Dietary Betaine Improves Intestinal Barrier Function and Ameliorates the Impact of Heat Stress in Multiple Vital Organs as Measured by Evans Blue Dye in Broiler Chickens


 , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics
2.2. Animals, Diets and Experimental Design
2.3. Physiological Responses
2.4. Evans Blue Dye Injection, Slaughter and Tissue Collection
2.5. Evans Blue Dye Extraction and Qualification
2.6. Intestinal Transepithelial Electrical Resistance
2.7. High-Performance Liquid Chromatography Analysis
2.8. Morphometric Analysis
2.9. Statistics Analysis
3. Results
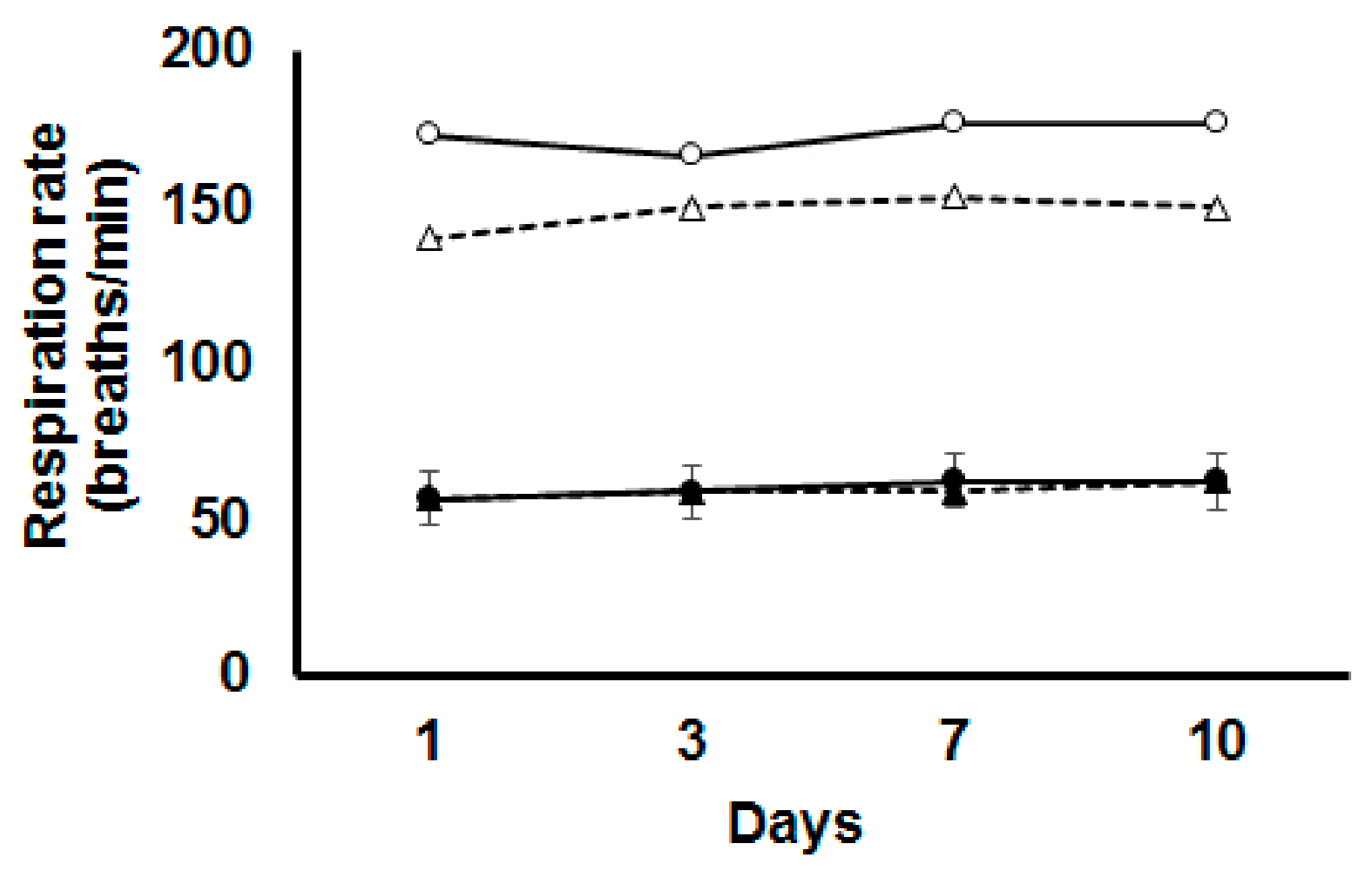
3.1. Respiration Rate
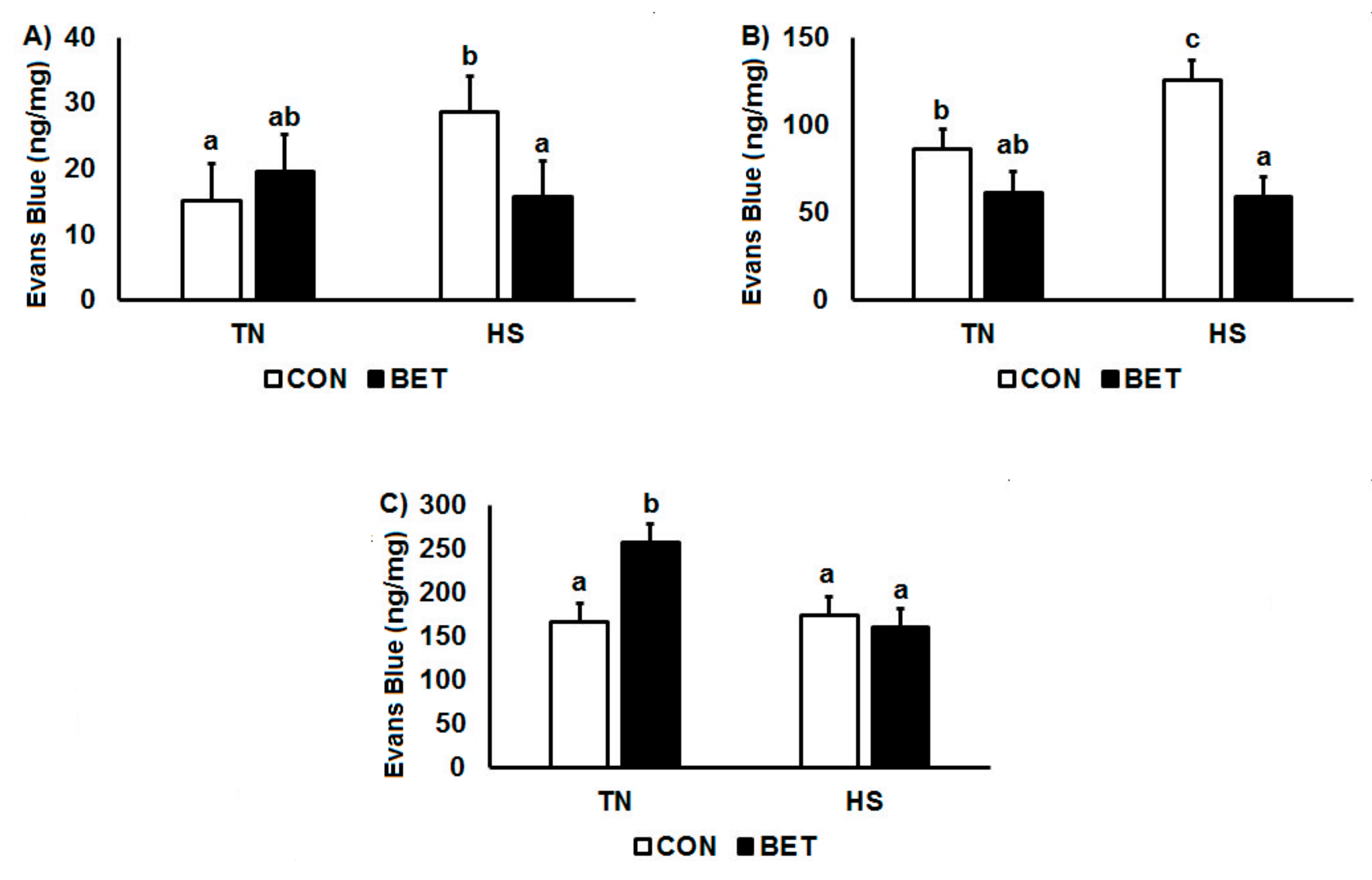
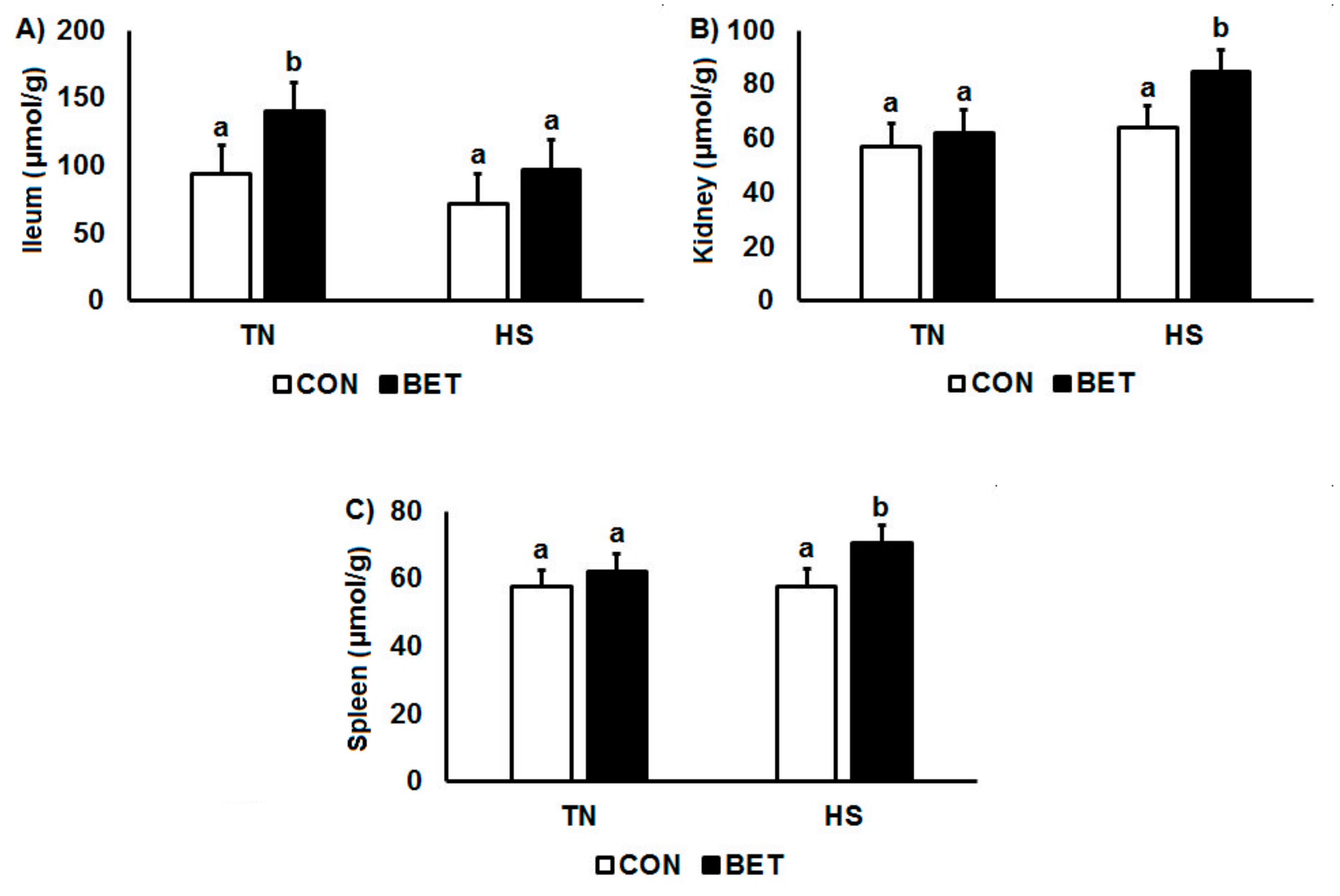
3.2. Evans Blue Dye Distribution
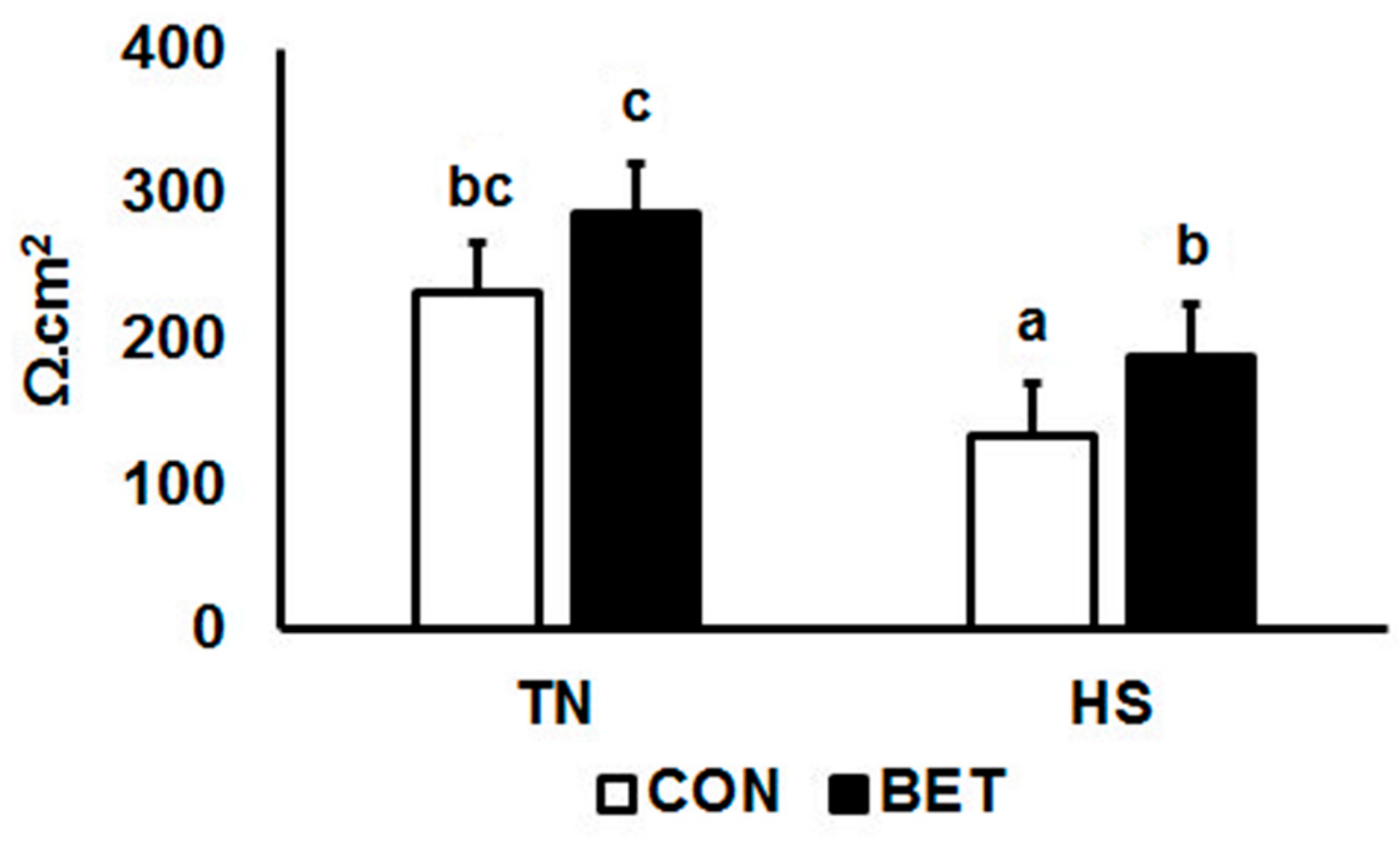
3.3. Transepithelial Electrical Resistance
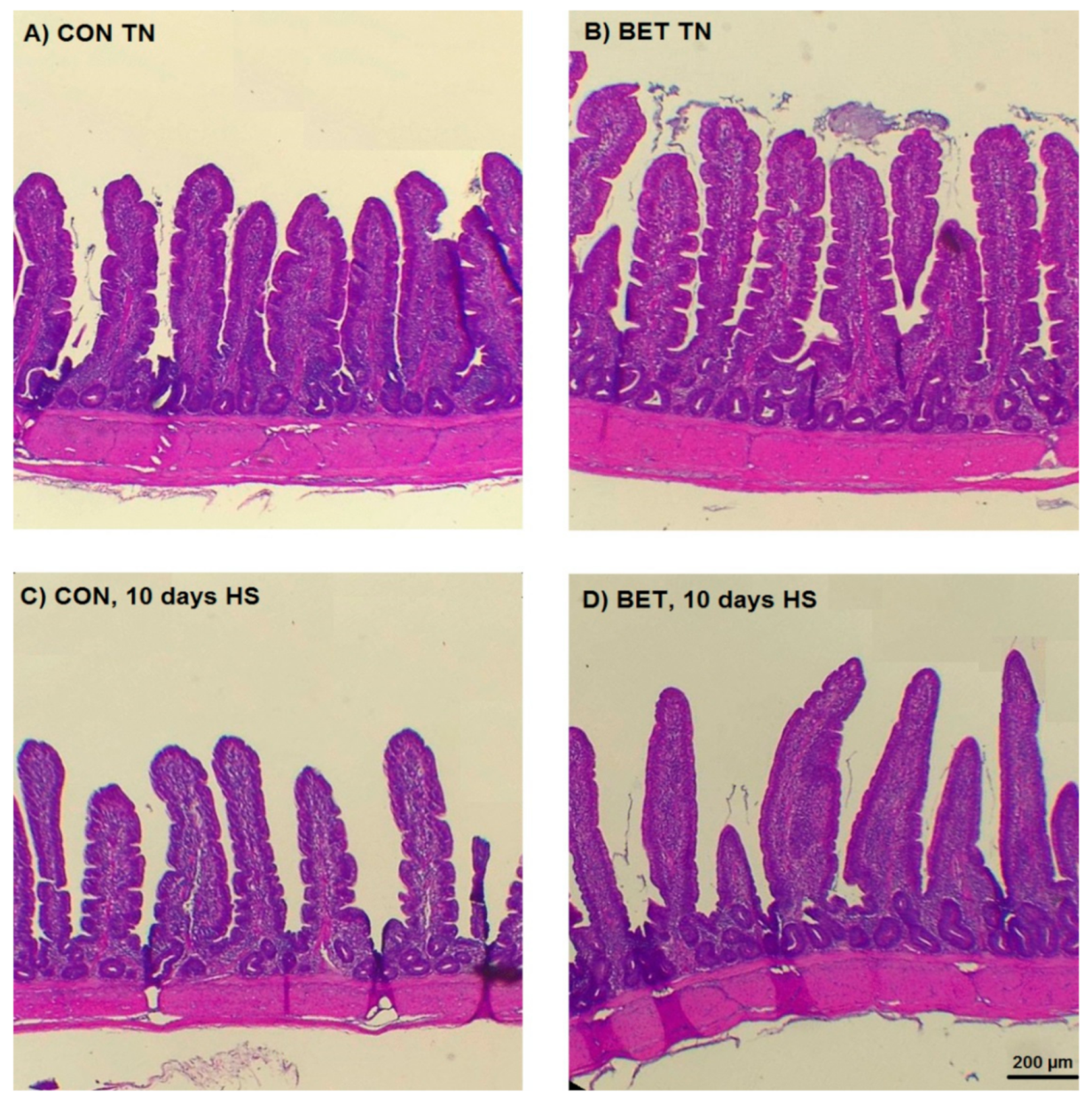
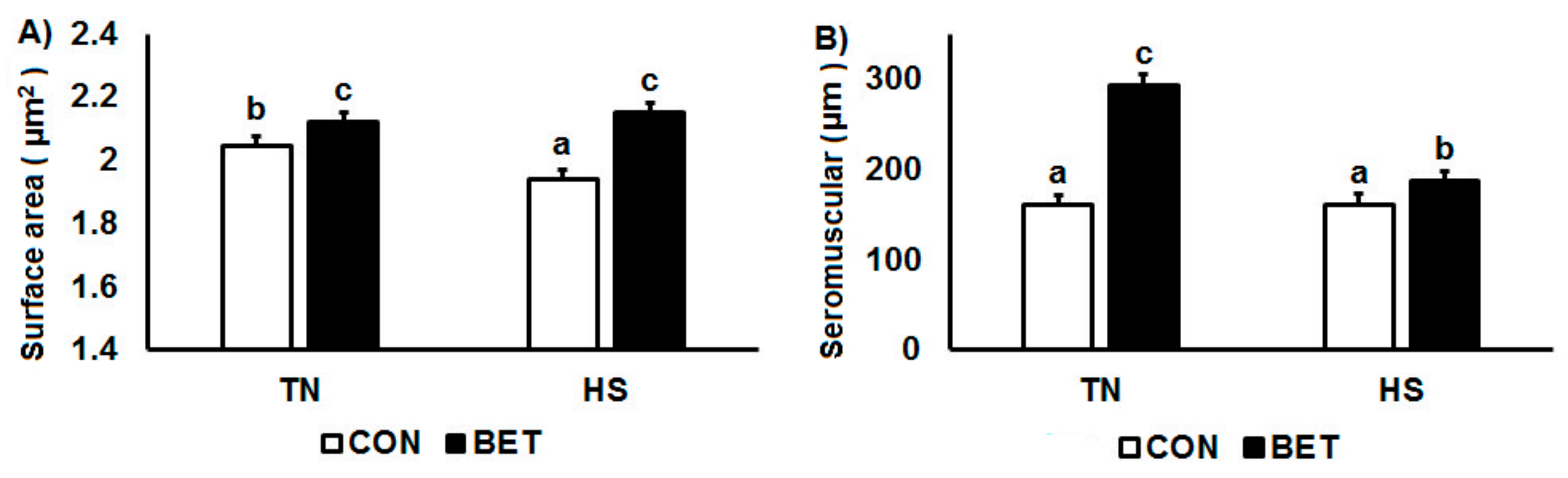
3.4. Morphometric Analysis
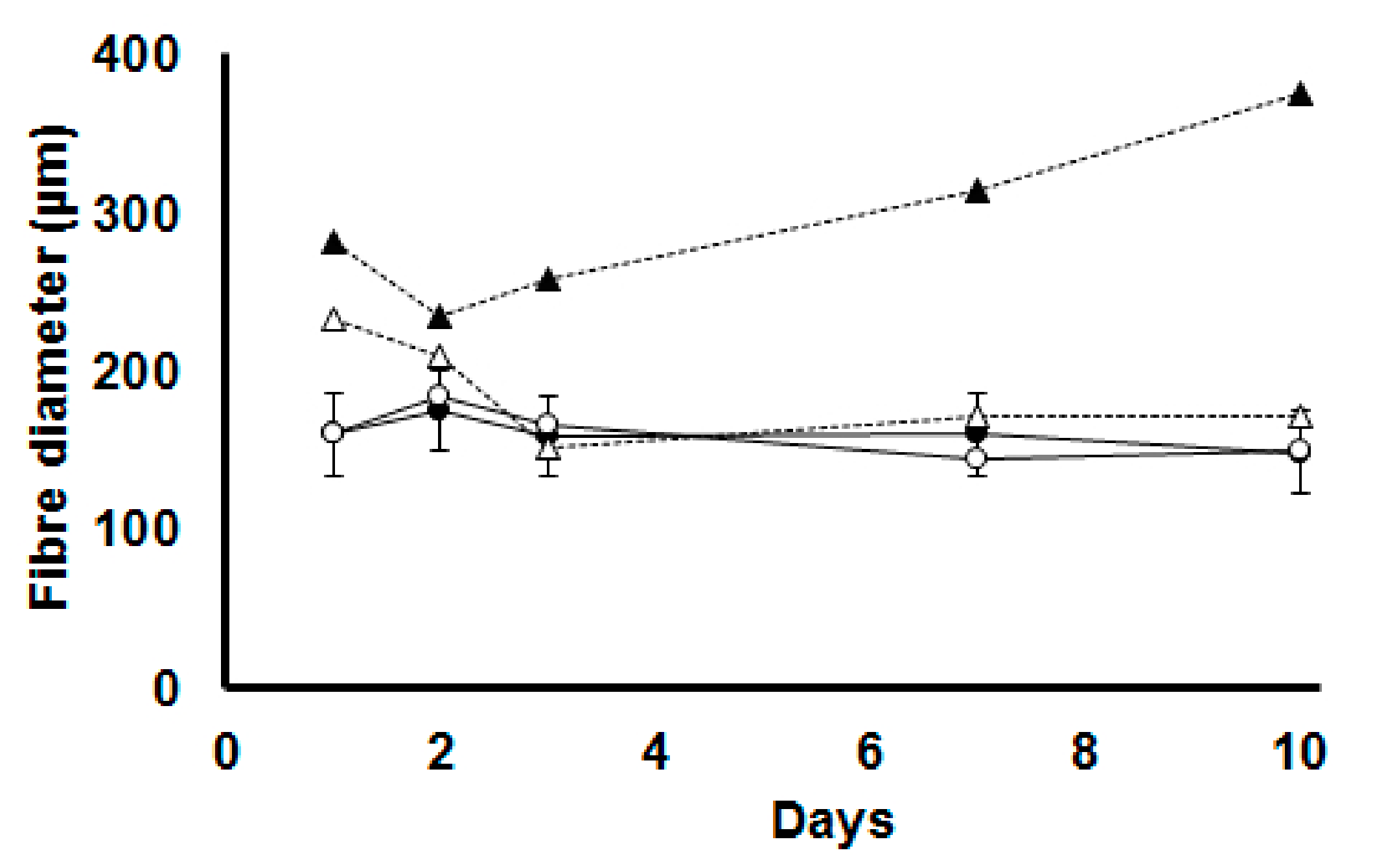
3.5. Betaine Distribution
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Renaudeau, D.; Collin, A.; Yahav, S.; de Basilio, V.; Gourdine, J.L.; Collier, R.J. Adaptation to hot climate and strategies to alleviate heat stress in livestock production. Animal 2012, 6, 707–728. [Google Scholar]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Le, H.H.; Suleria, H.A.R.; Warner, R.D.; Dunshea, F.R. Growth Performance and Characterization of Meat Quality of Broiler Chickens Supplemented with Betaine and Antioxidants under Cyclic Heat Stress. Antioxidants 2019, 8, 336. [Google Scholar]
- Tao, X.; Dong, H.; Zhang, Z.; Xin, H. Daily variation of thyroid hormones in broiler under high-temperature conditions. In Livestock Environment VII, Proceedings of the 7th International Symposium, Beijing, China, 18–20 May 2005; American Society of Agricultural and Biological Engineers: Saint Joseph, MO, USA, 2005. [Google Scholar]
- Liu, F.; Cottrell, J.J.; Furness, J.B.; Rivera, L.R.; Kelly, F.W.; Wijesiriwardana, U.; Pustovit, R.V.; Fothergill, L.J.; Bravo, D.M.; Celi, P. Selenium and vitamin E together improve intestinal epithelial barrier function and alleviate oxidative stress in heat-stressed pigs. Exp. Physiol. 2016, 101, 801–810. [Google Scholar]
- Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Ringuet, M.; Furness, J.; Dunshea, F.R. Betaine and Antioxidants Improve Growth Performance, Breast Muscle Development and Ameliorate Thermoregulatory Responses to Cyclic Heat Exposure in Broiler Chickens. Animals 2018, 8, 162. [Google Scholar]
- Goo, D.; Kim, J.H.; Park, G.H.; Reyes, D.; Badillo, J.; Kil, D.Y. Effect of Heat Stress and Stocking Density on Growth Performance, Breast Meat Quality, and Intestinal Barrier Function in Broiler Chickens. Animals 2019, 9, 107. [Google Scholar]
- Pearce, S.C.; Mani, V.; Boddicker, R.L.; Johnson, J.S.; Weber, T.E.; Ross, J.W.; Rhoads, R.P.; Baumgard, L.H.; Gabler, N.K. Heat stress reduces intestinal barrier integrity and favors intestinal glucose transport in growing pigs. PLoS ONE 2013, 8, e70215. [Google Scholar]
- Alhenaky, A.; Abdelqader, A.; Abuajamieh, M.; Al-Fataftah, A.-R. The effect of heat stress on intestinal integrity and Salmonella invasion in broiler birds. J. Therm. Biol. 2017, 70, 9–14. [Google Scholar]
- Eshel, G.M.; Safar, P.; Stezoski, W. The role of the gut in the pathogenesis of death due to hyperthermia. Am. J. Foren. Med. Path. 2001, 22, 100–104. [Google Scholar]
- Davis, B.C.; Tillman, H.; Chung, R.T.; Stravitz, R.T.; Reddy, R.; Fontana, R.J.; McGuire, B.; Davern, T.; Lee, W.M.; Group, A.L.F.S. Heat stroke leading to acute liver injury & failure: A case series from the Acute Liver Failure Study Group. Liver Int. 2017, 37, 509–513. [Google Scholar]
- Hifumi, T.; Kondo, Y.; Shimizu, K.; Miyake, Y. Heat stroke. J. Intensive Care 2018, 6, 30. [Google Scholar]
- Crooke, A.; Morris, C. The determination of plasma volume by the Evans blue method. J. Physiol. 1942, 101, 217. [Google Scholar]
- Radu, M.; Chernoff, J. An in vivo assay to test blood vessel permeability. J. Vis. Exp. 2013, 73, e580062. [Google Scholar]
- Caplan, B.A.; Schwartz, C.J. Increased endothelial cell turnover in areas of in vivo Evans Blue uptake in the pig aorta. Atherosclerosis 1973, 17, 401–417. [Google Scholar]
- Craig, S.A.S. Betaine in human nutrition. Am. J. Clin. Nutr. 2004, 80, 539–549. [Google Scholar]
- Tsai, M.T.; Chen, C.Y.; Pan, Y.H.; Wang, S.H.; Mersmann, H.J.; Ding, S.T. Alleviation of carbon-tetrachloride-induced liver injury and fibrosis by betaine supplementation in chickens. Evid. Based Complement. Altern. Med. 2015, 2015, 725379. [Google Scholar]
- Dunshea, F.R.; Leury, B.J.; Fahri, F.; DiGiacomo, K.; Hung, A.; Chauhan, S.; Clarke, I.J.; Collier, R.; Little, S.; Baumgard, L. Amelioration of thermal stress impacts in dairy cows. Anim. Prod. Sci. 2013, 53, 965–975. [Google Scholar]
- Suster, D.; Leury, B.J.; King, R.H.; Mottram, M.; Dunshea, F.R. Interrelationships between porcine somatotropin (pST), betaine, and energy level on body composition and tissue distribution of finisher boars. Aust. J. Agric. Res. 2004, 55, 983–990. [Google Scholar]
- Alirezaei, M.; Gheisari, H.R.; Ranjbar, V.R.; Hajibemani, A. Betaine: A promising antioxidant agent for enhancement of broiler meat quality. Br. Poult. Sci. 2012, 53, 699–707. [Google Scholar]
- NRC. Nutrient Requirements of Poultry; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Laryea, M.D.; Steinhagen, F.; Pawliczek, S.; Wendel, U. Simple method for the routine determination of betaine and N,N-dimethylglycine in blood and urine. Clin. Chem. 1998, 44, 1937–1941. [Google Scholar]
- Baddeley, A.; Gundersen, H.-J.G.; Cruz-Orive, L.M. Estimation of surface area from vertical sections. J. Microsc. 1986, 142, 259–276. [Google Scholar]
- Collins, T.J. ImageJ for microscopy. Biotechniques 2007, 43, S25–S30. [Google Scholar]
- Marchini, C.F.P.; Silva, P.L.; Nascimento, M.R.B.M.; Beletti, M.E.; Silva, N.M.; Guimarães, E.C. Body weight, intestinal morphometry and cell proliferation of broiler chickens submitted to cyclic heat stress. Int. J. Poult. Sci. 2011, 10, 455–460. [Google Scholar]
- Pearce, S.C.; Sanz-Fernandez, M.V.; Hollis, J.H.; Baumgard, L.H.; Gabler, N.K. Short-term exposure to heat stress attenuates appetite and intestinal integrity in growing pigs. J. Anim. Sci. 2014, 92, 5444–5454. [Google Scholar]
- Wolfenson, D.; Frei, Y.F.; Snapir, N.; Berman, A. Heat stress effects on capillary blood flow and its redistribution in the laying hen. Pflügers Arch. 1981, 390, 86–93. [Google Scholar]
- Hales, J.R.; Rowell, L.B.; King, R.B. Regional distribution of blood flow in awake heat-stressed baboons. Am. J. Physiol.-Heart Circ. Physiol. 1979, 237, H705–H712. [Google Scholar]
- Hall, D.M.; Baumgardner, K.R.; Oberley, T.D.; Gisolfi, C.V. Splanchnic tissues undergo hypoxic stress during whole body hyperthermia. Am. J. Physiol.-Gastrointest. Liver Physiol. 1999, 276, G1195–G1203. [Google Scholar]
- Lambert, G.P.; Gisolfi, C.V.; Berg, D.J.; Moseley, P.L.; Oberley, L.W.; Kregel, K.C. Selected contribution: Hyperthermia-induced intestinal permeability and the role of oxidative and nitrosative stress. J. Appl. Physiol. 2002, 92, 1750–1761. [Google Scholar]
- Lambert, G.P. Stress-induced gastrointestinal barrier dysfunction and its inflammatory effects. J. Anim. Sci. 2009, 87, E101–E108. [Google Scholar]
- Sils, I.V.; Szlyk-Modrow, P.C.; Tartarini, K.A.; Matthew, C.B.; Francesconi, R.P. Effect of nitric oxide synthase inhibition on regional blood flow during hyperthermia. J. Therm. Biol. 2001, 26, 1–7. [Google Scholar]
- Kregel, K.C.; Wall, P.T.; Gisolfi, C.V. Peripheral vascular responses to hyperthermia in the rat. J. Appl. Physiol. 1988, 64, 2582–2588. [Google Scholar]
- Glaser, J.; Lemery, J.; Rajagopalan, B.; Diaz, H.F.; García-Trabanino, R.; Taduri, G.; Madero, M.; Amarasinghe, M.; Abraham, G.; Anutrakulchai, S. Climate change and the emergent epidemic of CKD from heat stress in rural communities: The case for heat stress nephropathy. Clin. J. Am. Soc. Nephrol. 2016, 11, 1472–1483. [Google Scholar]
- Zhao, W.; Dunshea, F.R.; Zhang, Z.; Furness, J.B.; Ringuet, M.T.; DiGiacomo, K.; Leury, B.J.; Roura, E.; Wijffels, G.; Renaudeau, D.; et al. Quantification of the blood volume and pattern of organ permeability in the heat stressed pig. In Symposium on Gut Health in Production of Food Animals; The Federation of Animal Science Societies: St Louis, MO, USA, 2018. [Google Scholar]
- He, S.; Yu, Q.; He, Y.; Hu, R.; Xia, S.; He, J. Dietary resveratrol supplementation inhibits heat stress-induced high-activated innate immunity and inflammatory response in spleen of yellow-feather broilers. Poult. Sci. 2019, 98, 6378–6387. [Google Scholar]
- Trout, J.M.; Mashaly, M.M. The effects of adrenocorticotropic hormone and heat stress on the distribution of lymphocyte populations in immature male chickens. Poult. Sci. 1994, 73, 1694–1698. [Google Scholar]
- Egbuniwe, I.C.; Ayo, J.O.; Ocheja, O.B. Betaine and ascorbic acid modulate indoor behavior and some performance indicators of broiler chickens in response to hot-dry season. J. Therm. Biol. 2018, 76, 38–44. [Google Scholar]
- Kettunen, H.; Tiihonen, K.; Peuranen, S.; Saarinen, M.T.; Remus, J.C. Dietary betaine accumulates in the liver and intestinal tissue and stabilizes the intestinal epithelial structure in healthy and coccidia-infected broiler chicks. Comp. Biochem. Physiol. A 2001, 130, 759–769. [Google Scholar]
- Dos Santos, T.T.; Dassi, S.C.; Franco, C.R.C.; da Costa, C.R.V.; Lee, S.A.; da Silva, A.V.F. Influence of fibre and betaine on development of the gastrointestinal tract of broilers between hatch and 14 d of age. Anim. Nutr. 2019, 5, 163–173. [Google Scholar]
- Sakomura, N.K.; Barbosa, N.A.A.; da Silva, E.P.; Longo, F.A.; Kawauchi, I.M.; Fernandes, J.B.K. Effect of betaine supplementation in diets for broiler chickens on thermoneutral environment. Revista Brasileira de Ciências Agrárias 2013, 8, 336–341. [Google Scholar]
- Hamidi, H.; Pourreza, J.; Rahimi, H. Dietary betaine affect duodenal histology of broiler challenged with a mixed coccidial infection. Pak. J. Biol. Sci. 2009, 12, 291–295. [Google Scholar]
- Schhwahn, B.C.; Hafner, D.; Hohlfeld, T.; Balkenhol, N.; Laryea, M.D.; Wendel, U. Pharmacokinetics of oral betaine in healthy subjects and patients with homocystinuria. Br. J. Clin. Pharmacol. 2003, 55, 6–13. [Google Scholar]
- Bauchart-Thevret, C.; Stoll, B.; Chacko, S.; Burrin, D.G. Sulfur amino acid deficiency upregulates intestinal methionine cycle activity and suppresses epithelial growth in neonatal pigs. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E1239–E1250. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissues | Diet (D) | Temp (T) | Day of Thermal Challenge | SED | Significance 1 | ||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 7 | 10 | |||||
| Psoas Major (ng/mg) | CON | TN | 8.3 | 16.0 | 8.0 | 14.3 | 29.4 | 11.3 | D × T * |
| HS | 8.3 | 42.1 | 24.5 | 29.1 | 39.1 | ||||
| BET | TN | 6.7 | 30.8 | 16.4 | 18.8 | 25.2 | |||
| HS | 9.5 | 6.2 | 16.7 | 14.3 | 31.8 | ||||
| Liver (ng/mg) | CON | TN | 37 | 91 | 96 | 110 | 94 | 32.8 | |
| HS | 65 | 107 | 111 | 83 | 80 | ||||
| BET | TN | 128 | 111 | 106 | 90 | 80 | |||
| HS | 58 | 141 | 65 | 60 | 87 | ||||
| Kidney (ng/mg) | CON | TN | 96 a | 93 ab | 62 | 61 | 119 | 28.2 | T **, D ***, D × T ** |
| HS | 158 b | 117 b | 98 | 94 | 161 | D × T × Day + | |||
| BET | TN | 90 a | 31 a | 41 | 53 | 93 | |||
| HS | 62 a | 61 ab | 36 | 45 | 91 | ||||
| Spleen 2 (ng/mg) | CON | TN | 2.05 | 2.08 | 2.20 | 2.30 | 2.36 | 0.15 | T *, D × * |
| (112) | (120) | (157) | (199) | (229) | |||||
| HS | 2.04 | 1.89 | 2.31 | 2.31 | 2.41 | ||||
| (110) | (78) | (206) | (206) | (258) | |||||
| BET | TN | 2.28 | 2.19 | 2.51 | 2.43 | 2.49 | |||
| (191) | (153) | (321) | (267) | (309) | |||||
| HS | 2.04 | 2.03 | 2.36 | 2.14 | 2.22 | ||||
| (110) | (108) | (231) | (175) | (167) | |||||
| Jejunum 2 (ng/mg) | CON | TN | 2.02 | 2.29 a | 1.90 | 1.86 ab | 2.08 | 0.14 | D *, D × T × Day ** |
| (104) | (194) | (79) | (73) | (121) | |||||
| HS | 1.85 | 1.84 b | 1.95 | 2.08 b | 1.90 | ||||
| (71) | (69) | (89) | (119) | (79) | |||||
| BET | TN | 1.84 | 1.76 b | 1.86 | 1.90 ab | 1.98 | |||
| (69) | (58) | (73) | (80) | (100) | |||||
| HS | 1.99 | 2.10 ab | 1.87 | 1.69 a | 1.85 | ||||
| (97) | (125) | (75) | (49) | (70) | |||||
| Ileum (ng/mg) | CON | TN | 151 | 100 | 77 | 75 | 54 a | 34.2 | D *, D × T × Day ** |
| HS | 115 | 68 | 69 | 85 | 209 b | ||||
| BET | TN | 70 | 146 | 49 | 32 | 108 a | |||
| HS | 83 | 106 | 55 | 34 | 75 a | ||||
| Ileal | Diet (D) | Temp (T) | Day of Thermal Challenge | SED | Significance 1 | ||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 7 | 10 | |||||
| Transepithelial resistance 2 (Ω.cm2) | CON | TN | - | - | 268 a | 211 | 215 a | 35.7 | T ***, D **, Day *** |
| HS | - | - | 159 a | 148 | 93.0 b | D × T × Day + | |||
| BET | TN | - | - | 424 b | 246 | 175 ab | |||
| HS | - | - | 212 a | 192 | 161 ab | ||||
| Villous height (µm) | CON | TN | 754 a | 763 a | 727 a | 741 a | 775 a | 55.5 | T ***, D ***, D × T + |
| HS | 749 a | 722 a | 718 a | 763 a | 696 b | D × T × D *** | |||
| BET | TN | 885 b | 827 b | 894 b | 947 b | 939 c | |||
| HS | 921 b | 852 b | 775 ab | 796 a | 797 a | ||||
| Villous area 3 (µm2) | CON | TN | 2.10 a | 1.90 a | 2.00 b | 2.06 a | 2.14 c | 0.22 | D ***, Day **, D × T ** |
| (130) | (82) | (104) | (118) | (140) | T × Day *, D × T × Day * | ||||
| HS | 1.98 a | 1.96 a | 1.87 a | 1.96 a | 1.92 a | ||||
| (100) | (95) | (76) | (92) | (85) | |||||
| BET | TN | 2.08 a | 2.14 b | 2.04 b | 2.19 b | 2.14 bc | |||
| (134) | (142) | (112) | (165) | (141) | |||||
| HS | 2.35 b | 2.09 b | 2.07 b | 2.17 b | 2.08 b | ||||
| (228) | (127) | (126) | (156) | (123) | |||||
| Crypt depth (µm) | CON | TN | 172 a | 183 b | 152 | 132 a | 144 b | 8.9 | D ***, Day *** |
| HS | 141 a | 197 bc | 162 | 126 a | 112 a | D × Day ***, T × Day ** | |||
| BET | TN | 250 b | 153 a | 170 | 211 c | 186 c | |||
| HS | 253 c | 217 c | 158 | 179 b | 151 b | ||||
| Seromuscular layer (µm) | CON | TN | 160 a | 175 a | 159 a | 160 a | 149 a | 11.4 | T ***, D ***, D × T *** |
| HS | 161 a | 185 a | 167 a | 145 a | 150 a | D × Day **, T × Day * | |||
| BET | TN | 282 b | 235 b | 259 b | 315 b | 375 c | D × T × Day ** | ||
| HS | 233 c | 209 a | 151 a | 172 a | 171 b | ||||
| Tissue | Diet (D) | Temp (T) | Day of Thermal Challenge | SED | Significance 1 | ||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 7 | 10 | |||||
| Ileum (µmol/g) | CON | TN | 77.7 a | 90.5 b | 95.7 a | 113 ab | 77.7 a | 21.3 | T ***, D ***, T × Day ** |
| HS | 68.1 a | 48.7 a | 106 a | 66 a | 68.1 a | D × T × Day * | |||
| BET | TN | 91 ab | 153 c | 152 b | 165 b | 91 ab | |||
| HS | 126 b | 90.1 b | 84.1 a | 89.4 ab | 126 b | ||||
| Kidney (µmol/g) | CON | TN | 41.2 a | 85.1 b | 54.1 | 47.8 a | 41.2 a | 8.3 | T ***, D ***, T × D * |
| HS | 48.1 a | 87 b | 64.1 | 56.7 ab | 48.1 a | Day ***, D × Day *** | |||
| BET | TN | 55.9 b | 53.9 a | 75.6 | 63.6 b | 55.9 b | |||
| HS | 90.4 c | 81.2 b | 77.6 | 88.8 c | 90.4 c | ||||
| Spleen (µmol/g) | CON | TN | 57.1 | 50.5 ab | 62.7 a | 60.1 a | 57.1 | 5.1 | T *, D ***, Day *** |
| HS | 62.5 | 42.5 a | 75.1 b | 50.9 a | 62.5 | D × T *, D × Day *** | |||
| BET | TN | 63.5 | 59.3 bc | 69.6 ab | 56.5 a | 63.5 | T × Day *, D × T × Day *** | ||
| HS | 52.5 | 68.5 c | 72.1 ab | 89.2 b | 52.5 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shakeri, M.; Cottrell, J.J.; Wilkinson, S.; Zhao, W.; Le, H.H.; McQuade, R.; Furness, J.B.; Dunshea, F.R. Dietary Betaine Improves Intestinal Barrier Function and Ameliorates the Impact of Heat Stress in Multiple Vital Organs as Measured by Evans Blue Dye in Broiler Chickens. Animals 2020, 10, 38. https://doi.org/10.3390/ani10010038
Shakeri M, Cottrell JJ, Wilkinson S, Zhao W, Le HH, McQuade R, Furness JB, Dunshea FR. Dietary Betaine Improves Intestinal Barrier Function and Ameliorates the Impact of Heat Stress in Multiple Vital Organs as Measured by Evans Blue Dye in Broiler Chickens. Animals. 2020; 10(1):38. https://doi.org/10.3390/ani10010038
Chicago/Turabian StyleShakeri, Majid, Jeremy James Cottrell, Stuart Wilkinson, Weicheng Zhao, Hieu Huu Le, Rachel McQuade, John Barton Furness, and Frank Rowland Dunshea. 2020. "Dietary Betaine Improves Intestinal Barrier Function and Ameliorates the Impact of Heat Stress in Multiple Vital Organs as Measured by Evans Blue Dye in Broiler Chickens" Animals 10, no. 1: 38. https://doi.org/10.3390/ani10010038
APA StyleShakeri, M., Cottrell, J. J., Wilkinson, S., Zhao, W., Le, H. H., McQuade, R., Furness, J. B., & Dunshea, F. R. (2020). Dietary Betaine Improves Intestinal Barrier Function and Ameliorates the Impact of Heat Stress in Multiple Vital Organs as Measured by Evans Blue Dye in Broiler Chickens. Animals, 10(1), 38. https://doi.org/10.3390/ani10010038