Role of the JAK-STAT Pathway in Bovine Mastitis and Milk Production

 ,
,  ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
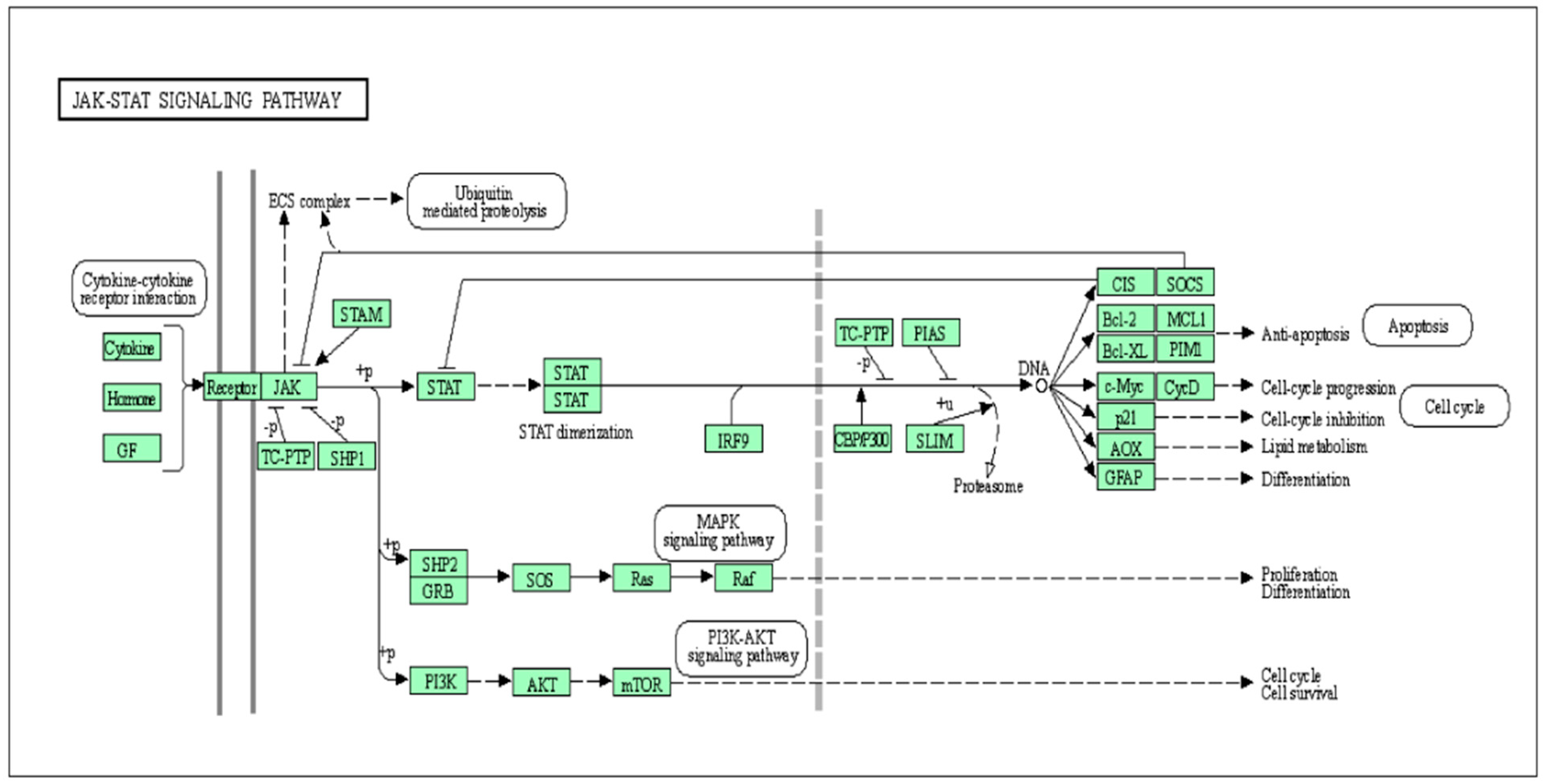
2. General Mechanism of the JAK-STAT Pathway Regulation
3. The JAK-STAT Pathway Role in Milk Production in Dairy Cattle
3.1. Role of JAKs in Milk Production in Dairy Cattle
3.2. Role of STATs in Milk Production in Dairy Cattle
4. The JAK-STAT Signaling Role in Bovine Mastitis
4.1. Association of JAK2 Activity with Bovine Mastitis
4.2. Role of STATs in Bovine Mastitis
5. Inhibitors of the JAK-STAT Pathway: Role in Mastitis and Milk Production
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Szyda, J.; Mielczarek, M.; Fraȩszczak, M.; Minozzi, G.; Williams, J.L.; Wojdak-Maksymiec, K. The genetic background of clinical mastitis in Holstein-Friesian cattle. Animal 2019, 13, 2156–2163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansor, R.; Mullen, W.; Albalat, A.; Zerefos, P.; Mischak, H.; Barrett, D.C.; Biggs, A.; Eckersall, P.D. A peptidomic approach to biomarker discovery for bovine mastitis. J. Proteom. 2013, 85, 89–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.J.; Wei, Z.K.; Zhang, X.; Wang, Y.N.; Fu, Y.H.; Yang, Z.T. Butyrate protects against disruption of the blood-milk barrier and moderates inflammatory responses in a model of mastitis induced by lipopolysaccharide. Br. J. Pharmacol. 2017, 174, 3811–3822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Liu, P.; Wang, S.; Zhao, G.; Zhang, T.; Guo, S.; Jiang, K.F.; Wu, H.C.; Deng, G. Shikonin exerts anti-inflammatory effects in LPS-induced mastitis by inhibiting NF-κB signaling pathway. Biochem. Biophys. Res. Commun. 2018, 505, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Pumipuntu, N.; Kulpeanprasit, S.; Santajit, S.; Tunyong, W.; Kong-ngoen, T.; Hinthong, W.; Indrawattana, N. Screening method for Staphylococcus aureus identification in subclinical bovine mastitis from dairy farms. Vet. World 2017, 10, 721–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, K.; Günther, J.; Talbot, R.; Petzl, W.; Zerbe, H.; Schuberth, H.J.; Seyfert, H.M.; Glass, E.J. Escherichia coli- and Staphylococcus aureus-induced mastitis differentially modulate transcriptional responses in neighbouring uninfected bovine mammary gland quarters. BMC Genom. 2013, 14, 36. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yang, F.; Wei, X.J.; Luo, Y.J.; Guo, W.Z.; Zhou, X.Z.; Guo, Z.T. Prevalence and risk factors of subclinical mastitis in lactating cows in Northwest China. Isr. J. Vet. Med. 2019, 74, 17–22. [Google Scholar]
- Seegers, H.; Fourichon, C.; Beaudeau, F. Production effects related to mastitis and mastitis economics in dairy cattle herds. Vet. Res. 2003, 34, 475–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamadani, H.; Khan, A.; Banday, M.; Ashraf, I.; Handoo, N.; Shah, A.; Hamadani, A. Bovine Mastitis—A Disease of Serious Concern for Dairy Farmers. Int. J. Livest. Res. 2013, 3, 42. [Google Scholar] [CrossRef] [Green Version]
- Viguier, C.; Arora, S.; Gilmartin, N.; Welbeck, K.; O’Kennedy, R. Mastitis detection: Current trends and future perspectives. Trends Biotechnol. 2009, 27, 486–493. Thompson-Crispi, K.; Atalla, H.; Miglior, F.; Mallard, B.A. Bovine mastitis: Frontiers in immunogenetics. Front. Immunol. 2014, 5, 1–10. [Google Scholar]
- Thompson-Crispi, K.; Atalla, H.; Miglior, F.; Mallard, B.A. Bovine mastitis: Frontiers in immunogenetics. Front. Immunol. 2014, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- González, R.N.; Wilson, D.J. Mycoplasmal mastitis in dairy herds. Vet. Clin. North Am. Food Anim. Pract. 2003, 19, 199–221. [Google Scholar] [CrossRef]
- Wellnitz, O.; Bruckmaier, R.M. The innate immune response of the bovine mammary gland to bacterial infection. Vet. J. 2012, 192, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Brenaut, P.; Lefèvre, L.; Rau, A.; Laloë, D.; Pisoni, G.; Moroni, P.; Bevilacqua, C.; Martin, P. Contribution of mammary epithelial cells to the immune response during early stages of a bacterial infection to Staphylococcus aureus. Vet. Res. 2014, 45. [Google Scholar] [CrossRef] [Green Version]
- Segal, S.; Hill, A.V.S. Genetic susceptibility to infectious disease. Trends Microbiol. 2003, 11, 445–448. [Google Scholar] [CrossRef]
- Albiger, B.; Dahlberg, S.; Henriques-Normark, B.; Normark, S. Role of the innate immune system in host defence against bacterial infections: Focus on the Toll-like receptors. J. Intern. Med. 2007, 261, 511–528. [Google Scholar] [CrossRef]
- Bhattarai, D.; Worku, T.; Dad, R.; Rehman, Z.U.; Gong, X.; Zhang, S. Mechanism of pattern recognition receptors (PRRs) and host pathogen interplay in bovine mastitis. Microb. Pathog. 2018, 120, 64–70. [Google Scholar] [CrossRef]
- Brooks, A.J.; Dai, W.; O’Mara, M.L.; Abankwa, D.; Chhabra, Y.; Pelekanos, R.A.; Gardon, O.; Tunny, K.A.; Blucher, K.M.; Morton, C.J.; et al. Mechanism of activation of protein kinase JAK2 by the growth hormone receptor. Science 2014, 344, 1249783. [Google Scholar] [CrossRef]
- Liongue, C.; Ward, A.C. Evolution of the JAK-STAT pathway. JAKSTAT 2013, 2, e22756. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Robinson, G.W.; Gouilleux, F.; Groner, B.; Hennighausen, L. Cloning and expression of Stat5 and an additional homologue (Stat5b) involved in prolactin signal transduction in mouse mammary tissue. Proc. Natl. Acad. Sci. USA 1995, 92, 8831–8835. [Google Scholar] [CrossRef] [Green Version]
- Hennighausen, L.; Robinson, W.G. Signaling Pathways in Mammary Gland Development. Dev. Cell 2001, 1, 467–475. [Google Scholar] [CrossRef] [Green Version]
- Furth, P.A.; Nakles, R.E.; Millman, S.; Diaz-Cruz, E.S.; Cabrera, M.C. Signal transducer and activator of transcription 5 as a key signaling pathway in normal mammary gland developmental biology and breast cancer. Breast Cancer Res. 2011, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ersahin, T.; Tuncbag, N.; Cetin-Atalay, R. The PI3K/AKT/mTOR interactive pathway. Mol. Biosyst. 2015, 11, 1946–1954. [Google Scholar] [CrossRef]
- Sobolewska, A.; Gajewska, M.; Zarzyńska, J.; Gajkowska, B.; Motyl, T. IGF-I, EGF, and sex steroids regulate autophagy in bovine mammary epithelial cells via the mTOR pathway. Eur. J. Cell Biol. 2009, 88, 117–130. [Google Scholar] [CrossRef]
- Schmidt, J.W.; Wehde, B.L.; Sakamoto, K.; Triplett, A.A.; Anderson, S.M.; Tsichlis, P.N.; Leone, G.; Wagner, K.-U. Stat5 Regulates the Phosphatidylinositol 3-Kinase/Akt1 Pathway during Mammary Gland Development and Tumorigenesis. Mol. Cell. Biol. 2014, 34, 1363–1377. [Google Scholar] [CrossRef] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Vafaizadeh, V.; Klemmt, P.A.B.; Groner, B. Stat5 assumes distinct functions in mammary gland development and mammary tumor formation. Front. Biosci. 2012, 17, 1232–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawlings, J.S.; Rosler, K.M.; Harrison, D.A. The JAK/STAT signaling pathway. J. Cell Sci. 2004, 117, 1281–1283. [Google Scholar] [CrossRef] [Green Version]
- Zhou, G.Y.; Yi, Y.X.; Jin, L.X.; Lin, W.; Fang, P.P.; Lin, X.Z.; Zheng, Y.; Pan, C.W. The protective effect of juglanin on fructose-induced hepatitis by inhibiting inflammation and apoptosis through TLR4 and JAK2/STAT3 signaling pathways in fructose-fed rats. Biomed. Pharmacother. 2016, 81, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, J.; Yuan, C.; Ji, Q.; Chen, D.; Zhao, H.; Jiang, W.; Ma, K.; Liu, L. JAK2/STAT3 is associated with the inflammatory process in periapical granuloma. Int. J. Clin. Exp. Pathol. 2019, 12, 190–197. [Google Scholar]
- Xin, P.; Xu, X.; Deng, C.; Liu, S.; Wang, Y.; Zhou, X.; Ma, H.; Wei, D.; Sun, S. The role of JAK/STAT signaling pathway and its inhibitors in diseases. Int. Immunopharmacol. 2020, 80, 106210. [Google Scholar] [CrossRef]
- Villarino, A.V.; Kanno, Y.; Shea, J.J.O. Mechanism and consequences of JAK/STAT signaling in the immune system. Nat. Immunol. 2017, 18, 374–384. [Google Scholar] [CrossRef]
- Kiu, H.; Nicholson, S.E. Biology and significance of the JAK/STAT signalling pathways. Growth Factors 2012, 30, 88–106. [Google Scholar] [CrossRef] [Green Version]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine mammary protein synthesis during the lactation cycle. Bioinform. Biol. Insights 2011, 5, 83–98. [Google Scholar] [CrossRef]
- Brenaut, P.; Bangera, R.; Bevilacqua, C.; Rebours, E.; Cebo, C.; Martin, P. Validation of RNA isolated from milk fat globules to profile mammary epithelial cell expression during lactation and transcriptional response to a bacterial infection. J. Dairy Sci. 2012, 95, 6130–6144. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, D.; Kang, K.; Robinson, G.W.; Hennighausen, L. Sequential activation of genetic programs in mouse mammary epithelium during pregnancy depends on STAT5A/Bconcentration. Nucleic Acids Res. 2013, 41, 1622–1636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennighausen, L.; Robinson, G.W.; Wagner, K.U.; Liu, X. Developing a mammary gland is a stat affair. J. Mammary Gland Biol. Neoplasia 1997, 2, 365–372. [Google Scholar] [CrossRef]
- Arun, S.J.; Thomson, P.C.; Sheehy, P.A.; Khatkar, M.S.; Raadsma, H.W.; Williamson, P. Targeted Analysis Reveals an Important Role of JAK-STAT-SOCS Genes for Milk Production Traits in Australian Dairy Cattle. Front. Genet. 2015, 6, 342. [Google Scholar] [CrossRef] [Green Version]
- Briscoe, J.; Guschin, D.; Müller, M. Signal Transduction: Just another signalling pathway. Curr. Biol. 1994, 4, 1033–1035. [Google Scholar] [CrossRef]
- Groner, B.; Gouilleux, F. Prolactin-mediated gene activation in mammary epithelial cells. Curr. Opin. Genet. Dev. 1995, 5, 587–594. [Google Scholar] [CrossRef]
- Sigl, T.; Meyer, H.H.D.; Wiedemann, S. Gene expression analysis of protein synthesis pathways in bovine mammary epithelial cells purified from milk during lactation and short-term restricted feeding. J. Anim. Physiol. Anim. Nutr. 2014, 98, 84–95. [Google Scholar] [CrossRef]
- Tian, Q.; Wang, H.R.; Wang, M.Z.; Wang, C.; Liu, S.M. Lactogenic hormones regulate mammary protein synthesis in bovine mammary epithelial cells via the mTOR and JAK-STAT signal pathways. Anim. Prod. Sci. 2016, 56, 1803–1809. [Google Scholar] [CrossRef]
- Bole-Feysot, C. Prolactin (PRL) and Its Receptor: Actions, Signal Transduction Pathways and Phenotypes Observed in PRL Receptor Knockout Mice. Endocr. Rev. 1998, 19, 225–268. [Google Scholar] [CrossRef]
- Auchtung, T.L.; Rius, A.G.; Kendall, P.E.; McFadden, T.B.; Dahl, G.E. Effects of photoperiod during the dry period on prolactin, prolactin receptor, and milk production of dairy cows. J. Dairy Sci. 2005, 88, 121–127. [Google Scholar] [CrossRef]
- Campo Verde Arboccó, F.; Persia, F.A.; Hapon, M.B.; Jahn, G.A. Hypothyroidism decreases JAK/STAT signaling pathway in lactating rat mammary gland. Mol. Cell. Endocrinol. 2017, 450, 14–23. [Google Scholar] [CrossRef]
- Shillingford, J.M.; Miyoshi, K.; Robinson, G.W.; Grimm, S.L.; Rosen, J.M.; Neubauer, H.; Pfeffer, K.; Hennighausen, L. Jak2 is an essential tyrosine kinase involved in pregnancy-mediated development of mammary secretory epithelium. Mol. Endocrinol. 2002, 16, 563–570. [Google Scholar] [CrossRef]
- Etherton, T.D.; Bauman, D.E. Biology of somatotropin in growth and lactation of domestic animals. Physiol. Rev. 1998, 78, 745–761. [Google Scholar] [CrossRef]
- Liu, X.F.; Li, M.; Li, Q.Z.; Lu, L.M.; Tong, H.L.; Gao, X.J. Stat5a increases lactation of dairy cow mammary gland epithelial cells cultured in vitro. Vitr. Cell. Dev. Biol. Anim. 2012, 48, 554–561. [Google Scholar] [CrossRef]
- Ali, N.; Niaz, S.; Khan, N.U.; Gohar, A.; Khattak, I.; Dong, Y.; Khattak, T.; Ahmad, I.; Wang, Y.; Usman, T. Polymorphisms in JAK2 Gene are Associated with Production Traits and Mastitis Resistance in Dairy Cattle. Ann. Anim. Sci. 2019, 20, 409–423. [Google Scholar] [CrossRef]
- Szewczuk, M. Association of a genetic marker at the bovine Janus kinase 2 locus (JAK2/RsaI) with milk production traits of four cattle breeds. J. Dairy Res. 2015, 82, 287–292. [Google Scholar] [CrossRef]
- Khan, M.Z.; Wang, D.; Liu, L.; Usman, T.; Wen, H.; Zhang, R.; Liu, S.; Shi, L.; Mi, S.; Xiao, W.; et al. Significant genetic effects of JAK2 and DGAT1 mutations on milk fat content and mastitis resistance in Holsteins. J. Dairy Res. 2019, 86, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Meredith, B.K.; Kearney, F.J.; Finlay, E.K.; Bradley, D.G.; Fahey, A.G.; Berry, D.P.; Lynn, D.J. Genome-wide associations for milk production and somatic cell score in Holstein-Friesian cattle in Ireland. BMC Genet. 2012, 13, 21. [Google Scholar] [CrossRef] [Green Version]
- Verdier, F.; Rabionet, R.; Gouilleux, F.; Beisenherz-Huss, C.; Varlet, P.; Muller, O.; Mayeux, P.; Lacombe, C.; Gisselbrecht, S.; Chretien, S. A sequence of the CIS gene promoter interacts preferentially with two associated STAT5A dimers: A distinct biochemical difference between STAT5A and STAT5B. Mol. Cell. Biol. 1998, 20, 389–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, B.; Zhang, C.; Fang, X.; Zhang, R.; Gu, C.; Lei, C.; Chen, H. Association between polymorphism in STAT5A gene and milk production traits in chinese holstein cattle. Anim. Sci. Pap. Rep. 2010, 28, 5–11. [Google Scholar]
- Liu, X.; Robinson, G.W.; Wagner, K.U.; Garrett, L.; Wynshaw-Boris, A.; Hennighausen, L. Stat5a is mandatory for adult mammary gland development and lactogenesis. Genes Dev. 1997, 11, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Bionaz, M.; Periasamy, K.; Rodriguez-Zas, S.L.; Everts, R.E.; Lewin, H.A.; Hurley, W.L.; Loor, J.J. Old and new stories: Revelations from functional analysis of the bovine mammary transcriptome during the lactation cycle. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Lacasse, P.; Ollier, S.; Lollivier, V.; Boutinaud, M. New insights into the importance of prolactin in dairy ruminants. J. Dairy Sci. 2016, 99, 864–874. [Google Scholar] [CrossRef]
- Watson, C.J.; Neoh, K. The Stat family of transcription factors have diverse roles in mammary gland development. Semin. Cell. Dev. Biol. 2008, 19, 401–406. [Google Scholar] [CrossRef]
- Teglund, S.; McKay, C.; Schuetz, E.; Van Deursen, J.M.; Stravopodis, D.; Wang, D.; Brown, M.; Bodner, S.; Grosveld, G.; Ihle, J.N. Stat5a and Stat5b proteins have essential and nonessential, or redundant, roles in cytokine responses. Cell 1998, 93, 841–850. [Google Scholar] [CrossRef] [Green Version]
- Dong, B.; Zhao, F.Q. Involvement of the ubiquitous Oct-1 transcription factor in hormonal induction of β-casein gene expression. Biochem. J. 2007, 401, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Ji, M.-R.; Lee, S.I.; Jang, Y.J.; Jeon, M.-H.; Kim, J.S.; Kim, K.W.; Park, J.K.; Yoo, J.G.; Jeon, I.S.; Kwon, D.J.; et al. STAT5 plays a critical role in regulating the 5′-flanking region of the porcine whey acidic protein gene in transgenic mice. Mol. Reprod. Dev. 2015, 82, 957–966. [Google Scholar] [CrossRef]
- Watson, C.J. Stat transcription factors in mammary gland development and tumorigenesis. J. Mammary Gland Biol. Neoplasia. 2001, 6, 115–127. [Google Scholar] [CrossRef]
- Boutinaud, M.; Jammes, H. Growth hormone increases Stat5 and Stat1 expression in lactating goat mammary gland: A specific effect compared to milking frequency. Domest. Anim. Endocrinol. 2004, 27, 363–378. [Google Scholar] [CrossRef]
- Sadeghi, M.; Shahrbabak, M.M.; Mianji, G.R.; Javaremi, A.N. Polymorphism at locus of STAT5A and its association with breeding values of milk production traits in Iranian Holstein bulls. Livest. Sci. 2009, 123, 97–100. [Google Scholar] [CrossRef]
- Selvaggi, M.; Albarella, S.; Dario, C.; Peretti, V.; Ciotola, F. Association of STAT5A Gene Variants with Milk Production Traits in Agerolese Cattle. Biochem. Genet. 2017, 55, 158–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvaggi, M.; Dario, C.; Normanno, G.; Celano, G.V.; Dario, M. Genetic polymorphism of STAT5A protein: Relationships with production traits and milk composition in Italian Brown cattle. J. Dairy Res. 2009, 76, 441–445. [Google Scholar] [CrossRef] [Green Version]
- Dario, C.; Selvaggi, M. Study on the STAT5A/AvaI polymorphism in Jersey cows and association with milk production traits. Mol. Biol. Rep. 2011, 38, 5387–5392. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Chu, M.X.; Qiao, L.; He, J.N.; Wang, P.Q.; Feng, T.; Di, R.; Cao, G.L.; Fang, L.; An, Y.F. Polymorphisms of STAT5A gene and their association with milk production traits in Holstein cows. Mol. Biol. Rep. 2012, 39, 2901–2907. [Google Scholar] [CrossRef]
- Schennink, A.; Bovenhuis, H.; Léon-Kloosterziel, K.M.; Van Arendonk, J.A.M.; Visker, M.H.P.W. Effect of polymorphisms in the FASN, OLR1, PPARGC1A, PRL and STAT5A genes on bovine milk-fat composition. Anim. Genet. 2009, 40, 909–916. [Google Scholar] [CrossRef]
- Khatib, H.; Monson, R.L.; Schutzkus, V.; Kohl, D.M.; Rosa, G.J.M.; Rutledge, J.J. Mutations in the STAT5A gene are associated with embryonic survival and milk composition in cattle. J. Dairy Sci. 2008, 91, 784–793. [Google Scholar] [CrossRef]
- He, Y.; Chu, Q.; Ma, P.; Wang, Y.; Zhang, Q.; Sun, D.; Zhang, Y.; Yu, Y.; Zhang, Y. Association of bovine CD4 and STAT5b single nucleotide polymorphisms with somatic cell scores and milk production traits in Chinese Holsteins. J. Dairy Res. 2011, 78, 242–249. [Google Scholar] [CrossRef]
- Li, S.; Rosen, J.M. Nuclear factor I and mammary gland factor (STAT5) play a critical role in regulating rat whey acidic protein gene expression in transgenic mice. Mol. Cell. Biol. 1995, 15, 2063–2070. [Google Scholar] [CrossRef] [Green Version]
- Cobanoglu, O.; Zaitoun, I.; Chang, Y.M.; Shook, G.E.; Khatib, H. Effects of the Signal Transducer and Activator of Transcription 1 (STAT1) Gene on Milk Production Traits in Holstein Dairy Cattle. J. Dairy Sci. 2006, 89, 4433–4437. [Google Scholar] [CrossRef] [Green Version]
- Cobanoglu, O.; Zaitoun, I.; Chang, Y.M.; Shook, G.E.; Khatib, H. The Detection of STAT1 Gene Influencing Milk Related Traits in Turkish Holstein and Jersey Cows. J. Agric. Sci. Technol. A 2016, 6, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Deng, T.X.; Pang, C.Y.; Lu, X.R.; Zhu, P.; Duan, A.Q.; Liang, X.W. Associations between polymorphisms of the STAT1 gene and milk production traits in water buffaloes1. J. Anim. Sci. 2016, 94, 927–935. [Google Scholar] [CrossRef]
- Buitenhuis, B.; Røntved, C.M.; Edwards, S.M.; Ingvartsen, K.L.; Sørensen, P. In depth analysis of genes and pathways of the mammary gland involved in the pathogenesis of bovine Escherichia coli-mastitis. BMC Genom. 2011, 12. [Google Scholar] [CrossRef] [Green Version]
- Tiezzi, F.; Parker-Gaddis, K.L.; Cole, J.B.; Clay, J.S.; Maltecca, C. A Genome-Wide Association Study for Clinical Mastitis in First Parity US Holstein Cows Using Single-Step Approach and Genomic Matrix Re-Weighting Procedure. PLoS ONE 2015, 10, e0114919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, H.S.; Shin, J.S.; Lee, S.B.; Park, J.C.; Lee, K.T. Cirsimarin, a flavone glucoside from the aerial part of Cirsium japonicum var. ussuriense (Regel) Kitam. ex Ohwi, suppresses the JAK/STAT and IRF-3 signaling pathway in LPS-stimulated RAW 264.7 macrophages. Chem. Biol. Interact. 2018, 293, 38–47. [Google Scholar] [CrossRef]
- Li, J.; Yin, P.; Gong, P.; Lv, A.; Zhang, Z.; Liu, F. 8-Methoxypsoralen protects bovine mammary epithelial cells against lipopolysaccharide-induced inflammatory injury via suppressing JAK/STAT and NF-κB pathway. Microbiol. Immunol. 2019, 63, 427–437. [Google Scholar] [CrossRef]
- Lee, S.B.; Lee, W.S.; Shin, J.S.; Jang, D.S.; Lee, K.T. Xanthotoxin suppresses LPS-induced expression of iNOS, COX-2, TNF-α, and IL-6 via AP-1, NF-κB, and JAK-STAT inactivation in RAW 264.7 macrophages. Int. Immunopharmacol. 2017, 49, 21–29. [Google Scholar] [CrossRef]
- Boutet, P.; Boulanger, D.; Gillet, L.; Vanderplasschen, A.; Closset, R.; Bureau, F.; Lekeux, P. Delayed neutrophil apoptosis in bovine subclinical mastitis. J. Dairy Sci. 2004, 87, 4104–4114. [Google Scholar] [CrossRef]
- Moyes, K.M.; Drackley, J.K.; Morin, D.E.; Bionaz, M.; Rodriguez-Zas, S.L.; Everts, R.E.; Lewin, H.A.; Loor, J.J. Gene network and pathway analysis of bovine mammary tissue challenged with Streptococcus uberis reveals induction of cell proliferation and inhibition of PPAR signaling as potential mechanism for the negative relationships between immune response and lipi. BMC Genom. 2009, 10. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Sørensen, P.; Thomsen, B.; Edwards, S.M.; Skarman, A.; Røntved, C.M.; Lund, M.S.; Workman, C.T. Gene prioritization for livestock diseases by data integration. Physiol. Gen. 2011, 44, 283–330. [Google Scholar] [CrossRef] [Green Version]
- Yue, Y.; QiuLing, L.; ZhiHua, J.; JinMing, H.; Lei, Z.; RongLing, L.; JianBin, L.; FangXiong, S.; JiFeng, Z.; ChangFa, W. Three novel single-nucleotide polymorphisms of complement component 4 gene (C4A) in Chinese Holstein cattle and their associations with milk performance traits and CH50. Vet. Immunol. Immunopathol. 2012, 145, 223–232. [Google Scholar]
- Usman, T.; Yu, Y.; Liu, C.; Wang, X.; Zhang, Q.; Wang, Y. Genetic effects of single nucleotide polymorphisms in JAK2 and STAT5A genes on susceptibility of Chinese Holsteins to mastitis. Mol. Biol. Rep. 2014, 41, 8293–8301. [Google Scholar] [CrossRef]
- Usman, T.; Wang, Y.; Liu, C.; Wang, X.; Zhang, Y.; Yu, Y. Association study of single nucleotide polymorphisms in JAK2 and STAT5B genes and their differential mRNA expression with mastitis susceptibility in Chinese Holstein cattle. Anim. Genet. 2015, 46, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Weigel, K.A.; Shook, G.E. Genetic Selection for Mastitis Resistance. Vet. Clin. N. Am. Food Anim. Pract. 2018, 34, 457–472. [Google Scholar] [CrossRef]
- Pilla, R.; Schwarz, D.; König, S.; Piccinini, R. Microscopic differential cell counting to identify inflammatory reactions in dairy cow quarter milk samples. J. Dairy Sci. 2012, 95, 4410–4420. [Google Scholar] [CrossRef]
- Schwarz, D.; Rivas, A.L.; König, S.; Diesterbeck, U.S.; Schlez, K.; Zschöck, M.; Wolter, W.; Czerny, C.P. CD2/CD21 index: A new marker to evaluate udder health in dairy cows. J. Dairy Sci. 2013, 96, 5106–5119. [Google Scholar] [CrossRef]
- Usman, T.; Wang, Y.; Song, M.; Wang, X.; Dong, Y.; Liu, C.; Wang, S.; Zhang, Y.; Xiao, W.; Yu, Y. Novel polymorphisms in bovine CD4 and LAG-3 genes associated with somatic cell counts of clinical mastitis cows. Genet. Mol. Res. 2017, 16. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhang, J.; Zhou, Y.H.; Jiang, Y.N.; Zhang, W.; Tang, X.J.; Ren, Y.; Han, S.P.; Liu, P.J.; Xu, J.; et al. IL-6/STAT3 signaling pathway is activated in plasma cell mastitis. Int. J. Clin. Exp. Pathol. 2015, 8, 12541–12548. [Google Scholar]
- Milner, J.D. IL-17 producing cells in host defense and atopy. Curr. Opin. Immunol. 2011, 23, 784–788. [Google Scholar] [CrossRef]
- Cua, D.J.; Tato, C.M. Innate IL-17-producing cells: The sentinels of the immune system. Nat. Rev. Immunol. 2010, 10, 479–489. [Google Scholar] [CrossRef]
- Shibata, K.; Yamada, H.; Hara, H.; Kishihara, K.; Yoshikai, Y. Resident Vδ1 + γδ T Cells Control Early Infiltration of Neutrophils after Escherichia coli Infection via IL-17 Production. J. Immunol. 2007, 178, 4466–4472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, X.Q.; Zhao, Y.Q.; Shang, C.C.; Yao, Y.L.; Tian, T.T.; Li, J.; Chen, D.K. Dynamics of cytokines associated with IL-17 producing cells in serum and milk in mastitis of experimental challenging with Staphylococcus aureus and Escherichia coli in dairy goats. J. Anim. Vet. Adv. 2012, 11, 475–479. [Google Scholar]
- Tassi, R.; McNeilly, T.N.; Fitzpatrick, J.L.; Fontaine, M.C.; Reddick, D.; Ramage, C.; Lutton, M.; Schukken, Y.H.; Zadoks, R.N. Strain-specific pathogenicity of putative host-adapted and nonadapted strains of Streptococcus uberis in dairy cattle. J. Dairy Sci. 2013, 96, 5129–5145. [Google Scholar] [CrossRef] [Green Version]
- Tao, W.; Mallard, B. Differentially expressed genes associated with Staphylococcus aureus mastitis of Canadian Holstein cows. Vet. Immunol. Immunopathol. 2007, 120, 201–211. [Google Scholar] [CrossRef]
- Bougarn, S.; Cunha, P.; Gilbert, F.B.; Harmache, A.; Foucras, G.; Rainard, P. Staphylococcal-associated molecular patterns enhance expression of immune defense genes induced by IL-17 in mammary epithelial cells. Cytokine 2011, 56, 749–759. [Google Scholar] [CrossRef]
- Roussel, P.; Cunha, P.; Porcherie, A.; Petzl, W.; Gilbert, F.B.; Riollet, C.; Zerbe, H.; Rainard, P.; Germon, P. Investigating the contribution of IL-17A and IL-17F to the host response during Escherichia coli mastitis. Vet. Res. 2015, 46. [Google Scholar] [CrossRef] [Green Version]
- Ruddy, M.J.; Wong, G.C.; Liu, X.K.; Yamamoto, H.; Kasayama, S.; Kirkwood, K.L.; Gaffen, S.L. Functional Cooperation between Interleukin-17 and Tumor Necrosis Factor-α Is Mediated by CCAAT/Enhancer-binding Protein Family Members. J. Biol. Chem. 2004, 279, 2559–2567. [Google Scholar] [CrossRef] [Green Version]
- Wilson, C.B.; Rowell, E.; Sekimata, M. Epigenetic control of T-helper-cell differentiation. Nat. Rev. Immunol. 2009, 9, 91–105. [Google Scholar] [CrossRef]
- Zahoor, A.; Yang, Y.; Yang, C.; Khan, S.B.; Reix, C.; Anwar, F.; Deng, G. MerTK negatively regulates Staphylococcus aureus induced inflammatory response via Toll-like receptor signaling in the mammary gland. Mol. Immunol. 2020, 122, 1–12. [Google Scholar] [CrossRef]
- Mehrzad, J.; Zhao, X. T lymphocyte proliferative capacity and CD4+/CD8+ ratio in primiparous and pluriparous lactating cows. J. Dairy Res. 2008, 75, 457–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas, A.L.; Schwager, S.J.; González, R.N.; Quimby, F.W.; Anderson, K.L. Multifactorial relationships between intramammary invasion by Staphylococcus aureus and bovine leukocyte markers. Can. J. Vet. Res. 2007, 71, 135–144. [Google Scholar]
- Eder, J.M.; Gorden, P.J.; Lippolis, J.D.; Reinhardt, T.A.; Sacco, R.E. Lactation stage impacts the glycolytic function of bovine CD4+ T cells during ex vivo activation. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef] [Green Version]
- Wei, G.; Wei, L.; Zhu, J.; Zang, C.; Hu-Li, J.; Yao, Z.; Cui, K.; Kanno, Y.; Roh, T.Y.; Watford, W.T.; et al. Global Mapping of H3K4me3 and H3K27me3 Reveals Specificity and Plasticity in Lineage Fate Determination of Differentiating CD4+ T Cells. Immunity 2009, 30, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Bochniarz, M.; Zdzisińska, B.; Wawron, W.; Szczubiał, M.; Dąbrowski, R. Milk and serum IL-4, IL-6, IL-10, and amyloid A concentrations in cows with subclinical mastitis caused by coagulase-negative staphylococci. J. Dairy Sci. 2017, 100, 9674–9680. [Google Scholar] [CrossRef] [Green Version]
- Kiyici, M.J.; Bilal, A.; Mahmut, K.; Korhan, A.; Esma, G.A.; Mehmet, U.C. Association of GH, STAT5A, MYF5 gene polymorphisms with milk somatic cell count, EC and pH levels of Holstein dairy cattle. Ani. Biotechnol. 2020. [Google Scholar] [CrossRef]
- Cai, Z.; Guldbrandtsen, B.; Lund, M.S.; Sahana, G. Prioritizing candidate genes for fertility in dairy cows using gene-based analysis, functional annotation and differential gene expression. BMC Genom. 2019, 20, 255. [Google Scholar] [CrossRef]
- Shuai, K. Regulation of cytokine signaling pathways by PIAS proteins. Cell Res. 2006, 16, 196–202. [Google Scholar] [CrossRef] [Green Version]
- Henenstreit, D.; Horeks-Hoeck, J.; Duschl, A. JAK/STAT-dependent gene regulation by cytokines. Drug News Perspect. 2005, 18, 243. [Google Scholar] [CrossRef]
- Krebs, D.L.; Hilton, D.J. SOCS proteins: Negative regulators of cytokine signaling. Stem Cells 2001, 19, 378–387. [Google Scholar] [CrossRef] [PubMed]
- Niu, G.J.; Xu, J.D.; Yuan, W.J.; Sun, J.J.; Yang, M.C.; He, Z.H.; Zhao, X.F.; Wang, J.X. Protein Inhibitor of Activated STAT (PIAS) Negatively Regulates the JAK/STAT Pathway by Inhibiting STAT Phosphorylation and Translocation. Front. Immunol. 2018, 9, 2392. [Google Scholar] [CrossRef]
- Sharrocks, A.D. PIAS proteins and transcriptional regulation—More than just SUMO E3 ligases? Genes Dev. 2006, 20, 754–758. [Google Scholar] [CrossRef] [Green Version]
- Shuai, K.; Liu, B.; Zhang, D.; Cui, Y.; Zhou, J.; Sheng, C. Regulation of gene-activation pathways by PIAS proteins in the immune system. Nat. Rev. Immunol. 2005, 5, 593–605. [Google Scholar] [CrossRef]
- Ungureanu, D.; Vanhatupa, S.; Grönholm, J.; Palvimo, J.; Silvennoinen, O. SUMO-1 conjugation selectively modulates STAT1-mediated gene responses. Blood 2005, 106, 224–226. [Google Scholar] [CrossRef] [Green Version]
- Chung, C.D.; Liao, J.; Liu, B.; Rao, X.; Jay, P.; Berta, P.; Shuai, K. Specific inhibition of Stat3 signal transduction by PIAS3. Science 1997, 278, 1803–1805. [Google Scholar] [CrossRef]
- Tolkunova, E.; Malashicheva, A.; Parfenov, V.N.; Sustmann, C.; Grosschedl, R.; Tomilin, A. PIAS proteins as repressors of Oct4 function. J. Mol. Biol. 2007, 374, 1200–1212. [Google Scholar] [CrossRef]
- Xu, D.; Qu, C. Protein tyrosine phosphatases in the JAK/STAT pathway. Front. Biosci. 2008, 13, 4925–4932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latanya, S.M.; Harshani, L.R.; Said, S.M.J.; Nicholas, L.J.; Wu, J. Targeting Protein Tyrosine Phosphatases for Anticancer Drug Discovery. Curr. Pharm. Des. 2010, 16, 1843–1862. [Google Scholar]
- Heather, B.; Dechert, U.; Jirik, F.; Schrader, J.W.; Welham, M.J. SHP1 and SHP2 Protein-tyrosine Phosphatases Associate with βc after Interleukin-3-induced Receptor Tyrosine Phosphorylation. J. Biol. Chem. 1997, 272, 14470–14476. [Google Scholar]
- Böhmer, F.; Friedrich, K. Protein tyrosine phosphatases as wardens of STAT signaling. JAKSTAT 2014, 3, e28087. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, A. Negative regulation of cytokine signaling. Clin. Rev. Allergy Immunol. 2005, 28, 205–220. [Google Scholar] [CrossRef]
- Collins, A.S.; McCoy, C.E.; Lloyd, A.T.; O’Farrelly, C.; Stevenson, N.J. miR-19a: An Effective Regulator of SOCS3 and Enhancer of JAK-STAT Signalling. PLoS ONE 2013, 8, e69090. [Google Scholar] [CrossRef]
- Kubo, M.; Hanada, T.; Yoshimura, A. Suppressors of cytokine signaling and immunity. Nat. Immunol. 2003, 4, 1169–1176. [Google Scholar] [CrossRef]
- Flowers, L.O.; Johnson, H.M.; Mujtaba, M.G.; Ellis, M.R.; Haider, S.M.I.; Subramaniam, P.S. Characterization of a Peptide Inhibitor of Janus Kinase 2 That Mimics Suppressor of Cytokine Signaling 1 Function. J. Immunol. 2004, 172, 7510–7518. [Google Scholar] [CrossRef] [Green Version]
- Waiboci, L.W.; Ahmed, C.M.; Mujtaba, M.G.; Flowers, L.O.; Martin, J.P.; Haider, M.I.; Johnson, H.M. Both the Suppressor of Cytokine Signaling 1 (SOCS-1) Kinase Inhibitory Region and SOCS-1 Mimetic Bind to JAK2 Autophosphorylation Site: Implications for the Development of a SOCS-1 Antagonist. J. Immunol. 2007, 178, 5058–5068. [Google Scholar] [CrossRef] [Green Version]
- Kershaw, N.J.; Murphy, J.M.; Lucet, I.S.; Nicola, N.A.; Babon, J.J. Regulation of Janus kinases by SOCS proteins. Biochem. Soc. Trans. 2013, 41, 1042–1047. [Google Scholar] [CrossRef]
- Kimura, A.; Tetsuji, N.; Tatsushi, M.; Osamu, T.; Shizuo, A.; Ichiro, K.; Tadamitsu, K. Suppressor of cytokine signaling-1 selectively inhibits LPS-induced IL-6 production by regulating JAK-STAT. Blood 2004, 102, 17089–17094. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.Z.; Zhang, Z.; Liu, L.; Wang, D.; Mi, S.; Liu, X.; Liu, G.; Guo, G.; Li, X.; Wang, Y.; et al. Folic acid supplementation regulates key immunity-associated genes and pathways during the periparturient period in dairy cows. Asian Australas. J. Anim. Sci. 2019, 33, 1507–1519. [Google Scholar] [CrossRef] [Green Version]
- Yasukawa, H.; Ohishi, M.; Mori, H.; Murakami, M.; Chinen, T.; Aki, D.; Hanada, T.; Takeda, K.; Akira, S.; Hoshijima, M.; et al. IL-6 induces an anti-inflammatory response in the absence of SOCS3 in macrophages. Nat. Immunol. 2003, 4, 551–556. [Google Scholar] [CrossRef]
- Fang, L.; Hou, Y.; An, J.; Li, B.; Song, M.; Wang, X.; Sørensen, P.; Dong, Y.; Liu, C.; Wang, Y.; et al. Genome-wide transcriptional and post-transcriptional regulation of innate immune and defense responses of bovine mammary gland to Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Zhao, F.; Luo, C.; Zhang, X.; Yu, S.; Sun, Z.; Li, Z.; Li, Q.; Gao, X. SOCS3-Mediated Blockade Reveals Major Contribution of JAK2/STAT5 Signaling Pathway to Lactation and Proliferation of Dairy Cow Mammary Epithelial Cells in Vitro. Molecules 2013, 18, 12987–13002. [Google Scholar] [CrossRef] [Green Version]
- Dif, F.; Saunier, E.; Demeneix, B.; Kelly, P.A.; Edery, M. Cytokine-inducible SH2-containing protein suppresses PRL signaling by binding the PRL receptor. Endocrinology 2001, 142, 5286–5293. [Google Scholar] [CrossRef]
- Rupp, R.; Senin, P.; Sarry, J.; Allain, C.; Tasca, C.; Ligat, L. A Point Mutation in Suppressor of Cytokine Signalling 2 (Socs2) Increases the Susceptibility to Inflammation of the Mammary Gland while Associated with Higher Body Weight and Size and Higher Milk Production in a Sheep Model. PLoS Genet. 2015, 11, e1005629. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Gene | Mutation | Reference | Position | Phenotypic Traits | Authors |
|---|---|---|---|---|---|
| JAK2 | C-T/EXON16 | rs210148032 | Chr8:39652267 | SCC | [51,85] |
| JAK2 | C-T/EXON20 | rs110298451 | Chr8:39645396 | IL-6, IFN-γ, SCC | [51,86] |
| JAK2 | C-T/3′flanking region | rs135128681 | Chr8:39630048 | IL-6, IFN-γ, SCC | [51,85] |
| JAK2 | T-C/3′flanking region | Novel | Chr8:39631175 | IL-6, SCC | [51,85] |
| JAK2 | G-A/3′flanking region | Novel | Chr8:39631044 | SCS | [85] |
| JAK2 | 5′ upstream | rs379754157 | Chr8:39750638 | SCC | [49] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.Z.; Khan, A.; Xiao, J.; Ma, Y.; Ma, J.; Gao, J.; Cao, Z. Role of the JAK-STAT Pathway in Bovine Mastitis and Milk Production. Animals 2020, 10, 2107. https://doi.org/10.3390/ani10112107
Khan MZ, Khan A, Xiao J, Ma Y, Ma J, Gao J, Cao Z. Role of the JAK-STAT Pathway in Bovine Mastitis and Milk Production. Animals. 2020; 10(11):2107. https://doi.org/10.3390/ani10112107
Chicago/Turabian StyleKhan, Muhammad Zahoor, Adnan Khan, Jianxin Xiao, Yulin Ma, Jiaying Ma, Jian Gao, and Zhijun Cao. 2020. "Role of the JAK-STAT Pathway in Bovine Mastitis and Milk Production" Animals 10, no. 11: 2107. https://doi.org/10.3390/ani10112107
APA StyleKhan, M. Z., Khan, A., Xiao, J., Ma, Y., Ma, J., Gao, J., & Cao, Z. (2020). Role of the JAK-STAT Pathway in Bovine Mastitis and Milk Production. Animals, 10(11), 2107. https://doi.org/10.3390/ani10112107