
Update on the Mechanisms of Antibiotic Resistance and the Mobile Resistome in the Emerging Zoonotic Pathogen Streptococcus suis

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Antibiotics Used to Treat S. suis Infections in Pig Production
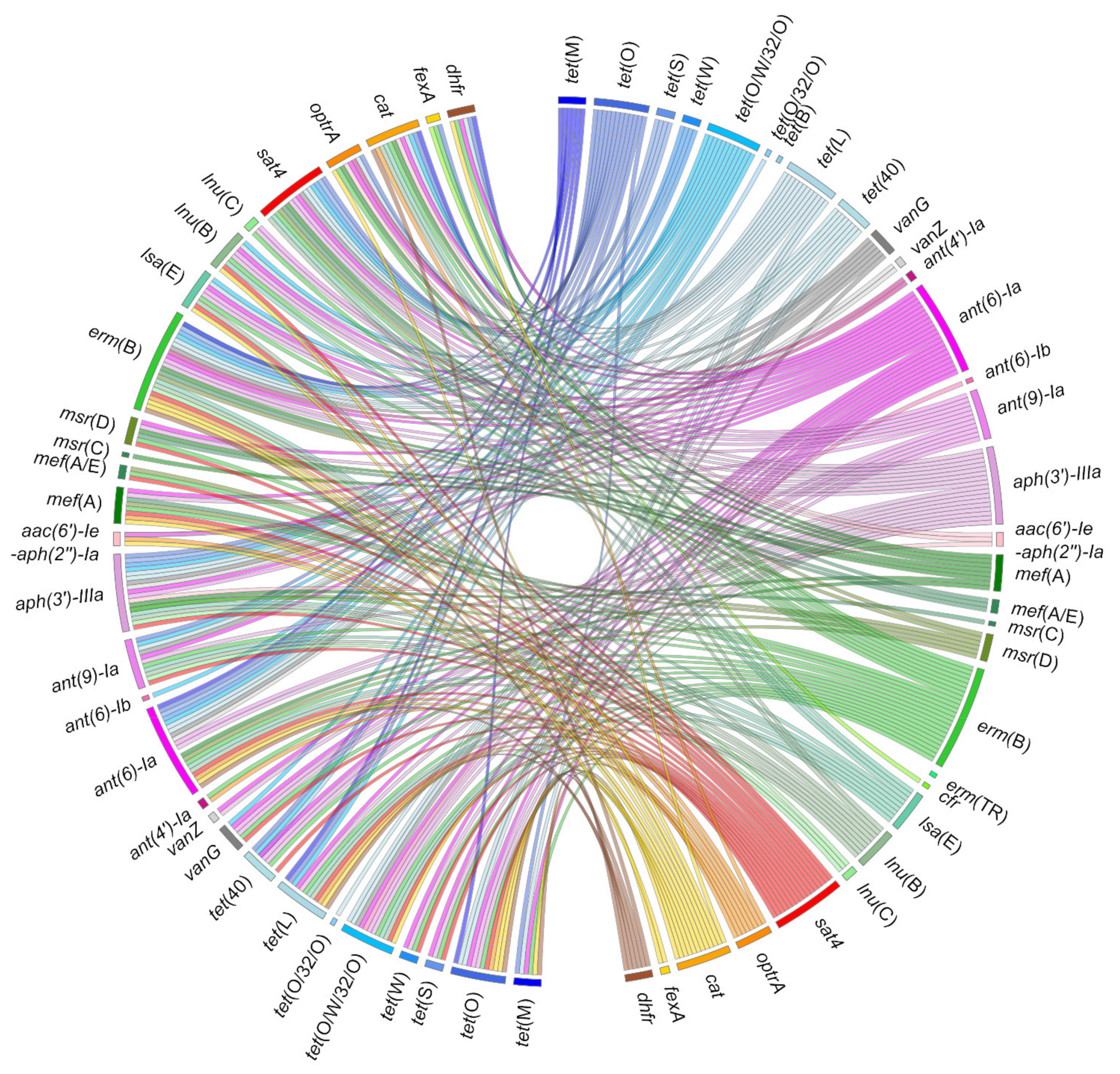
3. Mechanisms of Antimicrobial Resistance (AMR) and Resistance Genes Found in S. suis
3.1. Resistance to Tetracyclines
3.2. Resistance to Macrolides and Lincosamides
3.3. Resistance to Aminoglycosides
3.4. Resistance to Streptothricin
3.5. Resistance to β-Lactams
3.6. Resistance to Fluoroquinolones
3.7. Resistance to Glycopeptides
3.8. Resistance to Amphenicols
3.9. Resistance to Bacitracin
3.10. Resistance to Oxazolidinones
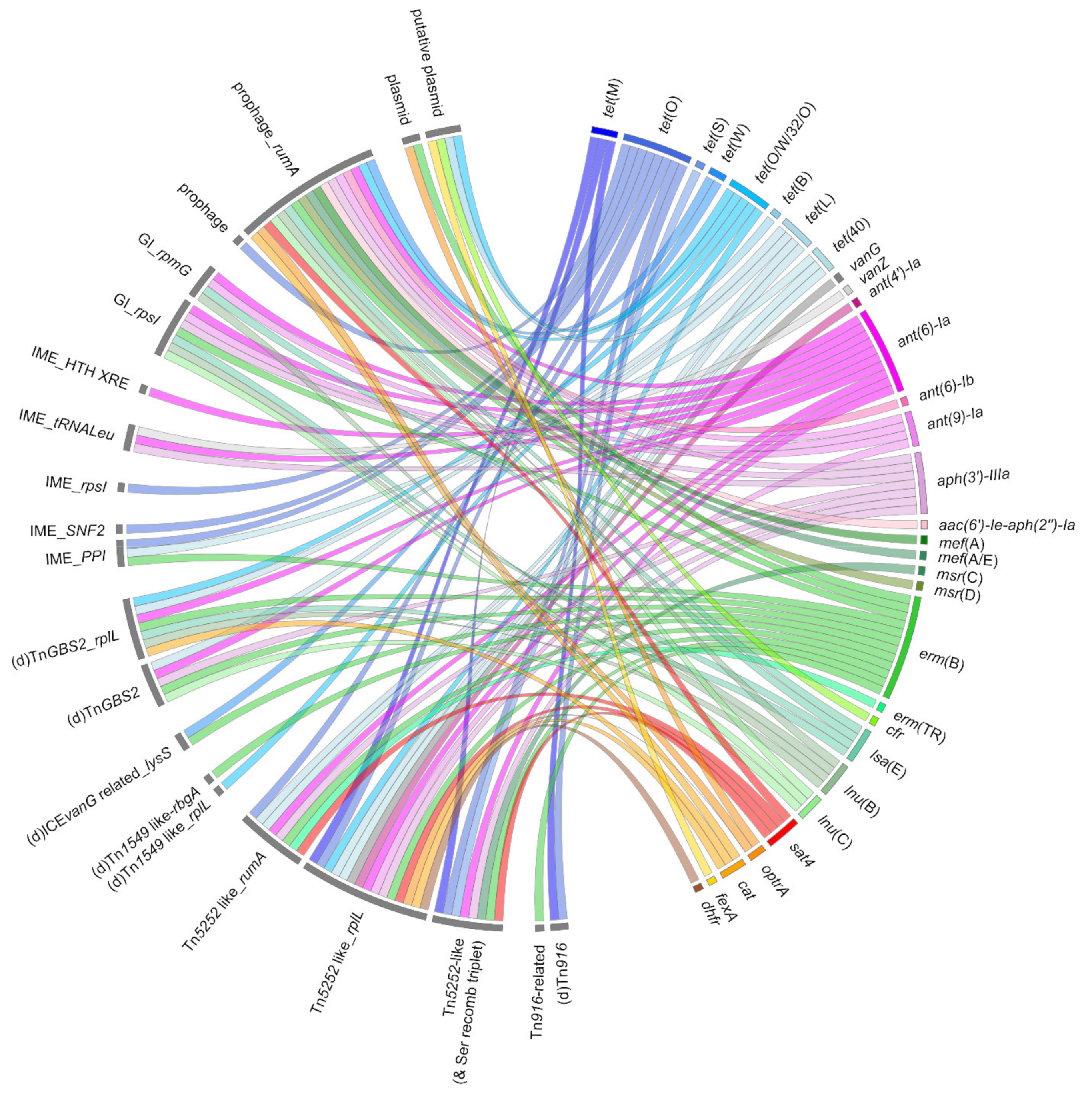
4. Dissemination of AMR Genes through Horizontal Gene Transfer in S. suis
4.1. Horizontal Gene Transfer by Transformation in S. suis
4.2. Horizontal Gene Transfer by Conjugation in S. suis
4.3. Horizontal Gene Transfer by Transduction in S. suis
5. Formation of Biofilm and Antimicrobial Resistance
6. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Goyette-Desjardins, G.; Auger, J.-P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent—An update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microbes Infect. 2014, 3, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Aragon, V.; Brockmeier, S.L.; Gebhart, C.; De Greeff, A.; Kerdsin, A.; O’Dea, M.A.; Okura, M.; Saléry, M.; Schultsz, C.; et al. Update on Streptococcus suis Research and Prevention in the Era of Antimicrobial Restriction: 4th International Workshop on S. suis. Pathogens 2020, 9, 374. [Google Scholar] [CrossRef] [PubMed]
- Staats, J.; Feder, I.; Okwumabua, O.; Chengappa, M. Streptococcus suis: Past and Present. Vet. Res. Commun. 1997, 21, 381–407. [Google Scholar] [CrossRef]
- Gottschalk, M. Streptococcosis. In Diseases of Swine, 10th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Eds.; Wiley-Blackwell Publishing: Ames, IA, USA, 2012; p. 841. Available online: https://www.academia.edu/24526077/Diseases_of_Swine_10th_edition (accessed on 12 July 2021).
- Huong, V.T.L.; Ha, N.; Huy, N.T.; Horby, P.; Nghia, H.D.T.; Thiem, V.D.; Zhu, X.; Hoa, N.T.; Hien, T.T.; Zamora, J.; et al. Epidemiology, Clinical Manifestations, and Outcomes of Streptococcus suis Infection in Humans. Emerg. Infect. Dis. 2014, 20, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, T.; Hudemann, C.; Hossain, H.M.; Hewer, A.; Tello, K.; Bandorski, D.; Rohde, M.; Valentin-Weigand, P.; Baums, C.G. Characterization of Five Zoonotic Streptococcus suis Strains from Germany, Including One Isolate from a Recent Fatal Case of Streptococcal Toxic Shock-Like Syndrome in a Hunter. J. Clin. Microbiol. 2015, 53, 3912–3915. [Google Scholar] [CrossRef] [Green Version]
- López-Mestanza, C.; Bolaño-Navarro, A.; Sánchez-Sánchez, A.; Aldea-Mansilla, C. Primer caso de shock séptico por Streptococcus suis de posible etiología alimentaria en España. Med. Intensiv. 2016, 40, 516–518. [Google Scholar] [CrossRef]
- Dutkiewicz, J.; Sroka, J.; Zając, V.; Wasiński, B.; Cisak, E.; Sawczyn-Domańska, A.; Kloc, A.; Wójcik-Fatla, A. Streptococcus suis: A re-emerging pathogen associated with occupational exposure to pigs or pork products. Part I—Epidemiology. Ann. Agric. Environ. Med. 2017, 24, 683–695. [Google Scholar] [CrossRef]
- Vötsch, D.; Willenborg, M.; Weldearegay, Y.B.; Valentin-Weigand, P. Streptococcus suis—The “Two Faces” of a Pathobiont in the Porcine Respiratory Tract. Front. Microbiol. 2018, 9, 480. [Google Scholar] [CrossRef]
- Marois-Créhan, C.; Segalés, J.; Holtkamp, D.; Chae, C.-H.; Deblanc, C.; Opriessnig, T.; Fablet, C. Interactions of Mycoplasma hyopneumoniae with other pathogens and economic impact. In Mycoplasmas in Swine; CABI Publishing: Wallingford, UK, 2021; pp. 127–145. [Google Scholar]
- Zeineldin, M.; Aldridge, B.; Lowe, J. Antimicrobial Effects on Swine Gastrointestinal Microbiota and Their Accompanying Antibiotic Resistome. Front. Microbiol. 2019, 10, 1035. [Google Scholar] [CrossRef]
- Pan, Z.; Ma, J.; Dong, W.; Song, W.; Wang, K.; Lu, C.; Yao, H. Novel Variant Serotype of Streptococcus suis Isolated from Piglets with Meningitis. Appl. Environ. Microbiol. 2014, 81, 976–985. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Liu, X.; Chen, H.; Chen, L.; Gao, X.; Pan, Z.; Wang, J.; Lu, C.; Yao, H.; Wang, L.; et al. Identification of six novel capsular polysaccharide loci (NCL) from Streptococcus suis multidrug resistant non-typeable strains and the pathogenic characteristic of strains carrying new NCL s. Transbound. Emerg. Dis. 2019, 66, 995–1003. [Google Scholar] [CrossRef]
- Weinert, L.A.; BRaDP1T Consortium; Chaudhuri, R.; Wang, J.; Peters, S.E.; Corander, J.; Jombart, T.; Baig, A.; Howell, K.J.; Vehkala, M.; et al. Genomic signatures of human and animal disease in the zoonotic pathogen Streptococcus suis. Nat. Commun. 2015, 6, 6740. [Google Scholar] [CrossRef] [Green Version]
- Estrada, A.A.; Gottschalk, M.; Rossow, S.; Rendahl, A.; Gebhart, C.; Marthaler, D.G. Serotype and Genotype (Multilocus Sequence Type) of Streptococcus suis Isolates from the United States Serve as Predictors of Pathotype. J. Clin. Microbiol. 2019, 57, e00377-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuong, N.V.; Padungtod, P.; Thwaites, G.; Carrique-Mas, J.J. Antimicrobial Usage in Animal Production: A Review of the Literature with a Focus on Low- and Middle-Income Countries. Antibiotics 2018, 7, 75. [Google Scholar] [CrossRef] [Green Version]
- Lekagul, A.; Tangcharoensathien, V.; Yeung, S. Patterns of antibiotic use in global pig production: A systematic review. Vet. Anim. Sci. 2019, 7, 100058. [Google Scholar] [CrossRef] [PubMed]
- Haenni, M.; Lupo, A.; Madec, J.-Y. Antimicrobial Resistance in Streptococcus spp. Microbiol. Spectr. 2018, 6, e0008-2017. [Google Scholar] [CrossRef]
- Varela, N.P.; Gadbois, P.; Thibault, C.; Gottschalk, M.; Dick, P.; Wilson, J. Antimicrobial resistance and prudent drug use for Streptococcus suis. Anim. Health Res. Rev. 2013, 14, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Yang, M.; Zhang, A.; Wu, J.; Chen, B.; Hua, Y.; Yu, J.; Chen, H.; Xiao, J.; Jin, M. Comparative Genomics Study of Multi-Drug-Resistance Mechanisms in the Antibiotic-Resistant Streptococcus suis R61 Strain. PLoS ONE 2011, 6, e24988. [Google Scholar] [CrossRef] [Green Version]
- Petrocchi-Rilo, M.; Martínez-Martínez, S.; Aguarón-Turrientes, Á.; Roca-Martínez, E.; García-Iglesias, M.-J.; Pérez-Fernández, E.; González-Fernández, A.; Herencia-Lagunar, E.; Gutiérrez-Martín, C.-B. Anatomical Site, Typing, Virulence Gene Profiling, Antimicrobial Susceptibility and Resistance Genes of Streptococcus suis Isolates Recovered from Pigs in Spain. Antibiotics 2021, 10, 707. [Google Scholar] [CrossRef]
- WHO. Advisory Group on Integrated Surveillance of Antimicrobial Resistance, World Health Organization, Critically Important Antimicrobials for Human Medicine: Ranking of Antimicrobial Agents for Risk Management of Antimicrobial Resistance due to Non-Human Use. Available online: http://apps.who.int/iris/bitstream/10665/255027/1/9789241512220-eng.pdf (accessed on 12 July 2021).
- Chen, L.; Song, Y.; Wei, Z.; He, H.; Zhang, A.; Jin, M. Antimicrobial Susceptibility, Tetracycline and Erythromycin Resistance Genes, and Multilocus Sequence Typing of Streptococcus suis Isolates from Diseased Pigs in China. J. Vet. Med. Sci. 2013, 75, 583–587. [Google Scholar] [CrossRef] [Green Version]
- Gurung, M.; Tamang, M.D.; Moon, D.C.; Kim, S.-R.; Jeong, J.-H.; Jang, G.-C.; Jung, S.-C.; Park, Y.-H.; Lim, S.-K. Molecular Basis of Resistance to Selected Antimicrobial Agents in the Emerging Zoonotic Pathogen Streptococcus suis. J. Clin. Microbiol. 2015, 53, 2332–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Eliang, Y.; Eguo, D.; Eshang, K.; Ege, L.; Ekashif, J.; Ewang, L. Comparative Genomic Analysis of the ICESa2603 Family ICEs and Spread of erm(B)- and tet(O)-Carrying Transferable 89K-Subtype ICEs in Swine and Bovine Isolates in China. Front. Microbiol. 2016, 7, 55. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Ma, J.; Shang, K.; Hu, X.; Liang, Y.; Li, D.; Wu, Z.; Dai, L.; Chen, L.; Wang, L. Evolution and Diversity of the Antimicrobial Resistance Associated Mobilome in Streptococcus suis: A Probable Mobile Genetic Elements Reservoir for Other Streptococci. Front. Cell. Infect. Microbiol. 2016, 6, 118. [Google Scholar] [CrossRef] [Green Version]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Markley, J.; Wencewicz, T.A. Tetracycline-Inactivating Enzymes. Front. Microbiol. 2018, 9, 1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Chen, L.; Li, D.; Wang, M.; Du, F.; Gao, Y.; Wu, Z.; Wang, L. Emergence of a vanG-carrying and multidrug resistant ICE in zoonotic pathogen Streptococccus suis. Vet. Microbiol. 2018, 222, 109–113. [Google Scholar] [CrossRef]
- Libante, V.; Nombre, Y.; Coluzzi, C.; Staub, J.; Guédon, G.; Gottschalk, M.; Teatero, S.; Fittipaldi, N.; Leblond-Bourget, N.; Payot, S. Chromosomal Conjugative and Mobilizable Elements in Streptococcus suis: Major Actors in the Spreading of Antimicrobial Resistance and Bacteriocin Synthesis Genes. Pathogens 2019, 9, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.; Tan, J.; Zeng, Y.; Li, H.; Yang, Q.; Zhou, R. Antimicrobial resistance phenotypes and genotypes of Streptococcus suis isolated from clinically healthy pigs from 2017 to 2019 in Jiangxi Province, China. J. Appl. Microbiol. 2021, 130, 797–806. [Google Scholar] [CrossRef]
- Hoa, N.T.; Chieu, T.T.B.; Nghia, H.D.T.; Mai, N.T.H.; Anh, P.H.; Wolbers, M.; Baker, S.; Campbell, J.I.; Chau, N.V.V.; Hien, T.T.; et al. The antimicrobial resistance patterns and associated determinants in Streptococcus suis isolated from humans in southern Vietnam, 1997–2008. BMC Infect. Dis. 2011, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, C.; Varaldo, P.E.; Facinelli, B. Streptococcus suis, an Emerging Drug-Resistant Animal and Human Pathogen. Front. Microbiol. 2011, 2, 235. [Google Scholar] [CrossRef] [Green Version]
- Gajic, I.; Mijac, V.; Opavski, N.; Stanojevic, M.; Lazarevic, I.; Smitran, A.; Hadnadjev, M.; Ranin, L. Distribution of macrolide-resistant genes among isolates of macrolide resistant Streptococcus pyogenes and Streptococcus pneumoniae in Serbia. Arch. Biol. Sci. 2014, 66, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Bojarska, A.; Molska, E.; Janas, K.; Skoczynska, A.; Stefaniuk, E.; Hryniewicz, W.; Sadowy, E. Streptococcus suis in invasive human infections in Poland: Clonality and determinants of virulence and antimicrobial resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 917–925. [Google Scholar] [CrossRef] [Green Version]
- Holmer, I.; Salomonsen, C.M.; Jorsal, S.E.; Astrup, L.B.; Jensen, V.F.; Høg, B.B.; Pedersen, K. Antibiotic resistance in porcine pathogenic bacteria and relation to antibiotic usage. BMC Vet. Res. 2019, 15, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, D.; Song, L.; Liu, Y.; He, T.; Liu, H.; Wu, C.; Schwarz, S.; Shen, J. First Report of the Multiresistance Gene cfr in Streptococcus suis. Antimicrob. Agents Chemother. 2013, 57, 4061–4063. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Wang, Y.; Schwarz, S. Presence and dissemination of the multiresistance gene cfr in Gram-positive and Gram-negative bacteria. J. Antimicrob. Chemother. 2013, 68, 1697–1706. [Google Scholar] [CrossRef]
- Bozdogan, B.; Berrezouga, L.; Kuo, M.-S.; Yurek, D.A.; Farley, K.A.; Stockman, B.; Leclercq, R. A New Resistance Gene, linB, Conferring Resistance to Lincosamides by Nucleotidylation in Enterococcus faecium. Antimicrob. Agents Chemother. 1999, 43, 925–929. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.; Zhang, Q.; Song, Y.; Zhang, Z.; Zhang, A.; Xiao, J.; Jin, M. Characterization of Spectinomycin Resistance in Streptococcus suis Leads to Two Novel Insights into Drug Resistance Formation and Dissemination Mechanism. Antimicrob. Agents Chemother. 2016, 60, 6390–6392. [Google Scholar] [CrossRef] [Green Version]
- Holden, M.T.G.; Hauser, H.; Sanders, M.; Ngo, T.H.; Cherevach, I.; Cronin, A.; Goodhead, I.; Mungall, K.; Quail, M.A.; Price, C.; et al. Rapid Evolution of Virulence and Drug Resistance in the Emerging Zoonotic Pathogen Streptococcus suis. PLoS ONE 2009, 4, e6072. [Google Scholar] [CrossRef] [Green Version]
- Du, F.; Lv, X.; Duan, D.; Wang, L.; Huang, J. Characterization of a Linezolid- and Vancomycin-Resistant Streptococcus suis Isolate That Harbors optrA and vanG Operons. Front. Microbiol. 2019, 10, 2026. [Google Scholar] [CrossRef]
- Yongkiettrakul, S.; Maneerat, K.; Arechanajan, B.; Malila, Y.; Srimanote, P.; Gottschalk, M.; Visessanguan, W. Antimicrobial susceptibility of Streptococcus suis isolated from diseased pigs, asymptomatic pigs, and human patients in Thailand. BMC Vet. Res. 2019, 15, 1–12. [Google Scholar] [CrossRef] [Green Version]
- O’Dea, M.A.; Laird, T.; Abraham, R.; Jordan, D.; Lugsomya, K.; Fitt, L.; Gottschalk, M.; Truswell, A.; Abraham, S. Examination of Australian Streptococcus suis isolates from clinically affected pigs in a global context and the genomic characterisation of ST1 as a predictor of virulence. Vet. Microbiol. 2018, 226, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Ku, X.; Yu, X.; Sun, Q.; Wu, H.; Chen, F.; Zhang, X.; Guo, L.; Tang, X.; He, Q. Prevalence and antimicrobial susceptibilities of bacterial pathogens in Chinese pig farms from 2013 to 2017. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sauvage, E.; Kerff, F.; Terrak, M.; Ayala, J.; Charlier, P. The penicillin-binding proteins: Structure and role in peptidoglycan biosynthesis. FEMS Microbiol. Rev. 2008, 32, 234–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, Y.; Wu, J.; Xia, Y.; Yang, M.; Xiao, J.; Yu, J. Molecular Dynamics Simulation of the Complex PBP-2x with Drug Cefuroxime to Explore the Drug Resistance Mechanism of Streptococcus suis R61. PLoS ONE 2012, 7, e35941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escudero, J.A.; Millan, A.S.; Catalan, A.; de la Campa, A.; Rivero, E.; Lopez, G.; Dominguez, L.; Moreno, M.; Gonzalez-Zorn, B. First Characterization of Fluoroquinolone Resistance in Streptococcus suis. Antimicrob. Agents Chemother. 2007, 51, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Hooper, D.C.; Jacoby, G.A. Mechanisms of drug resistance: Quinolone resistance. Ann. N. Y. Acad. Sci. 2015, 1354, 12–31. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Garcia, J.; Wang, J.; Restif, O.; Holmes, M.A.; Mather, A.E.; Weinert, L.A.; Wileman, T.M.; Thomson, J.R.; Langford, P.R.; Wren, B.W.; et al. Patterns of antimicrobial resistance in Streptococcus suis isolates from pigs with or without streptococcal disease in England between 2009 and 2014. Vet. Microbiol. 2017, 207, 117–124. [Google Scholar] [CrossRef]
- Ostrer, L.; Khodursky, R.F.; Johnson, J.R.; Hiasa, H.; Khodursky, A. Analysis of Mutational Patterns in Quinolone Resistance-Determining Regions of gyrA and parC of Clinical Isolates. Int. J. Antimicrob. Agents 2019, 53, 318–324. [Google Scholar] [CrossRef]
- Escudero, J.A.; San Millan, A.; Gutierrez, B.; Hidalgo, L.; La Ragione, R.M.; AbuOun, M.; Galimand, M.; Ferrándiz, M.J.; Domínguez, L.; de la Campa, A.G.; et al. Fluoroquinolone Efflux in Streptococcus suis Is Mediated by satAB and Not by smrA. Antimicrob. Agents Chemother. 2011, 55, 5850–5860. [Google Scholar] [CrossRef] [Green Version]
- Lai, L.; Dai, J.; Tang, H.; Zhang, S.; Wu, C.; Qiu, W.; Lu, C.; Yao, H.; Fan, H.; Wu, Z. Streptococcus suis Serotype 9 Strain GZ0565 Contains a Type VII Secretion System Putative Substrate EsxA That Contributes to Bacterial Virulence and a vanZ- like Gene That Confers Resistance to Teicoplanin and Dalbavancin in Streptococcus agalactiae. Vet. Microbiol. 2017, 205, 26–33. [Google Scholar] [CrossRef]
- El Garch, F.; de Jong, A.; Simjee, S.; Moyaert, H.; Klein, U.; Ludwig, C.; Marion, H.; Haag-Diergarten, S.; Richard-Mazet, A.; Thomas, V.; et al. Monitoring of Antimicrobial Susceptibility of Respiratory Tract Pathogens Isolated from Diseased Cattle and Pigs across Europe, 2009–2012: VetPath Results. Vet. Microbiol. 2016, 194, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Dvorak, C.M.T.; Estrada, A.A.; Gebhart, C.; Marthaler, D.G.; Murtaugh, M.P. MinION Sequencing of Streptococcus suis Allows for Functional Characterization of Bacteria by Multilocus Sequence Typing and Antimicrobial Resistance Profiling. J. Microbiol. Methods 2020, 169, 105817. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, T.; Oshima, M.; Yamagishi, J.; Muramatsu, C.; Asai, T. Changes in Antimicrobial Resistance Phenotypes and Genotypes in Streptococcus suis Strains Isolated from Pigs in the Tokai Area of Japan. J. Vet. Med. Sci. 2020, 82, 9–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Y.; Li, D.; Hao, W.; Schwarz, S.; Shan, X.; Liu, B.; Zhang, S.-M.; Li, X.-S.; Du, X.-D. A Prophage and Two ICESa2603-Family Integrative and Conjugative Elements (ICEs) Carrying optrA in Streptococcus suis. J. Antimicrob. Chemother. 2019, 74, 2876–2879. [Google Scholar] [CrossRef]
- Kehrenberg, C.; Schwarz, S. fexA, a Novel Staphylococcus lentus Gene Encoding Resistance to Florfenicol and Chloramphenicol. Antimicrob. Agents Chemother. 2004, 48, 615–618. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Chen, L.; Wu, Z.; Wang, L. Retrospective Analysis of Genome Sequences Revealed the Wide Dissemination of optrA in Gram-Positive Bacteria. J. Antimicrob. Chemother. 2017, 72, 614–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Liu, J.; Zhang, Y.; Wang, D.; Liu, R.; Liu, G.; Yao, H.; Pan, Z. Bacitracin Resistance and Enhanced Virulence of Streptococcus suis via a Novel Efflux Pump. BMC Vet. Res. 2019, 15, 377. [Google Scholar] [CrossRef]
- Huang, J.; Sun, J.; Wu, Y.; Chen, L.; Duan, D.; Lv, X.; Wang, L. Identification and Pathogenicity of an XDR Streptococcus suis Isolate That Harbours the Phenicol-Oxazolidinone Resistance Genes optrA and cfr, and the Bacitracin Resistance Locus bcrABDR. Int. J. Antimicrob. Agents 2019, 54, 43–48. [Google Scholar] [CrossRef]
- Arber, W. Horizontal Gene Transfer among Bacteria and Its Role in Biological Evolution. Life 2014, 4, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Soucy, S.M.; Huang, J.; Gogarten, J.P. Horizontal Gene Transfer: Building the Web of Life. Nat. Rev. Genet. 2015, 16, 472–482. [Google Scholar] [CrossRef]
- Soler, N.; Forterre, P. Vesiduction: The Fourth Way of HGT. Environ. Microbiol. 2020, 22, 2457–2460. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, L.; Wahl, A.; Fléchard, M.; Mignolet, J.; Hols, P. Regulation of Competence for Natural Transformation in Streptococci. Infect. Genet. Evol. 2014, 33, 343–360. [Google Scholar] [CrossRef] [PubMed]
- Shanker, E.; Morrison, D.A.; Talagas, A.; Nessler, S.; Federle, M.J.; Prehna, G. Pheromone Recognition and Selectivity by ComR Proteins among Streptococcus Species. PLoS Pathog. 2016, 12, e1005979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaccaria, E.; van Baarlen, P.; de Greeff, A.; Morrison, D.A.; Smith, H.; Wells, J.M. Control of Competence for DNA Transformation in Streptococcus suis by Genetically Transferable Pherotypes. PLoS ONE 2014, 9, e99394. [Google Scholar] [CrossRef] [Green Version]
- Okura, M.; Nozawa, T.; Watanabe, T.; Murase, K.; Nakagawa, I.; Takamatsu, D.; Osaki, M.; Sekizaki, T.; Gottschalk, M.; Hamada, S.; et al. A Locus Encoding Variable Defense Systems against Invading DNA Identified in Streptococcus suis. Genome Biol. Evol. 2017, 9, 1000–1012. [Google Scholar] [CrossRef]
- Zaccaria, E.; Wels, M.; van Baarlen, P.; Wells, J.M. Temporal Regulation of the Transformasome and Competence Development in Streptococcus suis. Front. Microbiol. 2016, 7, 1922. [Google Scholar] [CrossRef]
- Zhu, Y.; Ma, J.; Zhang, Y.; Zhong, X.; Bai, Q.; Dong, W.; Pan, Z.; Liu, G.; Zhang, C.; Yao, H. CrfP, a Fratricide Protein, Contributes to Natural Transformation in Streptococcus suis. Vet. Res. 2021, 52, 50. [Google Scholar] [CrossRef]
- Bjedov, I.; Tenaillon, O.; Gérard, B.; Souza, V.; Denamur, E.; Radman, M.; Taddei, F.; Matic, I. Stress-Induced Mutagenesis in Bacteria. Science 2003, 300, 1404. [Google Scholar] [CrossRef]
- Claverys, J.-P.; Prudhomme, M.; Martin, B. Induction of Competence Regulons as a General Response to Stress in Gram-Positive Bacteria. Annu. Rev. Microbiol. 2006, 60, 451–475. [Google Scholar] [CrossRef]
- Ferrando, M.L.; Gussak, A.; Mentink, S.; Gutierrez, M.F.; van Baarlen, P.; Wells, J.M. Active Human and Porcine Serum Induce Competence for Genetic Transformation in the Emerging Zoonotic Pathogen Streptococcus suis. Pathogens 2021, 10, 156. [Google Scholar] [CrossRef] [PubMed]
- Bellanger, X.; Payot, S.; Leblond-Bourget, N.; Guédon, G. Conjugative and Mobilizable Genomic Islands in Bacteria: Evolution and Diversity. FEMS Microbiol. Rev. 2014, 38, 720–760. [Google Scholar] [CrossRef] [Green Version]
- Guédon, G.; Libante, V.; Coluzzi, C.; Payot, S.; Leblond-Bourget, N. The Obscure World of Integrative and Mobilizable Elements, Highly Widespread Elements That Pirate Bacterial Conjugative Systems. Genes 2017, 8, 337. [Google Scholar] [CrossRef] [Green Version]
- Ambroset, C.; Coluzzi, C.; Guédon, G.; Devignes, M.-D.; Loux, V.; Lacroix, T.; Payot, S.; Leblond-Bourget, N. New Insights into the Classification and Integration Specificity of Streptococcus Integrative Conjugative Elements through Extensive Genome Exploration. Front. Microbiol. 2016, 6, 1483. [Google Scholar] [CrossRef] [Green Version]
- Yi, L.; Jin, M.; Li, J.; Grenier, D.; Wang, Y. Antibiotic Resistance Related to Biofilm Formation in Streptococcus suis. Appl. Microbiol. Biotechnol. 2020, 104, 8649–8660. [Google Scholar] [CrossRef]
- Palmieri, C.; Mingoia, M.; Varaldo, P.E. Unconventional Circularizable Bacterial Genetic Structures Carrying Antibiotic Resistance Determinants. Antimicrob. Agents Chemother. 2013, 57, 2440–2441. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, C.; Magi, G.; Mingoia, M.; Bagnarelli, P.; Ripa, S.; Varaldo, P.E.; Facinelli, B. Characterization of a Streptococcus suis tet(O/W/32/O)-Carrying Element Transferable to Major Streptococcal Pathogens. Antimicrob. Agents Chemother. 2012, 56, 4697–4702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chander, Y.; Oliveira, S.R.; Goyal, S.M. Identification of the tet(B) Resistance Gene in Streptococcus suis. Vet. J. 2011, 189, 359–360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, P.; Wang, Y.; Fu, L.; Liu, L.; Xu, D.; Hou, Y.; Li, Y.; Fu, M.; Wang, X.; et al. Capsular Serotypes, Antimicrobial Susceptibility, and the Presence of Transferable Oxazolidinone Resistance Genes in Streptococcus suis Isolated from Healthy Pigs in China. Vet. Microbiol. 2020, 247, 108750. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.L.; Lu, C.P. Isolation and Identification of a Bacteriophage Capable of Infecting Streptococcus suis Type 2 Strains. Vet. Microbiol. 2008, 132, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Klausen, M.; Gjermansen, M.; Kreft, J.-U.; Tolker-Nielsen, T. Dynamics of Development and Dispersal in Sessile Microbial Communities: Examples from Pseudomonas aeruginosa and Pseudomonas putida Model Biofilms. FEMS Microbiol. Lett. 2006, 261, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial Biofilms: A Common Cause of Persistent Infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grenier, D.; Grignon, L.; Gottschalk, M. Characterisation of Biofilm Formation by a Streptococcus suis Meningitis Isolate. Vet. J. 2009, 179, 292–295. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Y.; Sun, L.; Grenier, D.; Yi, L. Streptococcus suis Biofilm: Regulation, Drug-Resistance Mechanisms, and Disinfection Strategies. Appl. Microbiol. Biotechnol. 2018, 102, 9121–9129. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, B.; Li, J.; Gong, S.; Dong, X.; Mao, C.; Yi, L. LuxS/AI-2 System Is Involved in Fluoroquinolones Susceptibility in Streptococcus suis through Overexpression of Efflux Pump SatAB. Vet. Microbiol. 2019, 233, 154–158. [Google Scholar] [CrossRef]
- Wang, Y.; Gong, S.; Dong, X.; Li, J.; Grenier, D.; Yi, L. In Vitro Mixed Biofilm of Streptococcus suis and Actinobacillus pleuropneumoniae Impacts Antibiotic Susceptibility and Modulates Virulence Factor Gene Expression. Front. Microbiol. 2020, 11, 507. [Google Scholar] [CrossRef]
- Liu, B.; Yi, L.; Li, J.; Wang, Y.; Mao, C.; Wang, Y. Autoinducer-2 Influences Tetracycline Resistance in Streptococcus suis by Regulating the tet(M) Gene via Transposon Tn916. Res. Vet. Sci. 2020, 128, 269–274. [Google Scholar] [CrossRef]
- Waack, U.; Nicholson, T.L. Subinhibitory Concentrations of Amoxicillin, Lincomycin, and Oxytetracycline Commonly Used to Treat Swine Increase Streptococcus suis Biofilm Formation. Front. Microbiol. 2018, 9, 2707. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dechêne-Tempier, M.; Marois-Créhan, C.; Libante, V.; Jouy, E.; Leblond-Bourget, N.; Payot, S. Update on the Mechanisms of Antibiotic Resistance and the Mobile Resistome in the Emerging Zoonotic Pathogen Streptococcus suis. Microorganisms 2021, 9, 1765. https://doi.org/10.3390/microorganisms9081765
Dechêne-Tempier M, Marois-Créhan C, Libante V, Jouy E, Leblond-Bourget N, Payot S. Update on the Mechanisms of Antibiotic Resistance and the Mobile Resistome in the Emerging Zoonotic Pathogen Streptococcus suis. Microorganisms. 2021; 9(8):1765. https://doi.org/10.3390/microorganisms9081765
Chicago/Turabian StyleDechêne-Tempier, Manon, Corinne Marois-Créhan, Virginie Libante, Eric Jouy, Nathalie Leblond-Bourget, and Sophie Payot. 2021. "Update on the Mechanisms of Antibiotic Resistance and the Mobile Resistome in the Emerging Zoonotic Pathogen Streptococcus suis" Microorganisms 9, no. 8: 1765. https://doi.org/10.3390/microorganisms9081765
APA StyleDechêne-Tempier, M., Marois-Créhan, C., Libante, V., Jouy, E., Leblond-Bourget, N., & Payot, S. (2021). Update on the Mechanisms of Antibiotic Resistance and the Mobile Resistome in the Emerging Zoonotic Pathogen Streptococcus suis. Microorganisms, 9(8), 1765. https://doi.org/10.3390/microorganisms9081765