Molecular Identification of the “Facciuta Della Valnerina” Local Goat Population Reared in the Umbria Region, Italy

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Sampling
2.2. Molecular Analyses
2.3. Statistical Analysis
3. Results and Discussion
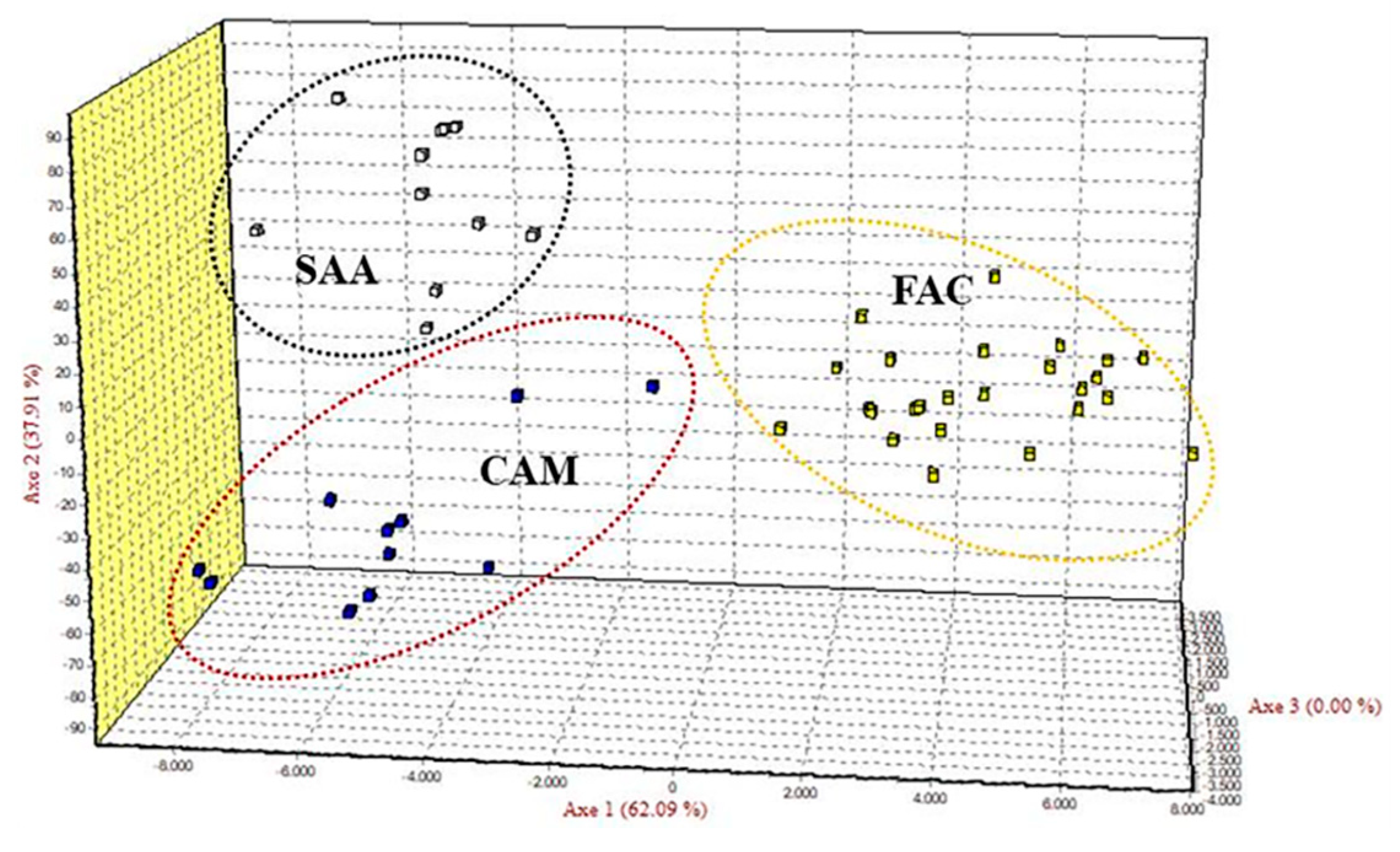
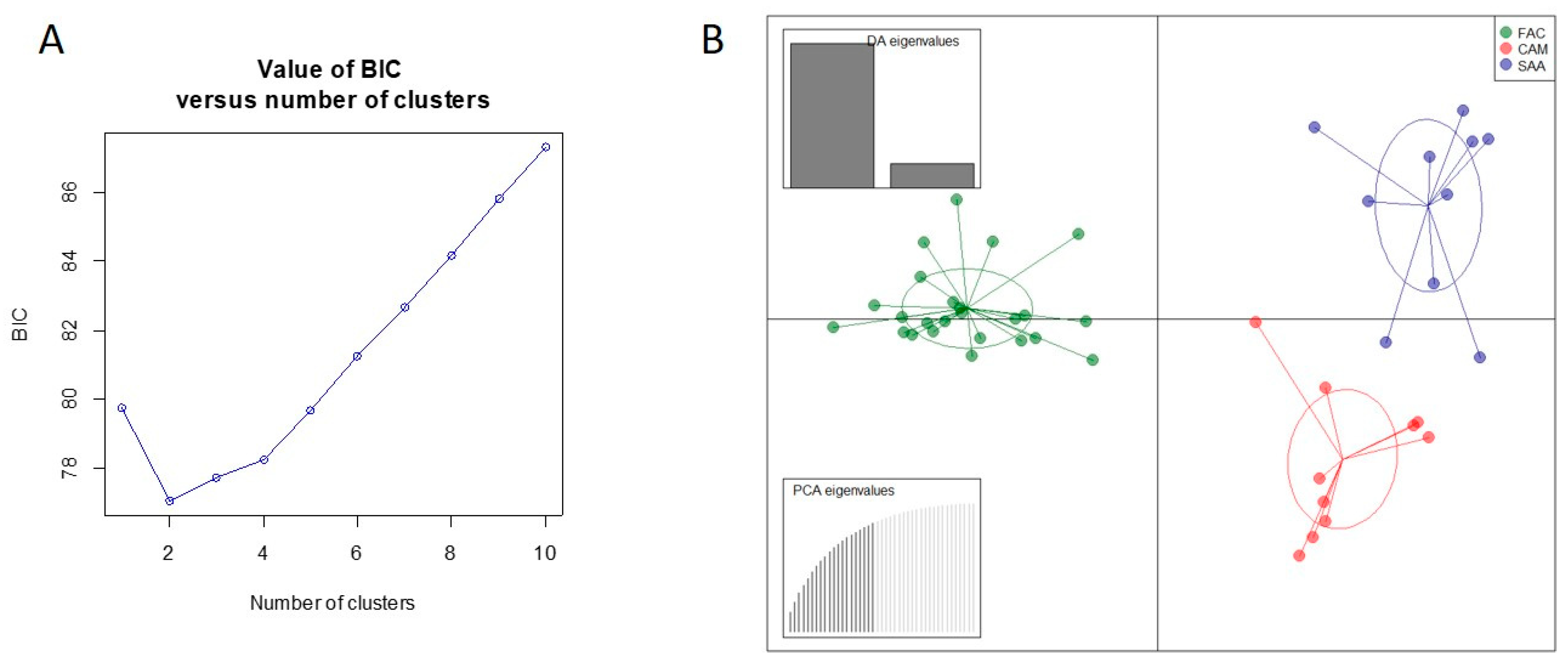
3.1. Genetic Variation
3.2. Genetic Differentiation
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Miller, B.A.; Lu, C.D. Current status of global dairy goat production: An overview. Asian-Australas. J. Anim. Sci. 2019, 32, 1219–1232. [Google Scholar] [CrossRef] [PubMed]
- Aziz, M.A. Present status of the world goat populations and their productivity. Lohmann. Inf. 2010, 45, 42–52. [Google Scholar]
- Hersleth, M.; Næs, T.; Rødbotten, M.; Lind, V.; Monteleone, E. Lamb meat—Importance of origin and grazing system for Italian and Norwegian consumers. Meat Sci. 2012, 90, 899–907. [Google Scholar] [CrossRef]
- Global Plan of Action for Animal Genetic Resources and the Interlaken Declaration; FAO Document Repository: Rome, Italy, 2007.
- Webb, E.C.; Casey, N.H.; Simela, L. Goat meat quality. Small Rumin. 2005, 60, 153–166. [Google Scholar] [CrossRef]
- Montossi, F.; Font-i-Furnols, M.; Del Campo, M.; San Julián, R.; Brito, G.; Sanudo, C. Sustainable sheep production and consumer preference trends compatibilities, contradictions, and unresolved dilemmas. Meat Sci. 2013, 95, 772–789. [Google Scholar] [CrossRef] [PubMed]
- Di Stasio, L.; Piatti, P.; Fontanella, E.; Costa, S.; Bigi, D.; Lasagna, E.; Pauciullo, A. Lamb meat traceability: The case of Sambucana sheep. Small Rumin. Res. 2017, 149, 85–90. [Google Scholar] [CrossRef]
- Ciampolini, R.; Leveziel, H.; Mozzanti, E.; Grohs, C.; Cianci, D. Genomic identification of an individual or its tissue. Meat Sci. 2000, 54, 35–40. [Google Scholar] [CrossRef]
- Pizzuti, T.; Mirabelli, G.; Grasso, G.; Paldino, G. MESCO (MEat Supply Chain Ontology): An ontology for supporting traceability in the meat supply chain. Food Control 2017, 72, 123–133. [Google Scholar] [CrossRef]
- Biswas, A.K.; Mandal, P.K. Current perspectives of meat quality evaluation: Techniques, technologies, and challenges. Meat Qual. Anal. 2020, 3–17. [Google Scholar] [CrossRef]
- Orrú, L.; Napolitano, F.; Catillo, G.; Moioli, B. Meat molecular traceability: How to choose the best set of microsatellites? Meat Sci. 2006, 72, 312–317. [Google Scholar] [CrossRef]
- Jobling, M.A.; Gill, P. Encoded evidence: DNA in forensic analysis. Nat. Rev. Genet. 2004, 10, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Dalvit, C.; De Marchi, M.; Cassandro, M. Genetic traceability of livestock products: A review. Meat Sci. 2007, 77, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Negrini, R.; Nicoloso, L.; Crepaldi, P.; Milanesi, E.; Marino, R.; Perini, D.; Pariset, L.; Dunner, S.; Leveziel, H.; Williams, J.L.; et al. Traceability of four European protected geographic indication (PGI) beef products using single nucleotide polymorphisms (SNP) and Bayesian statistics. Meat Sci. 2008, 80, 1212–1217. [Google Scholar] [CrossRef] [PubMed]
- Martins-Lopes, P.; Gomes, S.; Pereira, L.; Guedes-Pinto, H. Molecular markers for food traceability. Food Technol. Biotechnol. 2013, 51, 198–207. [Google Scholar]
- Zhao, J.; Zhu, C.; Xu, Z.; Jiang, X.; Yang, S.; Chen, A. Microsatellite markers for animal identification and meat traceability of six beef cattle breeds in the Chinese market. Food Control 2017, 78, 469–475. [Google Scholar] [CrossRef]
- Dalvit, C.; De Marchi, M.; Targhetta, C.; Gervaso, M.; Cassandro, M. Genetic traceability of meat using microsatellite markers. Food Res. Int. 2008, 41, 301–307. [Google Scholar] [CrossRef]
- Cao, J.; Li, X.; Du, X.; Zhao, S. Microsatellite based genetic diversity and population structure of nine indigenous Chinese domestic goats. Small Rumin. Res. 2017, 148, 80–86. [Google Scholar] [CrossRef]
- Selepe, M.M.; Ceccobelli, S.; Lasagna, E.; Kunene, N.W. Genetic structure of South African Nguni (Zulu) sheep populations reveals admixture with exotic breeds. PLoS ONE 2018, 13, e0196276. [Google Scholar] [CrossRef] [Green Version]
- Tefiel, H.; Ata, N.; Chabbar, M.; Benyarou, M.; Fantazi, K.; Yilmaz, O.; Cemal, I.; Karaca, O.; Boudouma, D.; Gaouar, S.B.S. Genetic characterization of four Algerian goat breeds assessed by microsatellite markers. Small Rumin. Res. 2018, 160, 65–71. [Google Scholar] [CrossRef]
- Secondary Guidelines for Development of National Farm Animal Genetic Resources Management Plans. Measurements of Domestic Animal Diversity (MoDAD): Recommended Microsatellite Markers; FAO/ISAG Document: Rome, Italy, 2004.
- Murital, I.; Afolayan, O.; Bemji, M.N.; Dadi, O.; Landi, V.; Martínez, A.; Delgado, J.V.; Aina, A.B.J.; Adebambo, A.O. Genetic diversity and population structure of Nigerian indigenous goat using DNA microsatellite markers. Archivos de zootecnia 2015, 64, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Park, S.D.E. Trypanotolerance in West African Cattle and the Population Genetic Effects of Selection. Ph.D. Thesis, Dublin University, Dublin, Ireland, 2001. [Google Scholar]
- Kalinowski, S.T. HP-RARE 1.0: A computer program for performing rarefaction on measures of allelic richness. Mol. Ecol. Notes 2005, 5, 187–189. [Google Scholar] [CrossRef]
- Rousset, F. GENEPOP’007: A complete re-implementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358–1370. [Google Scholar]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetic analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Lewis, P.O.; Zaykin, D. Genetic data analysis. In Computer Program for the Analysis of Allelic Data; Version 1.0.; Lewis Labs, Univ of Connecticut: Storrs, CT, USA, 1999. [Google Scholar]
- Benzécri, J.P. L’analyse des données Volume II. L’analyse des correspondances; Dunod: Paris, France, 1973. [Google Scholar]
- Jombart, T. Adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2017. [Google Scholar]
- Criscione, A.; Marletta, D.; Ådnøy, T.; Bordonaro, S.; Guastella, A.M.; Lien, S.; D’Urso, G. Characterization of biodiversity in six goat breeds reared in Southern Italy by means of microsatellite and SNP markers. Ital. J. Anim. 2007, 6 (Suppl. 1), 95–97. [Google Scholar] [CrossRef]
- Bruno-de-Sousa, C.; Martinez, A.M.; Ginja, C.L.; Santos-Silva, F.; Carolino, M.I.; Delgado, J.V.; Gama, L.T. Genetic diversity and population structure in Potuguese goat breeds. Livest. Sci. 2011, 135, 131–139. [Google Scholar] [CrossRef]
- Hoda, A. Genetic diversity of the Capore goat in Albania based on 30 microsatellite markers. Maced. J. Anim. Sci. 2011, 1, 53–56. [Google Scholar]
- Aljumaah, R.S.; Musthafa, M.M.; Al-Shaikh, M.A.; Badri, O.M.; Hussein, M.F. Genetic diversity of Ardi goat based on microsatellite analysis. Afr. J. Biotechnol. 2012, 11, 16539–16545. [Google Scholar]
- Asroush, F.; Mirhoseini, S.Z.; Badbarin, N.; Seidavi, A.; Tufarelli, V.; Laudadio, V.; Dario, C.; Selvaggi, M. Genetic characterization of Markhoz goat breed using microsatellite markers. Arch. Anim. Breed. 2018, 61, 469–473. [Google Scholar] [CrossRef]
- Guang-Xin, E.; Hong, Q.; Zhao, Y.; Ma, Y.; Chu, M.; Zhu, L.; Huang, Y. Genetic diversity estimation of Yunnan indigenous goat breeds using microsatellite markers. Ecol. Evol. 2019, 9, 5916–5924. [Google Scholar]
- Hussain, T.; Shaheen, M.; Barbar, M.; Musthafa, M.; Nadeem, A.; Nawaz, A.; Javed, M.; Marikar, F. Molecular diversity analysis of Jattal and Dera Din Panah goat breeds of Pakistan using microsatellite markers. J. Hell. Vet. Med. Soc. 2018, 69, 791–796. [Google Scholar] [CrossRef] [Green Version]
- Nguluma, A.S.; Huang, Y.; Zhao, Y.; Chen, L.; Msalya, G.; Lyimo, C.; Guangxin, E.; Chenyambuga, S.W. Assessment of genetic variation among four populations of Small East African goats using microsatellite markers. S. Afr. J. Anim. Sci. 2018, 48, 117–127. [Google Scholar] [CrossRef] [Green Version]
- Iamartino, D.; Bruzzone, A.; Lanza, A.; Blasi, M.; Pilla, F. Genetic diversity of Southern Italian goat populations assessed by microsatellite markers. Small Rumin. Res. 2005, 57, 249–255. [Google Scholar] [CrossRef]
- Negrini, R.; D’Andrea, M.; Crepaldi, P.; Colli, L.; Nicoloso, L.; Guastella, A.M.; Sechi, T.; Bordonaro, S.; Ajmone-Marsan, P.; Pilla, F. Econogene Consortium. Effect of microsatellite outliers on the genetic structure of eight Italian goat breeds. Small Rumin. Res. 2012, 103, 99–107. [Google Scholar] [CrossRef]
- Sardina, M.T.; Tortorici, L.; Mastrangelo, S.; Di Gerlando, R.; Tolone, M.; Portolano, B. Application of microsatellite markers as potential tools for traceability of Girgentana goat breed dairy products. Food Res. Int. 2015, 74, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Ojo, O.A.; Akpa, G.N.; Orunmuyi, M.; Adeyinka, I.A.; Kabir, M.; Alphonsus, C. Genetic analysis of Nigerian indigenous goat populations using microsatellite markers. Iran. J. Appl. Anim. Sci. 2018, 8, 287–294. [Google Scholar]
- Takahashi, H.; Nyamsamba, D.; Mandakh, B.; Zagdsuren, Y.; Amano, T.; Nomura, K.; Yokohama, M.; Ito, S.; Minezawa, M. Genetic structure of Mongolian goat populations using microsatellite loci analysis. Asian-Australas. J. Anim. Sci. 2008, 21, 947–953. [Google Scholar] [CrossRef]
- Oliveira, J.D.; Igarashi, M.L.S.P.; Machado, T.M.M.; Miretti, M.M.; Ferro, J.A.; Contel, E.P.B. Structure and genetic relationships between Brazilian naturalized and exotic purebreed goat domestic goat (Capra hircus) breeds based on microsatellites. Genet. Mol. Biol. 2007, 30, 356–363. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Locus | Chr. | S.R. (bp) | Na | HE | HO | PIC | HWE Breed † |
|---|---|---|---|---|---|---|---|
| INRA005 | 10 | 176–190 | 5 | 0.59 | 0.54 | 0.51 | 0 |
| BM8125 | 17 | 110–130 | 9 | 0.71 | 0.63 | 0.63 | 1 |
| CSRD247 | 14 | 220–247 | 8 | 0.65 | 0.57 | 0.59 | 1 |
| HAUT27 | 26 | 128–158 | 7 | 0.77 | 0.82 | 0.71 | 0 |
| TGLA122 | 21 | 137–181 | 8 | 0.75 | 0.78 | 0.68 | 0 |
| HSC | 20 | 267–301 | 13 | 0.86 | 0.78 | 0.80 | 0 |
| MCM527 | 5 | 165–187 | 7 | 0.65 | 0.72 | 0.60 | 0 |
| SRCRSP8 | Not reported | 215–255 | 9 | 0.52 | 0.56 | 0.50 | 0 |
| BM1329 | 6 | 155–200 | 6 | 0.66 | 0.50 | 0.58 | 1 |
| OarFCB11 | 2 | 122–140 | 7 | 0.75 | 0.71 | 0.70 | 2 |
| MAF209 | 17 | 100–104 | 4 | 0.20 | 0.19 | 0.18 | 2 |
| MAF65 | 15 | 116–158 | 10 | 0.75 | 0.52 | 0.68 | 1 |
| CRSM60 | Not reported | 75–91 | 6 | 0.72 | 0.43 | 0.66 | 3 |
| ETH10 | 5 | 212–224 | 4 | 0.46 | 0.44 | 0.50 | 0 |
| ILSTS019 | Not reported | 142–162 | 6 | 0.78 | 0.78 | 0.72 | 2 |
| SRCRSP5 | 21 | 156–178 | 7 | 0.64 | 0.76 | 0.57 | 0 |
| Total (±SD) | 116 ± 2.29 | 0.65 ± 0.16 | 0.61 ± 0.17 | 0.60 ± 0.15 |
| Population/Breed | N | MNA ± SD | Rt (1) | PA | HO ± SD | HE ± SD |
|---|---|---|---|---|---|---|
| FAC | 24 | 6.67 ± 2.10 | 5.17 | 25 | 0.68 ± 0.03 | 0.74 ± 0.03 |
| CAM | 10 | 4.58 ± 1.62 | 4.36 | 4 | 0.59 ± 0.05 | 0.63 ± 0.06 |
| SAA | 10 | 4.92 ± 1.38 | 4.56 | 5 | 0.64 ± 0.04 | 0.64 ± 0.04 |
| Locus | Population/Breed | ||
|---|---|---|---|
| FAC | CAM | SAA | |
| INRA5 | 113 (0.1000) | ||
| BM8125 | 109 (0.0217) | 123 (0.0500) | 119 (0.0500) |
| 121 (0.0217) | |||
| 127 (0.0217) | |||
| CSRD247 | 216 (0.1304) | 228 (0.3125) | 228 (0.1111) |
| 232 (0.2174) | 234 (0.1875) | 242 (0.1250) | |
| HAUT27 | 145 (0.0500) | 145 (0.1000) | |
| TGLA122 | 147 (0.0455) | 133 (0.1000) | |
| HSC | 268 (0.0217) | 266 (0.0500) | |
| 276 (0.0435) | 270 (0.0500) | 270 (0.2000) | |
| 278 (0.0435) | |||
| 296 (0.0217) | |||
| MCM527 | 160 (0.1304) | ||
| SRCRSP8 | 218 (0.0217) | 224 (0.0500) | |
| 230 (0.0217) | 242 (0.1000) | ||
| 238 (0.0435) | |||
| BM1329 | 174 (0.1087) | ||
| 180 (0.0870) | |||
| MAF65 | 117 (0.0870) | ||
| 119 (0.0435) | |||
| 125 (0.1957) | |||
| 127 (0.0435) | |||
| 129 (0.2391) | |||
| MAF209 | 105 (0.7708) | 101 (0.0500) | 101 (0.0500) |
| 107 (0.1042) | |||
| SRCRSP5 | 161 (0.1250) | ||
| 179 (0.0313) | |||
| Population/breed | N | FAC | CAM | SAA |
|---|---|---|---|---|
| FAC | 24 | 0.0000 | ||
| CAM | 10 | 0.0897 (0.038–0.131) | 0.0000 | |
| SAA | 10 | 0.0928 (0.060–0.109) | 0.0729 (0.042–0.141) | 0.0000 |
| Global FST = 0.084 (0.061–0.113) | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceccobelli, S.; Lasagna, E.; Demir, E.; Rovelli, G.; Albertini, E.; Veronesi, F.; Sarti, F.M.; Rosellini, D. Molecular Identification of the “Facciuta Della Valnerina” Local Goat Population Reared in the Umbria Region, Italy. Animals 2020, 10, 601. https://doi.org/10.3390/ani10040601
Ceccobelli S, Lasagna E, Demir E, Rovelli G, Albertini E, Veronesi F, Sarti FM, Rosellini D. Molecular Identification of the “Facciuta Della Valnerina” Local Goat Population Reared in the Umbria Region, Italy. Animals. 2020; 10(4):601. https://doi.org/10.3390/ani10040601
Chicago/Turabian StyleCeccobelli, Simone, Emiliano Lasagna, Eymen Demir, Giacomo Rovelli, Emidio Albertini, Fabio Veronesi, Francesca Maria Sarti, and Daniele Rosellini. 2020. "Molecular Identification of the “Facciuta Della Valnerina” Local Goat Population Reared in the Umbria Region, Italy" Animals 10, no. 4: 601. https://doi.org/10.3390/ani10040601
APA StyleCeccobelli, S., Lasagna, E., Demir, E., Rovelli, G., Albertini, E., Veronesi, F., Sarti, F. M., & Rosellini, D. (2020). Molecular Identification of the “Facciuta Della Valnerina” Local Goat Population Reared in the Umbria Region, Italy. Animals, 10(4), 601. https://doi.org/10.3390/ani10040601