Liver Expression of IGF2 and Related Proteins in ZBED6 Gene-Edited Pig by RNA-Seq

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples Collection and Hematological and Blood Biochemical Indices Analysis
2.2. RNA Isolation and Library Preparation
2.3. Primers Design and qRT-PCR
2.4. ZBED6 SKO Efficiency in RNA Level
2.5. Western Blotting
2.6. RNA-seq Analysis and Statistical
3. Results
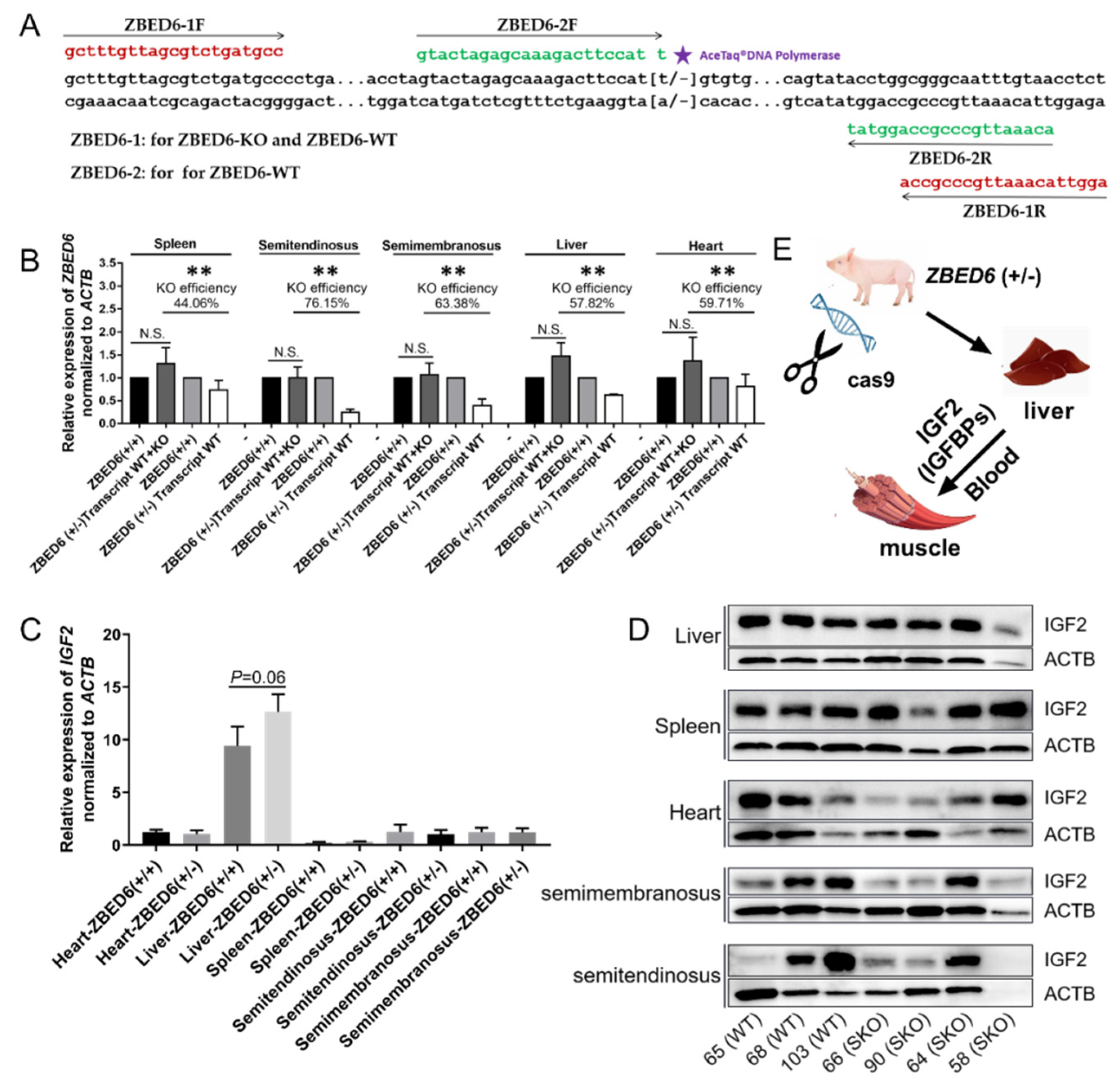
3.1. Detection of ZBED6 Gene SKO Efficiency in Pig Tissues and Blood Routine Examinations
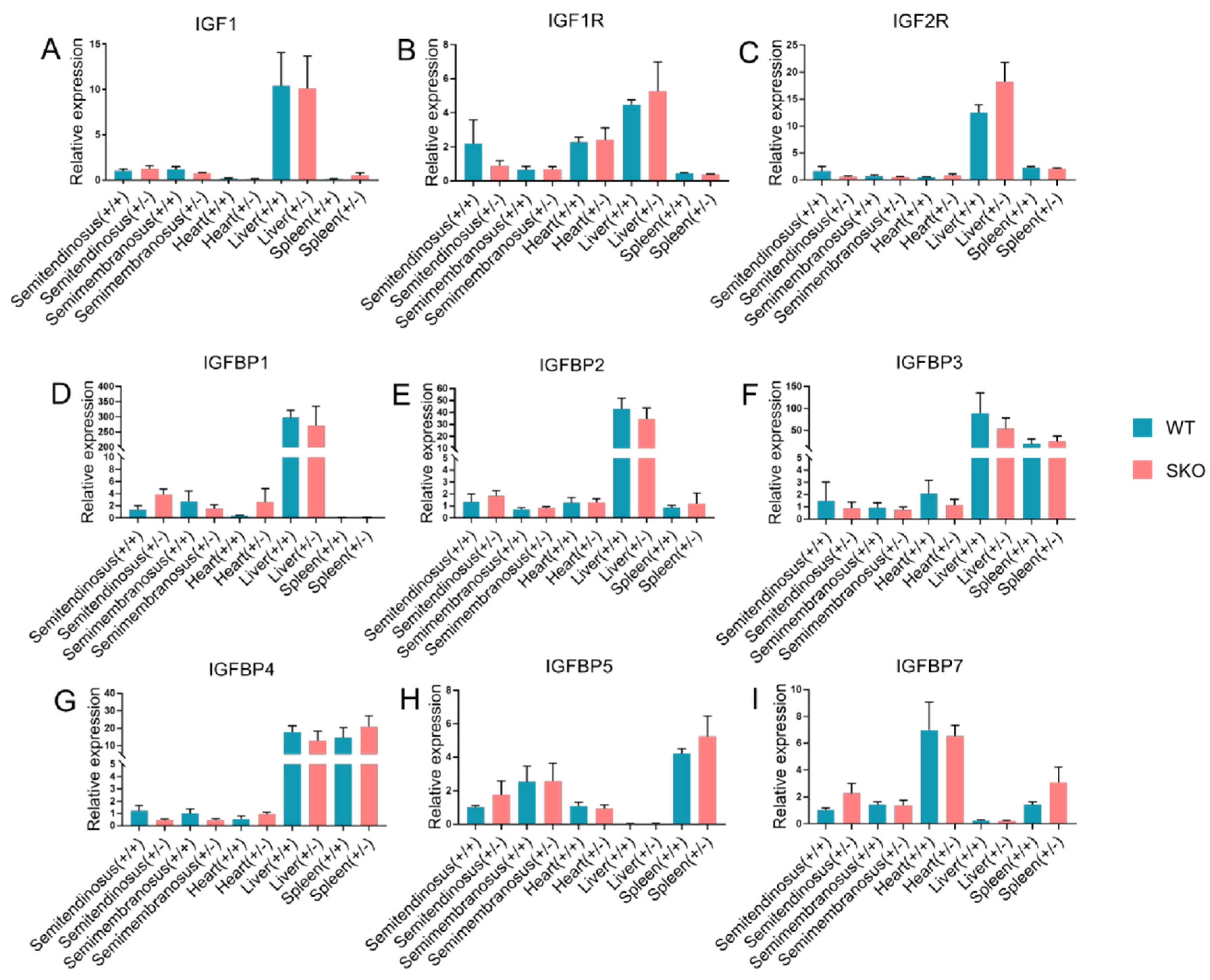
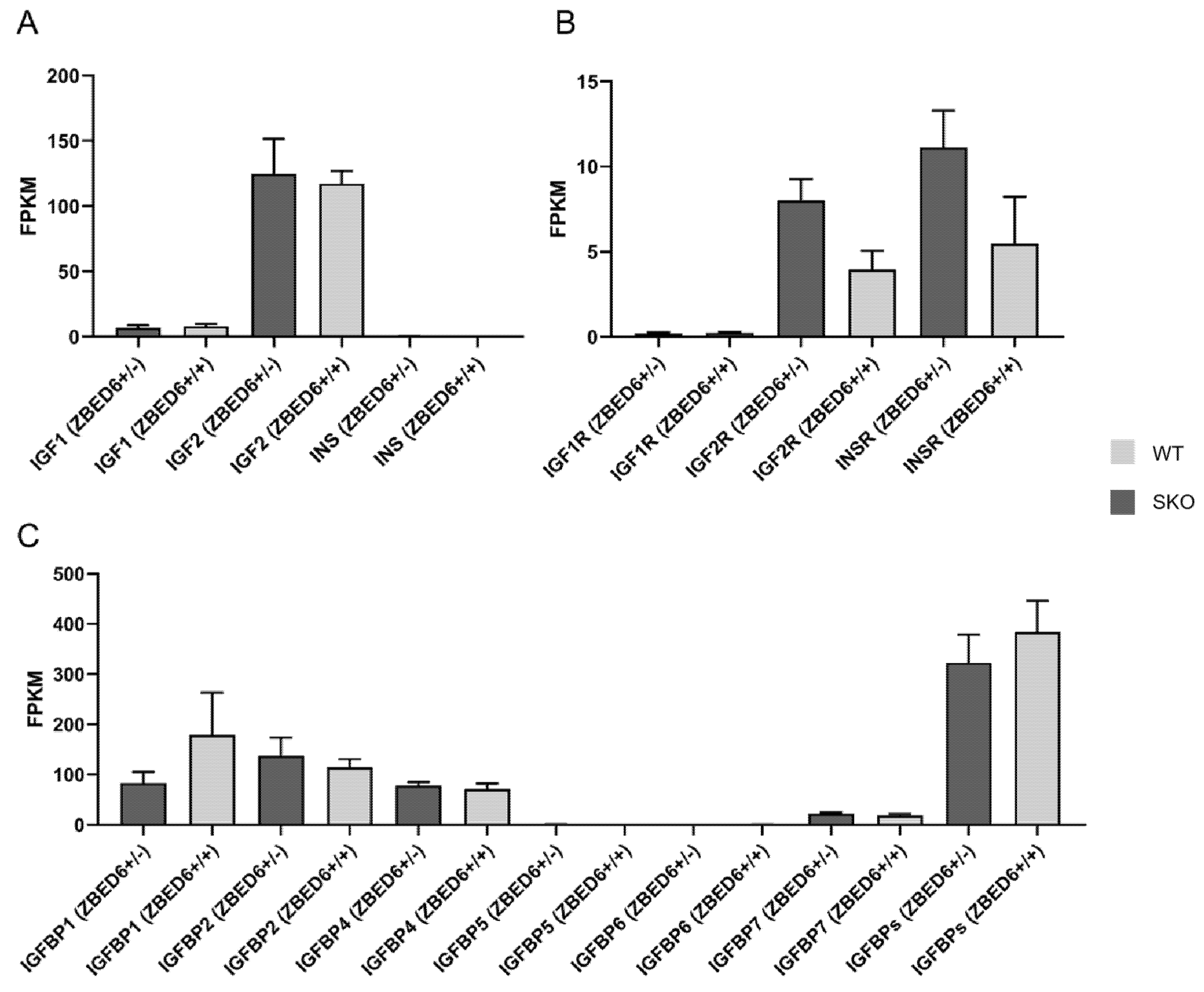
3.2. Expression of IGF2 and Related Genes in ZBED6-SKO Pig Tissues
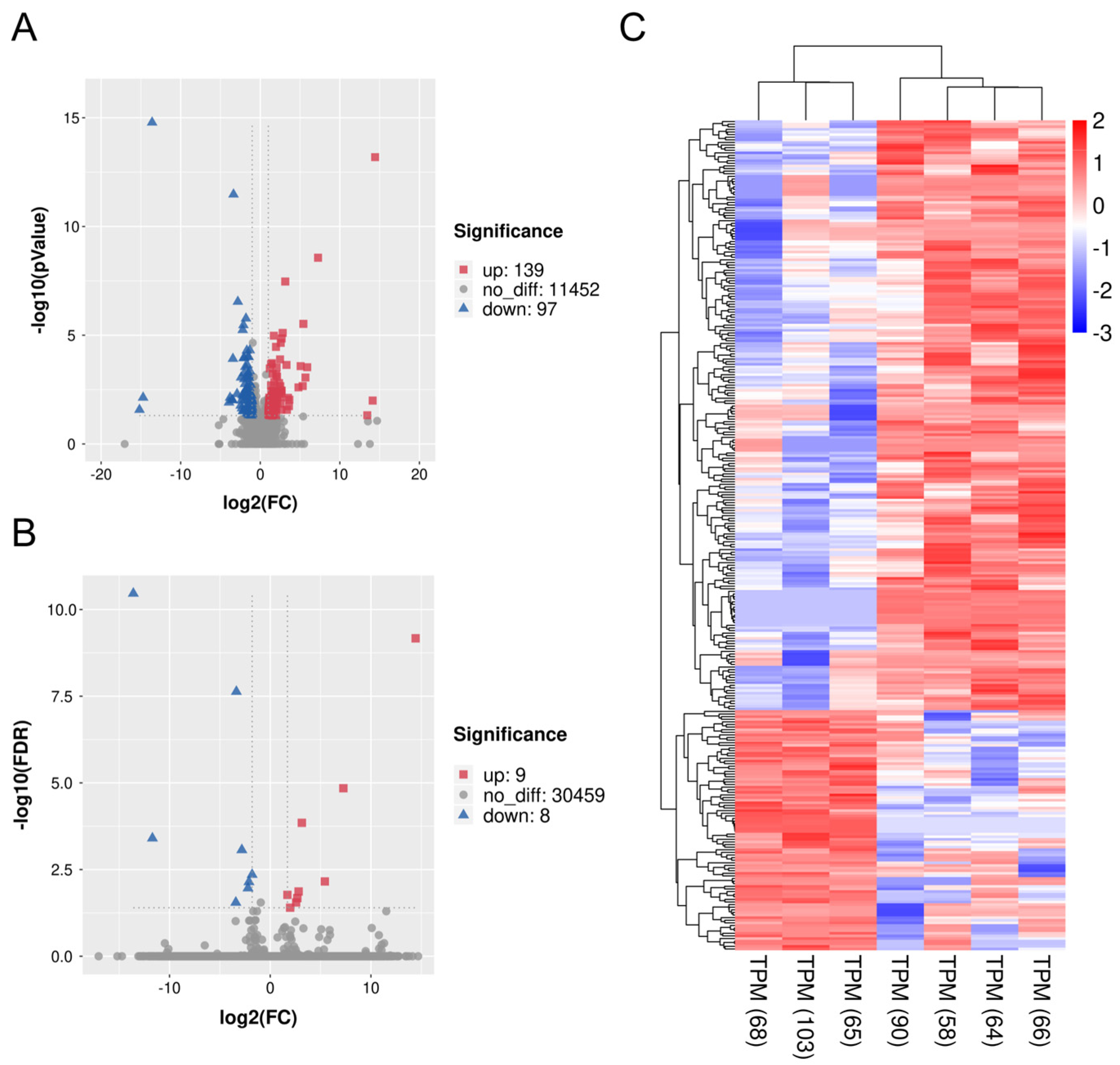
3.3. Quality Evaluation of Sequencing Data
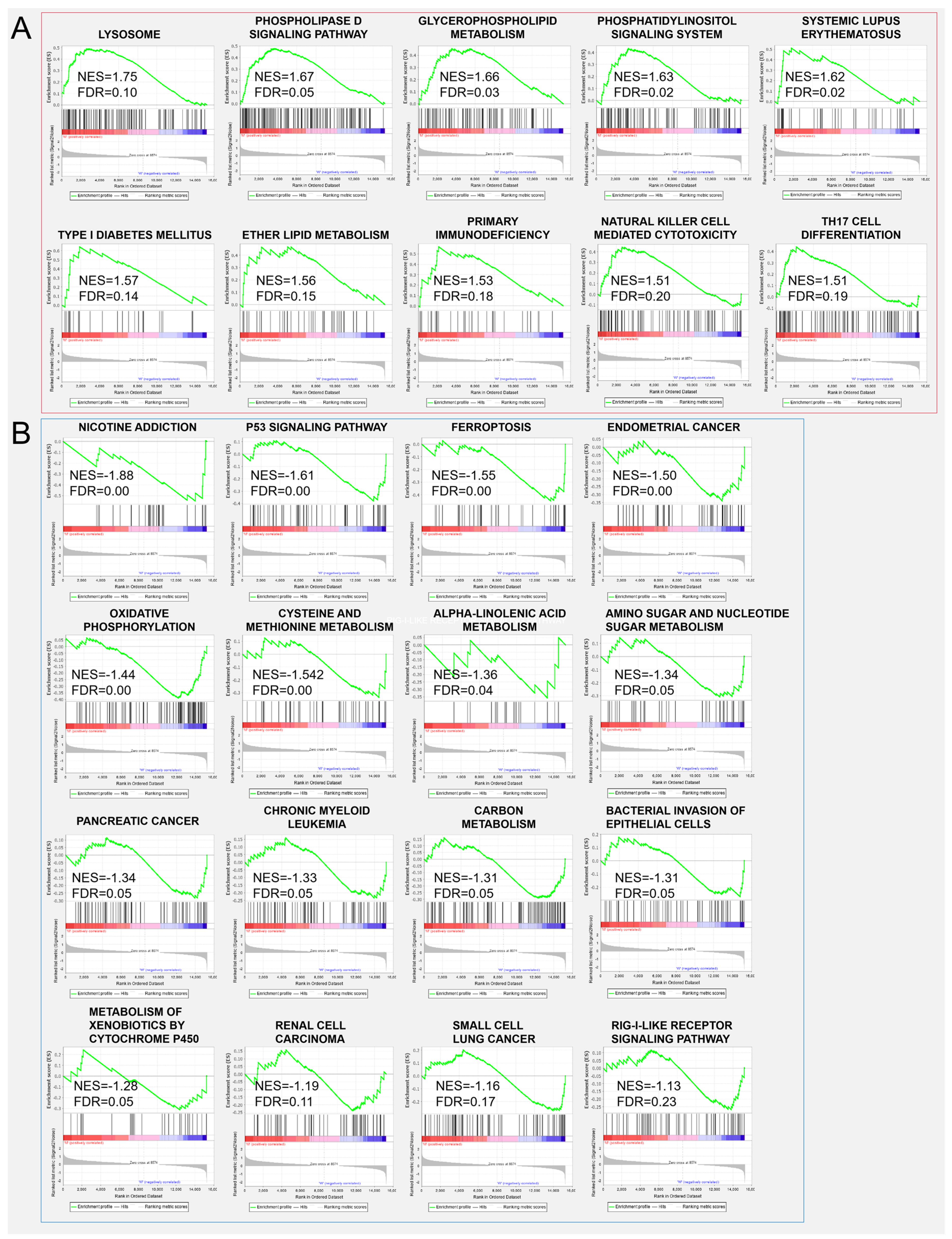
3.4. DEGs and Gene Enrichment in ZBED6-SKO Pig Livers
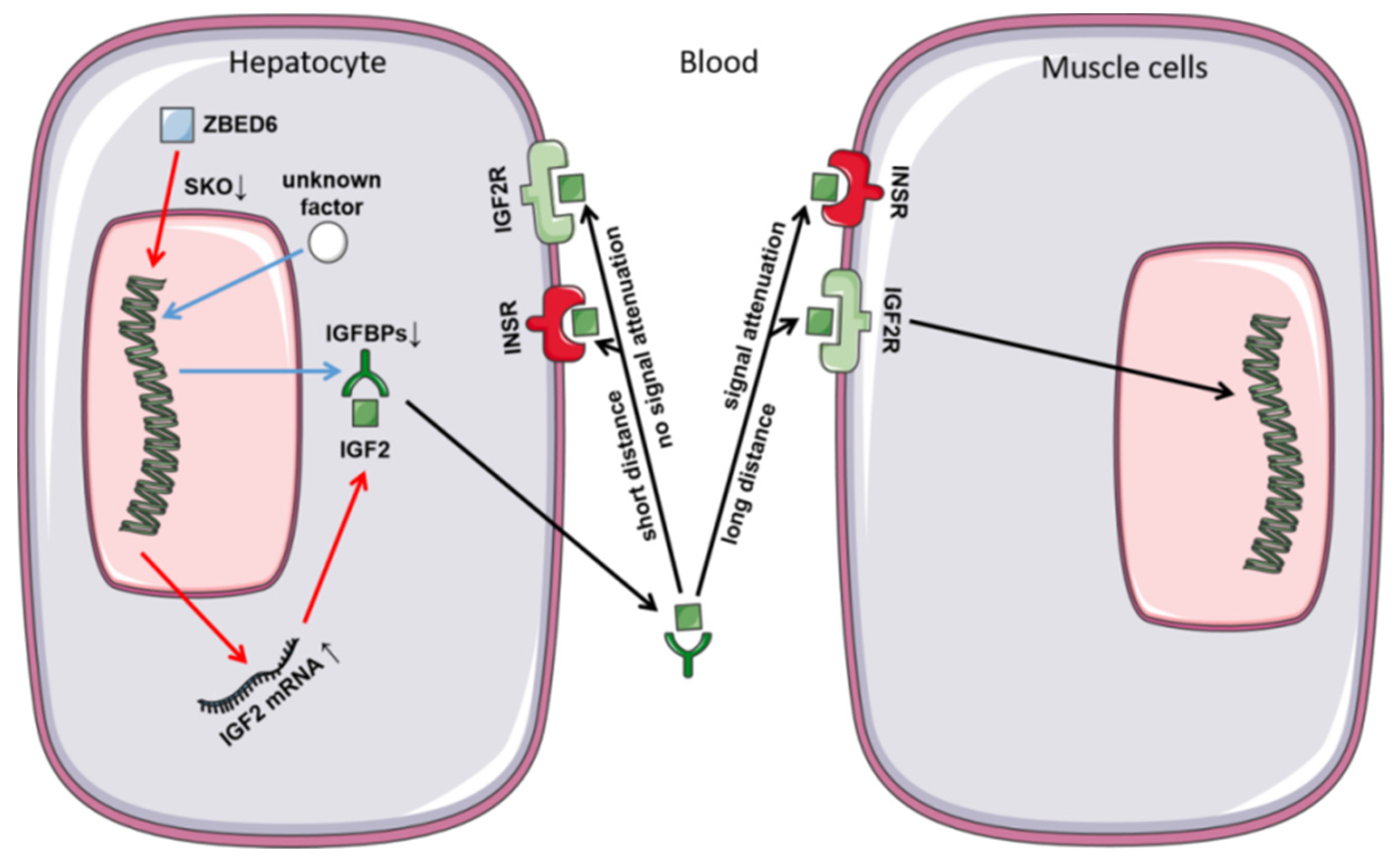
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Markljung, E.; Jiang, L.; Jaffe, J.D.; Mikkelsen, T.S.; Wallerman, O.; Larhammar, M.; Zhang, X.; Wang, L.; Saenz-Vash, V.; Gnirke, A.; et al. ZBED6, a novel transcription factor derived from a domesticated DNA transposon regulates IGF2 expression and muscle growth. PLoS Biol. 2009, 7, e1000256. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Jiang, L.; Wallerman, O.; Younis, S.; Yu, Q.; Klaesson, A.; Tengholm, A.; Welsh, N.; Andersson, L. ZBED6 negatively regulates insulin production, neuronal differentiation, and cell aggregation in MIN6 cells. FASEB J. 2019, 33, 88–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mourmouras, N.; Philippou, A.; Christopoulos, P.; Kostoglou, K.; Grivaki, C.; Konstantinidis, C.; Serafetinides, E.; Delakas, D.; Koutsilieris, M. Differential Expression of IGF-I Transcripts in Bladder Cancer. Anticancer Res. 2018, 38, 3453–3459. [Google Scholar] [CrossRef] [PubMed]
- Bella, P.; Farini, A.; Banfi, S.; Parolini, D.; Tonna, N.; Meregalli, M.; Belicchi, M.; Erratico, S.; D’Ursi, P.; Bianco, F.; et al. Blockade of IGF2R improves muscle regeneration and ameliorates Duchenne muscular dystrophy. EMBO Mol. Med. 2020, 12, e11019. [Google Scholar] [CrossRef] [PubMed]
- Garrett, S.M.; Hsu, E.; Thomas, J.M.; Pilewski, J.M.; Feghali-Bostwick, C. Insulin-like growth factor (IGF)-II- mediated fibrosis in pathogenic lung conditions. PLoS ONE 2019, 14, e0225422. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cao, X.; Dong, D.; Shen, X.; Cheng, J.; Jiang, R.; Yang, Z.; Peng, S.; Huang, Y.; Lan, X.; et al. Circular RNA TTN Acts As a miR-432 Sponge to Facilitate Proliferation and Differentiation of Myoblasts via the IGF2/PI3K/AKT Signaling Pathway. Mol. Ther.-Nucl. Acids 2019, 18, 966–980. [Google Scholar] [CrossRef] [Green Version]
- Younis, S.; Schönke, M.; Massart, J.; Hjortebjerg, R.; Sundström, E.; Gustafson, U.; Björnholm, M.; Krook, A.; Frystyk, J.; Zierath, J.R.; et al. The ZBED6-IGF2 axis has a major effect on growth of skeletal muscle and internal organs in placental mammals. Proc. Natl. Acad. Sci. USA 2018, 115, E2048–E2057. [Google Scholar] [CrossRef] [Green Version]
- Yi, T.; Wang, T.; Shi, Y.; Peng, X.; Tang, S.; Zhong, L.; Chen, Y.; Li, Y.; He, K.; Wang, M.; et al. Long noncoding RNA 91H overexpression contributes to the growth and metastasis of HCC by epigenetically positively regulating IGF2 expression. Liver Int. 2020, 40, 456–467. [Google Scholar] [CrossRef]
- Xiang, G.; Ren, J.; Hai, T.; Fu, R.; Yu, D.; Wang, J.; Li, W.; Wang, H.; Zhou, Q. Editing porcine IGF2 regulatory element improved meat production in Chinese Bama pigs. Cell Mol. Life Sci. 2018, 75, 4619–4628. [Google Scholar] [CrossRef]
- Van Laere, A.S.; Nguyen, M.; Braunschweig, M.; Nezer, C.; Collette, C.; Moreau, L.; Archibald, A.L.; Haley, C.S.; Buys, N.; Tally, M.; et al. A regulatory mutation in IGF2 causes a major QTL effect on muscle growth in the pig. Nature 2003, 425, 832–836. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, L.; Wallerman, O.; Engström, U.; Ameur, A.; Gupta, R.K.; Qi, Y.; Andersson, L.; Welsh, N. Transcription factor ZBED6 affects gene expression, proliferation, and cell death in pancreatic beta cells. Proc. Natl. Acad. Sci. USA 2013, 110, 15997–16002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Liu, H.; Wang, M.; Li, R.; Zeng, J.; Mo, D.; Cong, P.; Liu, X.; Chen, Y.; He, Z. Disruption of the ZBED6 binding site in intron 3 of IGF2 by CRISPR/Cas9 leads to enhanced muscle development in Liang Guang Small Spotted pigs. Transgenic Res. 2019, 28, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Younis, S.; Naboulsi, R.; Wang, X.; Cao, X.; Larsson, M.; Sargsyan, E.; Bergsten, P.; Welsh, N.; Andersson, L. The importance of the ZBED6-IGF2 axis for metabolic regulation in mouse myoblast cells. FASEB J. 2020. [Google Scholar] [CrossRef] [PubMed]
- Calkins, S.S.; Elledge, N.C.; Mueller, K.E.; Marek, S.M.; Couger, M.B.; Elshahed, M.S.; Youssef, N.H. Development of an RNA interference (RNAi) gene knockdown protocol in the anaerobic gut fungus Pecoramyces ruminantium strain C1A. PeerJ 2018, 6, e4276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Wang, Q.; Gong, J.; Du, J.; Zhang, Y.; Zhao, X. Yak IGF2 Promotes Fibroblast Proliferation Via Suppression of IGF1R and PI3KCG Expression. Genes 2018, 9, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livingstone, C. IGF2 and cancer. Endocr.-Relat. Cancer 2013, 20, R321–R339. [Google Scholar] [CrossRef] [Green Version]
- Frago, S.; Nicholls, R.D.; Strickland, M.; Hughes, J.; Williams, C.; Garner, L.; Surakhy, M.; Maclean, R.; Rezgui, D.; Prince, S.N.; et al. Functional evolution of IGF2:IGF2R domain 11 binding generates novel structural interactions and a specific IGF2 antagonist. Proc. Natl. Acad. Sci. USA 2016, 113, E2766–E2775. [Google Scholar] [CrossRef] [Green Version]
- Agnieszka, A.; Aldona, K. Insulin-Like Growth Factor (IGF) System in Liver Diseases. Int. J. Mol. Sci. 2018, 19, 1308. [Google Scholar] [CrossRef] [Green Version]
- Minchenko, D.O.; Tsymbal, D.O.; Davydov, V.V.; Minchenko, O.H. Expression of genes encoding IGF1, IGF2, and IGFBPs in blood of obese adolescents with insulin resistance. Endocr. Regul. 2019, 53, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Neuzillet, Y.; Chapeaublanc, E.; Krucker, C.; De Koning, L.; Lebret, T.; Radvanyi, F.; Bernard-Pierrot, I. IGF1R activation and the in vitro antiproliferative efficacy of IGF1R inhibitor are inversely correlated with IGFBP5 expression in bladder cancer. BMC Cancer 2017, 17, 636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraldi, A.; Biffi, A.; Lombardi, A.; Visigalli, I.; Pepe, S.; Settembre, C.; Nusco, E.; Auricchio, A.; Naldini, L.; Ballabio, A.; et al. SUMF1 enhances sulfatase activities in vivo in five sulfatase deficiencies. Biochem. J. 2007, 403, 305–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, K.; Ohashi, M.; Ohno, K.; Takeuchi, A.; Matsuoka, E.; Fujisato, K.; Minami, T.; Ito, S.; Okuda-Ashitaka, E. Involvement of NIPSNAP1, a neuropeptide nocistatin-interacting protein, in inflammatory pain. Mol. Pain 2016, 12, 1744806916637699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellis, K.L.; Zhou, Y.; Beshansky, J.R.; Ainehsazan, E.; Yang, Y.; Selker, H.P.; Huggins, G.S.; Cupples, L.A.; Peter, I. Genetic variation at glucose and insulin trait loci and response to glucose-insulin-potassium (GIK) therapy: The IMMEDIATE trial. Pharm. J. 2015, 15, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, Y.; Torii, T.; Tanoue, A.; Yamauchi, J. VCAM1 acts in parallel with CD69 and is required for the initiation of oligodendrocyte myelination. Nat. Commun. 2016, 7, 13478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomioka, M.; Toda, Y.; Mañucat, N.B.; Akatsu, H.; Fukumoto, M.; Kono, N.; Arai, H.; Kioka, N.; Ueda, K. Lysophosphatidylcholine export by human ABCA7. Biochimica et biophysica acta. BBA-Mol. Cell. Biol. 2017, 1862, 658–665. [Google Scholar] [CrossRef]
- Crawley, S.W.; Weck, M.L.; Grega-Larson, N.E.; Shifrin, D.A., Jr.; Tyska, M.J. ANKS4B Is Essential for Intermicrovillar Adhesion Complex Formation. Dev. Cell 2016, 36, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Hua, Y.; Ying, X.; Qian, Y.; Liu, H.; Lan, Y.; Xie, A.; Zhu, X. Physiological and pathological impact of AQP1 knockout in mice. Biosci. Rep. 2019, 39, BSR20182303. [Google Scholar] [CrossRef]
- Brown, S.G.; Miller, M.R.; Lishko, P.V.; Lester, D.H.; Publicover, S.J.; Barratt, C.; Martins Da Silva, S. Homozygous in-frame deletion in CATSPERE in a man producing spermatozoa with loss of CatSper function and compromised fertilizing capacity. Hum. Reprod. 2018, 33, 1812–1816. [Google Scholar] [CrossRef] [Green Version]
- Stacey, M.; Chang, G.W.; Sanos, S.L.; Chittenden, L.R.; Stubbs, L.; Gordon, S.; Lin, H.H. EMR4, a novel epidermal growth factor (EGF)-TM7 molecule up-regulated in activated mouse macrophages, binds to a putative cellular ligand on B lymphoma cell line A20. JBC 2002, 277, 29283–29293. [Google Scholar] [CrossRef] [Green Version]
- Li, H.D.; Chen, X.; Xu, J.J.; Du, X.S.; Yang, Y.; Li, J.; Yang, X.J.; Huang, H.M.; Li, X.F.; Wu, M.F.; et al. DNMT3b-mediated methylation of ZSWIM3 enhances inflammation in alcohol-induced liver injury via regulating TRAF2-mediated NF-κB pathway. Clin. Sci. 2020, 134, 1935–1956. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xie, B.; Qi, Y.; Wallerman, O.; Vasylovska, S.; Andersson, L.; Kozlova, E.N.; Welsh, N. Knock-down of ZBED6 in insulin-producing cells promotes N-cadherin junctions between beta-cells and neural crest stem cells in vitro. Sci. Rep. 2016, 6, 19006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avanzato, D.; Pupo, E.; Ducano, N.; Isella, C.; Bertalot, G.; Luise, C.; Pece, S.; Bruna, A.; Rueda, O.M.; Caldas, C.; et al. High USP6NL Levels in Breast Cancer Sustain Chronic AKT Phosphorylation and GLUT1 Stability Fueling Aerobic Glycolysis. Cancer Res. 2018, 78, 3432–3444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Huynh, W.; Skokan, T.D.; Lu, W.; Weiss, A.; Vale, R.D. CRACR2a is a calcium-activated dynein adaptor protein that regulates endocytic traffic. J. Cell Biol. 2019, 218, 1619–1633. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Stieglitz, K.A. Structural Insights for Drugs Developed for Phospholipase D Enzymes. Curr. Drug Discov. Technol. 2018, 15, 81–93. [Google Scholar] [CrossRef]
- Park, S.H.; Lee, J.E.; Lee, S.M.; Lee, J.; Seo, C.S.; Hwang, G.S.; Jung, J. An unbiased lipidomics approach identifies key lipid molecules as potential therapeutic targets of Dohongsamul-tang against non-alcoholic fatty liver diseases in a mouse model of obesity. J. Ethnopharmacol. 2020, 260, 112999. [Google Scholar] [CrossRef]
- Mironova, Y.A.; Lenk, G.M.; Lin, J.P.; Lee, S.J.; Twiss, J.L.; Vaccari, I.; Bolino, A.; Havton, L.A.; Min, S.H.; Abrams, C.S.; et al. PI(3,5)P2 biosynthesis regulates oligodendrocyte differentiation by intrinsic and extrinsic mechanisms. eLife 2016, 5, e13023. [Google Scholar] [CrossRef] [Green Version]
- Dean, J.M.; Lodhi, I.J. Structural and functional roles of ether lipids. Protein Cell 2018, 9, 196–206. [Google Scholar] [CrossRef]
- Brites, P.; Ferreira, A.S.; da Silva, T.F.; Sousa, V.F.; Malheiro, A.R.; Duran, M.; Waterham, H.R.; Baes, M.; Wanders, R.J. Alkyl-glycerol rescues plasmalogen levels and pathology of ether-phospholipid deficient mice. PLoS ONE 2011, 6, e28539. [Google Scholar] [CrossRef] [Green Version]
- Inouye, H.; Kirschner, D.A. Evolution of myelin ultrastructure and the major structural myelin proteins. Brain Res. 2016, 1641 Pt A, 43–63. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers Name | Transcript_id | Sequence | Notes |
|---|---|---|---|
| IGF11 | XM_021091138.1 | F: GCCCAAGGCTCAGAAGGAAG | qRT-PCR |
| R: GAGCAAAGGATCCTGCCAGT | |||
| IGF22 | XM_021080621.1 | F: GTGGCATCGTGGAAGAGTGC | qRT-PCR |
| R: CCAGGTGTCATAGCGGAAGAA | |||
| IGF1R3 | XM_005659775.3 | F: ACGAGTGGAGAAATCTGCGG | qRT-PCR |
| R: TGAGCTTGGGAAAGCGGTAG | |||
| IGF2R4 | NM_001244473.1 | F: AGAAAGAGGTGCCGTGCTAC | qRT-PCR |
| R: CCGGAGCGTGTCTATGTCTC | |||
| IGFBP15 | NM_001195105.1 | F: ACAGCAAACAGTGCGAGACG | qRT-PCR |
| R: TACTGATGGCATTTGGGGTCC | |||
| IGFBP25 | NM_214003.1 | F: CAACCTCAAACAGTGCAAGATGT | qRT-PCR |
| R: GCTGTGGTTTACTGCATCCG | |||
| IGFBP35 | NM_001005156.1 | F: AGACGGAATACGGGCCTTGC | qRT-PCR |
| R: TCTTGTCGCAGTTGGGGATG | |||
| IGFBP45 | NM_001123129.1 | F: CAGCCCTCTGACAAGGACGA | qRT-PCR |
| R: GCTCCGGTCTCGGATCTTG | |||
| IGFBP55 | NM_214099.1 | F: CGAGCAAGCCAAGATCGAGAG | qRT-PCR |
| R: AGCTTCTTTCTGCGGTCCTT | |||
| IGFBP65 | NM_001100190.1 | F: CGCCCTCGGGGGAGAATC | qRT-PCR |
| R: ATCTCAGTGTCTTGGACGCC | |||
| IGFBP75 | XM_003129037.5 | F: TAAGCGGTGTGTGCGTGT | qRT-PCR |
| R: ATGGAAGGACCTTGCTCGC | |||
| ZBED6-16 | NM_001166552.1 | F: GCTTTGTTAGCGTCTGATGCC | qRT-PCR for ZBED6-KO 8 and ZBED6-WT 9 |
| R: AGGTTACAAATTGCCCGCCA | |||
| ZBED6-26 | NM_001166552.1 | F: GTACTAGAGCAAAGACTTCCATT | qRT-PCR for ZBED6-WT |
| R:ACAAATTGCCCGCCAGGTAT | |||
| ACTB7 | XM_021086047.1 | F: GGACTTCGAGCAGGAGATGG | qRT-PCR |
| R: AGGAAGGAGGGCTGGAAGAG |
| Type | ZBED6 SKO Group | Wild Group | ||
|---|---|---|---|---|
| Total Reads Count (#) | No.66 | 54877418 | No.65 | 48523032 |
| No.90 | 42383082 | No.68 | 44928968 | |
| No.64 | 48377850 | No.103 | 55037976 | |
| No.58 | 54016486 | |||
| Total Bases Count (bp) | No.66 | 7856669548 | No.65 | 6866998313 |
| No.90 | 6069354838 | No.68 | 6387348025 | |
| No.64 | 6912265759 | No.103 | 7908177595 | |
| No.58 | 7696200796 | |||
| Average Read Length (bp) | No.66 | 143.17 | No.65 | 141.52 |
| No.90 | 143.20 | No.68 | 142.17 | |
| No.64 | 142.88 | No.103 | 143.69 | |
| No.58 | 142.48 | |||
| Q20 Bases Ratio (%) | No.66 | 98.83% | No.65 | 98.79% |
| No.90 | 98.82% | No.68 | 98.70% | |
| No.64 | 98.83% | No.103 | 98.83% | |
| No.58 | 98.79% | |||
| Q30 Bases Ratio (%) | No.66 | 95.61% | No.65 | 95.51% |
| No.90 | 95.56% | No.68 | 95.27% | |
| No.64 | 95.64% | No.103 | 95.60% | |
| No.58 | 95.52% | |||
| GC Bases Ratio (%) | No.66 | 50.05% | No.65 | 49.21% |
| No.90 | 48.97% | No.68 | 50.11% | |
| No.64 | 50.33% | No.103 | 49.26% | |
| No.58 | 49.61% | |||
| Gene ID | Gene Name | MeanTPM 6 (SKO) | MeanTPM 6 (WT 7) | log2 (FC) | p Value | FDR Value | Notes |
|---|---|---|---|---|---|---|---|
| ENSSSCG00000049098 | - | 0.00010000 | 1.23588067 | −13.59325183 | 1.65 × 10−15 | 3.45 × 10−11 | LncRNA |
| ENSSSCG00000044627 | - | 2.20908400 | 0.00010000 | 14.43116066 | 6.46 × 10−14 | 6.76 × 10−10 | LncRNA |
| ENSSSCG00000046118 | - | 0.13868175 | 1.42510900 | −3.361222417 | 3.35 × 10−12 | 2.34 × 10−8 | Pseudogene |
| ENSSSCG00000003986 | ZFP211L | 1.90845525 | 0.01244633 | 7.260540758 | 2.74 × 10−9 | 1.43 × 10−5 | Protein coding |
| ENSSSCG00000011532 | SUMF1 | 18.72840650 | 2.12422033 | 3.140222832 | 3.39 × 10−8 | 1.42 × 10−4 | Protein coding |
| ENSSSCG00000051359 | NIPSNAP1 | 0.00010000 | 0.33187667 | −11.69643149 | 1.13 × 10−7 | 3.96 × 10−4 | Protein coding |
| ENSSSCG00000011114 | SEC61A2 | 1.51700350 | 10.70244267 | −2.818643788 | 2.87 × 10−7 | 8.57 × 10−4 | Protein coding |
| ENSSSCG00000006862 | VCAM1 | 7.08961450 | 24.63916067 | −1.797174024 | 1.70 × 10−6 | 4.43 × 10−3 | Protein coding |
| ENSSSCG00000023121 | ABCA7 | 1.06993575 | 0.02500733 | 5.419029131 | 3.01 × 10−6 | 6.99 × 10−3 | Protein coding |
| ENSSSCG00000007850 | ANKS4B | 9.24855825 | 39.92152333 | −2.109866385 | 3.43 × 10−6 | 7.18 × 10−3 | Protein coding |
| ENSSSCG00000033190 | AQP1 | 0.99121175 | 4.57049600 | −2.205085543 | 5.73 × 10−6 | 1.09 × 10−2 | Protein coding |
| ENSSSCG00000000732 | CRACR2A | 1.02969150 | 0.14599400 | 2.81823118 | 7.75 × 10−6 | 1.35 × 10−2 | Protein coding |
| ENSSSCG00000011120 | USP6NL | 3.51777325 | 1.07724900 | 1.707310733 | 1.05 × 10−5 | 1.70 × 10−2 | Protein coding |
| ENSSSCG00000037808 | novel gene | 417.64328400 | 64.70200867 | 2.690388832 | 1.41 × 10−5 | 2.11 × 10−2 | Protein coding |
| ENSSSCG00000035850 | CATSPERE | 0.07047350 | 0.75291833 | −3.417340618 | 2.29 × 10−5 | 2.81 × 10−2 | Protein coding |
| ENSSSCG00000040016 | EMR4 | 12.88731075 | 2.17047567 | 2.569868088 | 2.26 × 10−5 | 2.81 × 10−2 | Protein coding |
| ENSSSCG00000007427 | ZSWIM3 | 1.82821425 | 0.46398233 | 1.978293372 | 3.43 × 10−5 | 3.98 × 10−2 | Protein coding |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.; Wu, M.; Liu, S.; Tang, X.; Yi, X.; Li, Q.; Wang, S.; Sun, X. Liver Expression of IGF2 and Related Proteins in ZBED6 Gene-Edited Pig by RNA-Seq. Animals 2020, 10, 2184. https://doi.org/10.3390/ani10112184
Zhao H, Wu M, Liu S, Tang X, Yi X, Li Q, Wang S, Sun X. Liver Expression of IGF2 and Related Proteins in ZBED6 Gene-Edited Pig by RNA-Seq. Animals. 2020; 10(11):2184. https://doi.org/10.3390/ani10112184
Chicago/Turabian StyleZhao, Haidong, Mingli Wu, Shirong Liu, Xiaoqin Tang, Xiaohua Yi, Qi Li, Shuhui Wang, and Xiuzhu Sun. 2020. "Liver Expression of IGF2 and Related Proteins in ZBED6 Gene-Edited Pig by RNA-Seq" Animals 10, no. 11: 2184. https://doi.org/10.3390/ani10112184
APA StyleZhao, H., Wu, M., Liu, S., Tang, X., Yi, X., Li, Q., Wang, S., & Sun, X. (2020). Liver Expression of IGF2 and Related Proteins in ZBED6 Gene-Edited Pig by RNA-Seq. Animals, 10(11), 2184. https://doi.org/10.3390/ani10112184