kappaB activation by Helicobacter pylori in CEACAM expressing cells. Int. J. Med. Microbiol. 2018, 308, 527–533. [Google Scholar] [CrossRef]
- Rahman, M.M.; McFadden, G. Modulation of NF-kappaB signalling by microbial pathogens. Nat. Rev. Microbiol. 2011, 9, 291–306. [Google Scholar] [CrossRef]
- Gilmore, T.D. Introduction to NF-kappaB: Players, pathways, perspectives. Oncogene 2006, 25, 6680–6684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mejias-Luque, R.; Zoller, J.; Anderl, F.; Loew-Gil, E.; Vieth, M.; Adler, T.; Engler, D.B.; Urban, S.; Browning, J.L.; Muller, A.; et al. Lymphotoxin beta receptor signalling executes Helicobacter pylori-driven gastric inflammation in a T4SS-dependent manner. Gut 2017, 66, 1369–1381. [Google Scholar] [CrossRef]
- Feige, M.H.; Vieth, M.; Sokolova, O.; Tager, C.; Naumann, M. Helicobacter pylori induces direct activation of the lymphotoxin beta receptor and non-canonical nuclear factor-kappa B signaling. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Mejias-Luque, R.; Lozano-Pope, I.; Wanisch, A.; Heikenwalder, M.; Gerhard, M.; Obonyo, M. Increased LIGHT expression and activation of non-canonical NF-kappaB are observed in gastric lesions of MyD88-deficient mice upon Helicobacter felis infection. Sci. Rep. 2019, 9, 7030. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.C. The noncanonical NF-kappaB pathway. Immunol. Rev. 2012, 246, 125–140. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.C. Non-canonical NF-kappaB signaling pathway. Cell Res. 2011, 21, 71–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tegtmeyer, N.; Ghete, T.D.; Schmitt, V.; Remmerbach, T.; Cortes, M.C.C.; Bondoc, E.M.; Graf, H.L.; Singer, B.B.; Hirsch, C.; Backert, S. Type IV secretion of Helicobacter pylori CagA into oral epithelial cells is prevented by the absence of CEACAM receptor expression. Gut Pathog. 2020, 12, 25. [Google Scholar] [CrossRef]
- Belogolova, E.; Bauer, B.; Pompaiah, M.; Asakura, H.; Brinkman, V.; Ertl, C.; Bartfeld, S.; Nechitaylo, T.Y.; Haas, R.; Machuy, N.; et al. Helicobacter pylori outer membrane protein HopQ identified as a novel T4SS-associated virulence factor. Cell. Microbiol. 2013, 15, 1896–1912. [Google Scholar] [PubMed] [Green Version]
- Hamway, Y.; Taxauer, K.; Moonens, K.; Neumeyer, V.; Fischer, W.; Schmitt, V.; Singer, B.B.; Remaut, H.; Gerhard, M.; Mejias-Luque, R. Cysteine Residues in Helicobacter pylori Adhesin HopQ are Required for CEACAM-HopQ Interaction and Subsequent CagA Translocation. Microorganisms 2020, 8, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, R.; Meyer, T.F.; van Putten, J.P. Aflagellated mutants of Helicobacter pylori generated by genetic transformation of naturally competent strains using transposon shuttle mutagenesis. Mol. Microbiol. 1993, 8, 753–760. [Google Scholar] [CrossRef]
- Xiang, Z.; Censini, S.; Bayeli, P.F.; Telford, J.L.; Figura, N.; Rappuoli, R.; Covacci, A. Analysis of expression of CagA and VacA virulence factors in 43 strains of Helicobacter pylori reveals that clinical isolates can be divided into two major types and that CagA is not necessary for expression of the vacuolating cytotoxin. Infect. Immun. 1995, 63, 94–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Censini, S.; Lange, C.; Xiang, Z.; Crabtree, J.E.; Ghiara, P.; Borodovsky, M.; Rappuoli, R.; Covacci, A. cag, a pathogenicity island of Helicobacter pylori, encodes type I-specific and disease-associated virulence factors. Proc. Natl. Acad. Sci. USA 1996, 93, 14648–14653. [Google Scholar] [CrossRef] [Green Version]
- Backert, S.; Muller, E.C.; Jungblut, P.R.; Meyer, T.F. Tyrosine phosphorylation patterns and size modification of the Helicobacter pylori CagA protein after translocation into gastric epithelial cells. Proteomics 2001, 1, 608–617. [Google Scholar] [CrossRef]
- Shariq, M.; Kumar, N.; Kumari, R.; Kumar, A.; Subbarao, N.; Mukhopadhyay, G. Biochemical Analysis of CagE: A VirB4 Homologue of Helicobacter pylori Cag-T4SS. PLoS ONE 2015, 10, e0142606. [Google Scholar] [CrossRef] [PubMed]
- Pavlopoulou, A.; Scorilas, A. A comprehensive phylogenetic and structural analysis of the carcinoembryonic antigen (CEA) gene family. Genome Biol. Evol. 2014, 6, 1314–1326. [Google Scholar] [CrossRef] [Green Version]
- Obrink, B. CEA adhesion molecules: Multifunctional proteins with signal-regulatory properties. Curr. Opin. Cell Biol. 1997, 9, 616–626. [Google Scholar] [CrossRef]
- Gebauer, F.; Wicklein, D.; Horst, J.; Sundermann, P.; Maar, H.; Streichert, T.; Tachezy, M.; Izbicki, J.R.; Bockhorn, M.; Schumacher, U. Carcinoembryonic antigen-related cell adhesion molecules (CEACAM) 1, 5 and 6 as biomarkers in pancreatic cancer. PLoS ONE 2014, 9, e113023. [Google Scholar] [CrossRef] [Green Version]
- Hammarstrom, S. The carcinoembryonic antigen (CEA) family: Structures, suggested functions and expression in normal and malignant tissues. Semin. Cancer Biol. 1999, 9, 67–81. [Google Scholar] [CrossRef]
- Yu, Q.; Wang, L.C.; Di Benigno, S.; Gray-Owen, S.D.; Stein, D.C.; Song, W. Neisseria gonorrhoeae infects the heterogeneous epithelia of the human cervix using distinct mechanisms. PLoS Pathog. 2019, 15, e1008136. [Google Scholar] [CrossRef] [Green Version]
- Kalali, B.; Mejias-Luque, R.; Javaheri, A.; Gerhard, M. H. pylori virulence factors: Influence on immune system and pathology. Mediat. Inflamm. 2014, 2014, 426309. [Google Scholar] [CrossRef] [Green Version]
- Maubach, G.; Sokolova, O.; Tager, C.; Naumann, M. CEACAMs interaction with Helicobacter pylori HopQ supports the type 4 secretion system-dependent activation of non-canonical NF-kappaB. Int. J. Med. Microbiol. 2020, 310, 151444. [Google Scholar] [CrossRef]
- Guo, J.Q.; Yu, W.H.; Wang, H.J.; Liu, B.; Zhu, K.X.; Zhang, Q.H.; Zhang, T.G.; Xu, W.H.; Wang, H.B.; Wu, H.L.; et al. Different expression patterns of CEACAM1 and its impacts on angiogenesis in gastric nonneoplastic and neoplastic lesions. Ann. Surg. Oncol. 2012, 19 (Suppl. 3), S365–S374. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.F.; Xu, S.X.; He, P.; Xi, Z.H. Expression of carcinoembryonic antigen-related cell adhesion molecule 1(CEACAM1) and its correlation with angiogenesis in gastric cancer. Pathol. Res. Pract. 2014, 210, 473–476. [Google Scholar] [CrossRef]
- Frick, A.G.; Joseph, T.D.; Pang, L.; Rabe, A.M.; St Geme, J.W., 3rd; Look, D.C. Haemophilus influenzae stimulates ICAM-1 expression on respiratory epithelial cells. J. Immunol. 2000, 164, 4185–4196. [Google Scholar] [CrossRef] [PubMed]
- Avadhanula, V.; Rodriguez, C.A.; Ulett, G.C.; Bakaletz, L.O.; Adderson, E.E. Nontypeable Haemophilus influenzae adheres to intercellular adhesion molecule 1 (ICAM-1) on respiratory epithelial cells and upregulates ICAM-1 expression. Infect. Immun. 2006, 74, 830–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnich, N.; Carvalho, F.A.; Glasser, A.L.; Darcha, C.; Jantscheff, P.; Allez, M.; Peeters, H.; Bommelaer, G.; Desreumaux, P.; Colombel, J.F.; et al. CEACAM6 acts as a receptor for adherent-invasive E. coli, supporting ileal mucosa colonization in Crohn disease. J. Clin. Investig. 2007, 117, 1566–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, M.J.; Seleznik, G.M.; Zeller, N.; Heikenwalder, M. The unexpected role of lymphotoxin beta receptor signaling in carcinogenesis: From lymphoid tissue formation to liver and prostate cancer development. Oncogene 2010, 29, 5006–5018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, M.T.; Dejardin, E.; dos Santos, N.R. Context-dependent roles for lymphotoxin-beta receptor signaling in cancer development. Biochim. Biophys. Acta 2016, 1865, 204–219. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| LTβ | GAG GAC TGG TAA CGG AGA CG | GGG CTG AGA TCT GTT TCT GG |
| A20 | TCC TCA GGC TTT GTA TTT GAG C | TCT CCC GTA TCT TCA CAG CTT |
| CEACAM1 | GCA ACA GGA CCA CAG TCA AG | CCA GGG CTA CTG CTA TCA G |
| CEACAM5 | AGG CCA ATA ACT CAG CCA GT [36] | GGC TTG GGC AGC TCC GC |
| CEACAM6 | CGT CGG CAT CAC GAT TGG [37] | TGG GAT TGG AGG AGC TAG AAG [37] |
| CCL2 | CTT CGG AGT TTG GGT TTG CTT | CAT TGT GGC CAA GGA GAT CTG |
| CXCL10 | TAT TCC TGC AAG CCA ATT TTG TC | TCT TGA TGG CCT TCG ATT CTG |
| GAPDH | GAA GGT GAA GGT CGG AGT | GAA GAT GGT GAT GGG ATT TC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taxauer, K.; Hamway, Y.; Ralser, A.; Dietl, A.; Mink, K.; Vieth, M.; Singer, B.B.; Gerhard, M.; Mejías-Luque, R. Engagement of CEACAM1 by Helicobacter pylori HopQ Is Important for the Activation of Non-Canonical NF-κB in Gastric Epithelial Cells. Microorganisms 2021, 9, 1748. https://doi.org/10.3390/microorganisms9081748
Taxauer K, Hamway Y, Ralser A, Dietl A, Mink K, Vieth M, Singer BB, Gerhard M, Mejías-Luque R. Engagement of CEACAM1 by Helicobacter pylori HopQ Is Important for the Activation of Non-Canonical NF-κB in Gastric Epithelial Cells. Microorganisms. 2021; 9(8):1748. https://doi.org/10.3390/microorganisms9081748
Chicago/Turabian StyleTaxauer, Karin, Youssef Hamway, Anna Ralser, Alisa Dietl, Karin Mink, Michael Vieth, Bernhard B. Singer, Markus Gerhard, and Raquel Mejías-Luque. 2021. "Engagement of CEACAM1 by Helicobacter pylori HopQ Is Important for the Activation of Non-Canonical NF-κB in Gastric Epithelial Cells" Microorganisms 9, no. 8: 1748. https://doi.org/10.3390/microorganisms9081748
APA StyleTaxauer, K., Hamway, Y., Ralser, A., Dietl, A., Mink, K., Vieth, M., Singer, B. B., Gerhard, M., & Mejías-Luque, R. (2021). Engagement of CEACAM1 by Helicobacter pylori HopQ Is Important for the Activation of Non-Canonical NF-κB in Gastric Epithelial Cells. Microorganisms, 9(8), 1748. https://doi.org/10.3390/microorganisms9081748