Ultrastructural Characterization of Porcine Growing and In Vitro Matured Oocytes

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Ovaries and Oocyte In Vitro Maturation (IVM)
2.2. Light Microscopic Evaluation
2.3. Transmission Electron Microscopy (TEM)
2.4. Morphometric Analysis of Mitochondria
3. Results
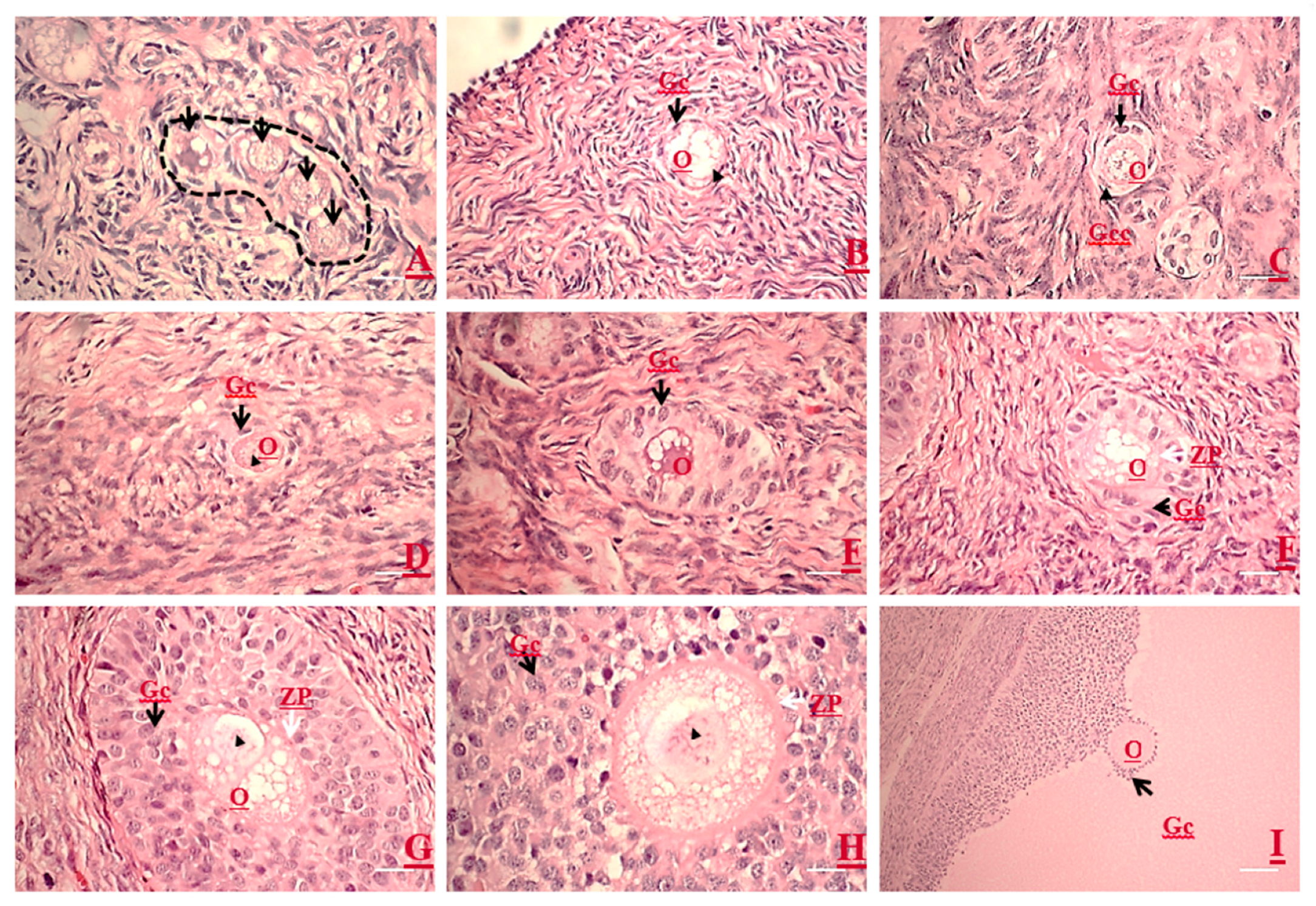
3.1. Light Microscopic Structures Examined by HE Staining
3.2. Ultrastructures of Follicles Examined by TEM
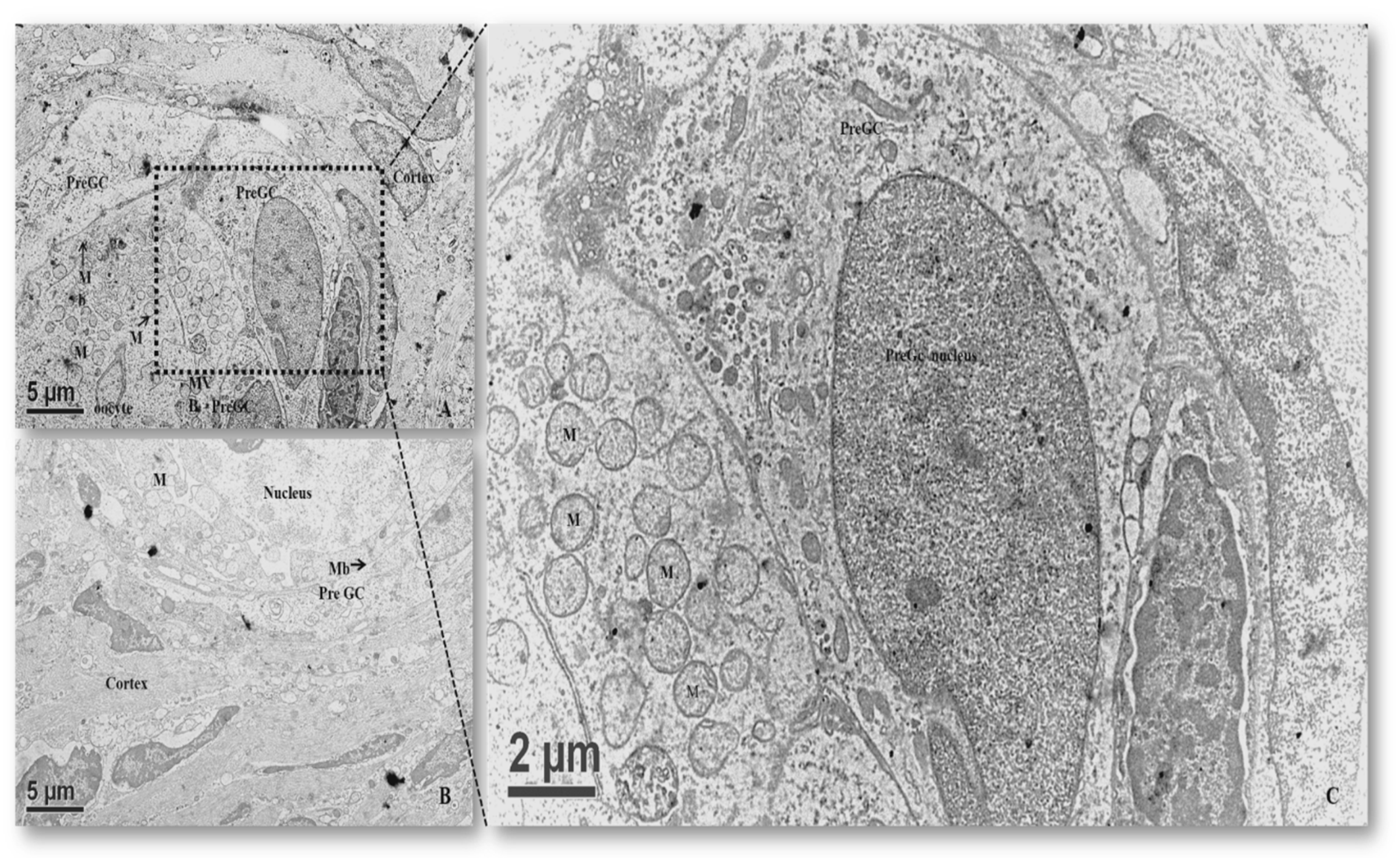
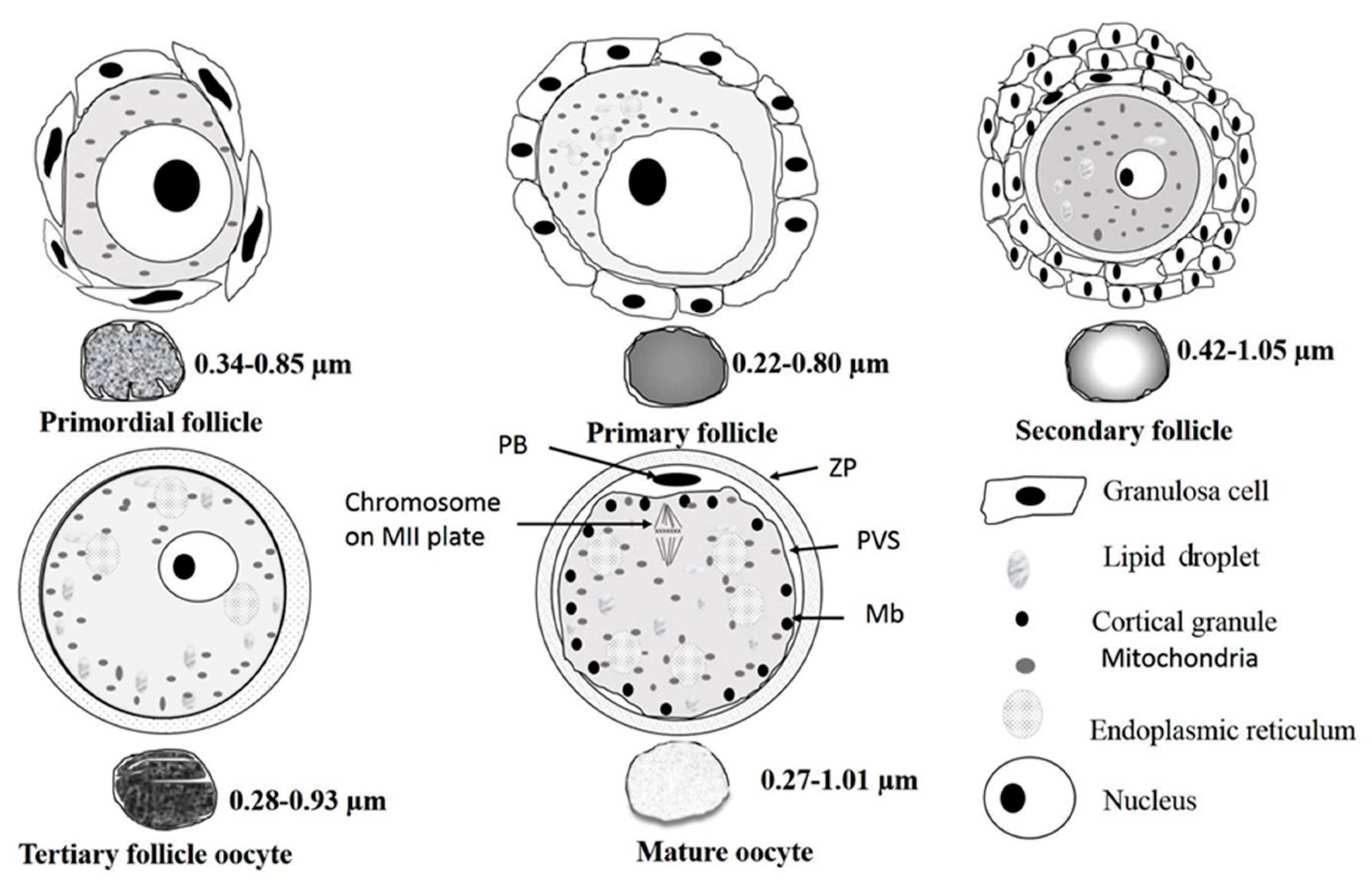
3.2.1. Primordial Follicles
3.2.2. Primary Follicles
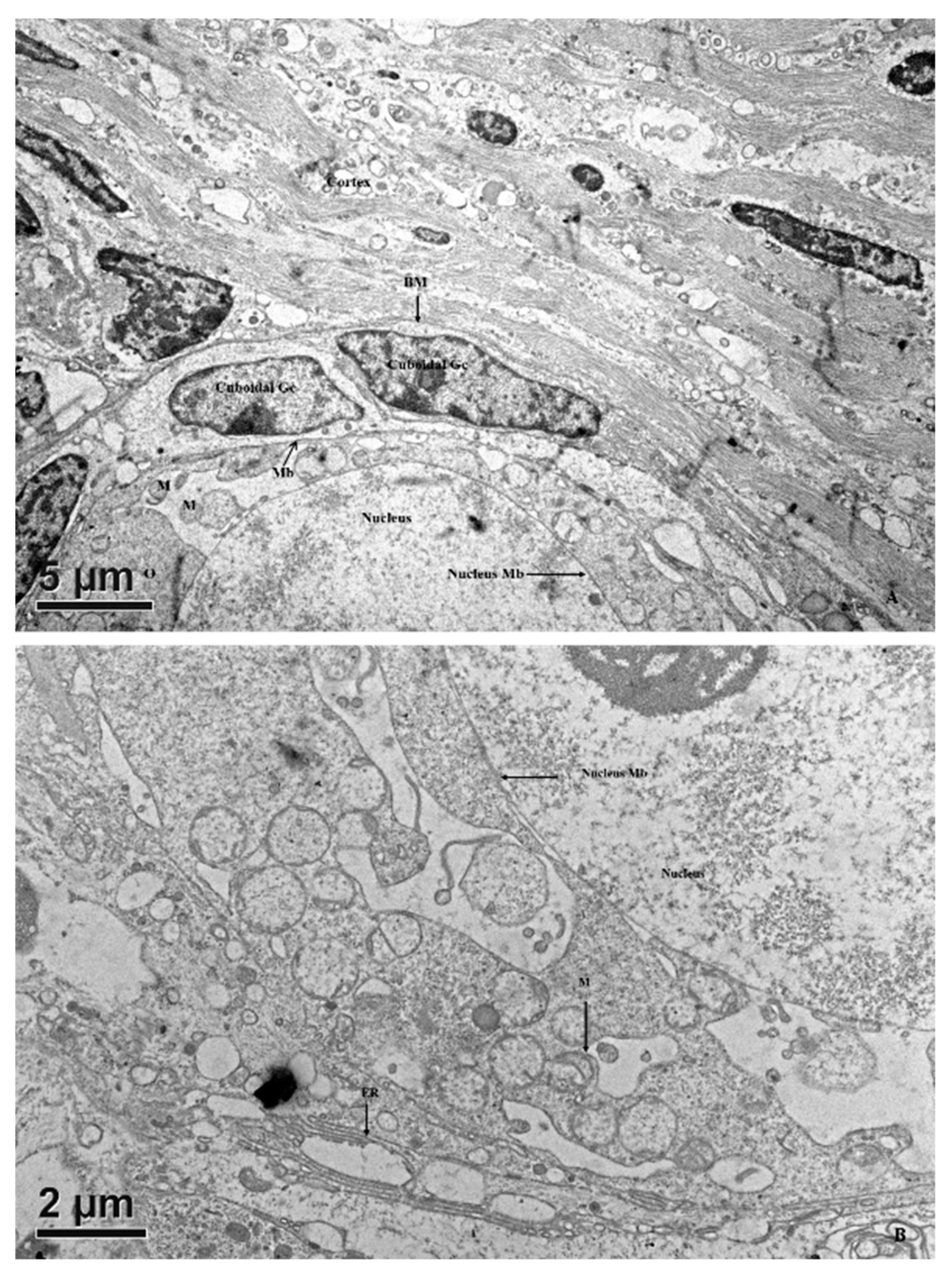
3.2.3. Secondary Follicles
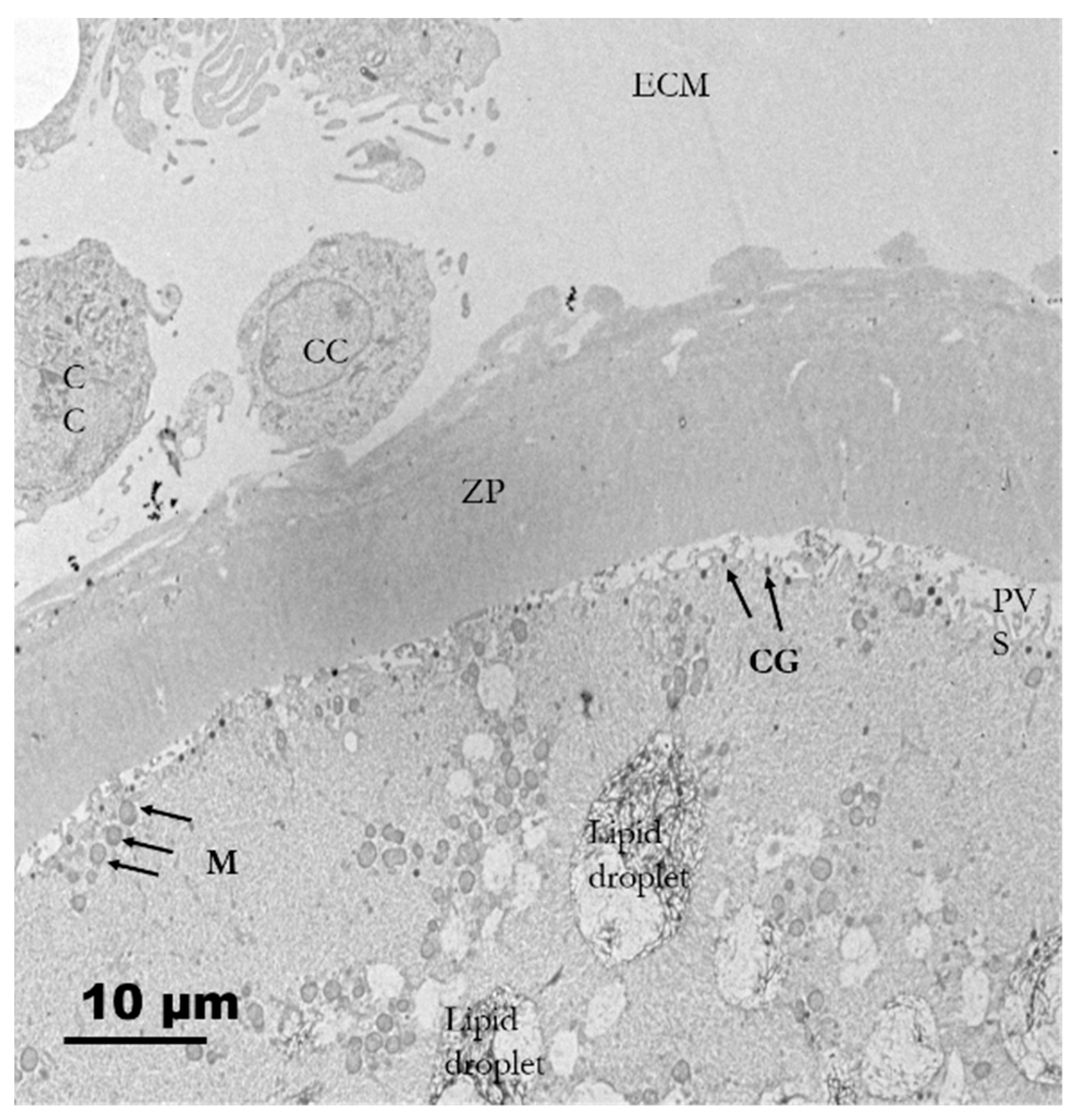
3.2.4. Tertiary Follicles
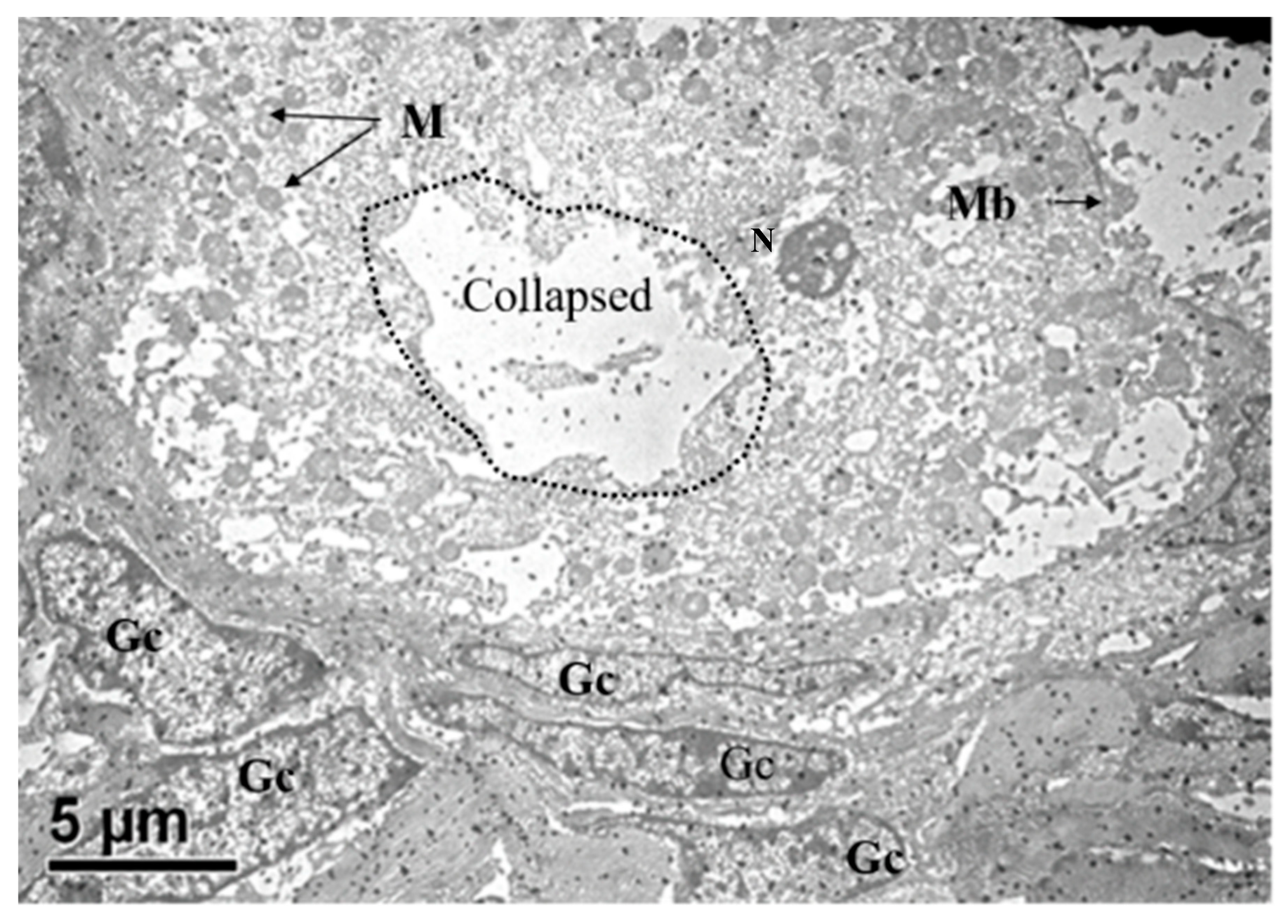
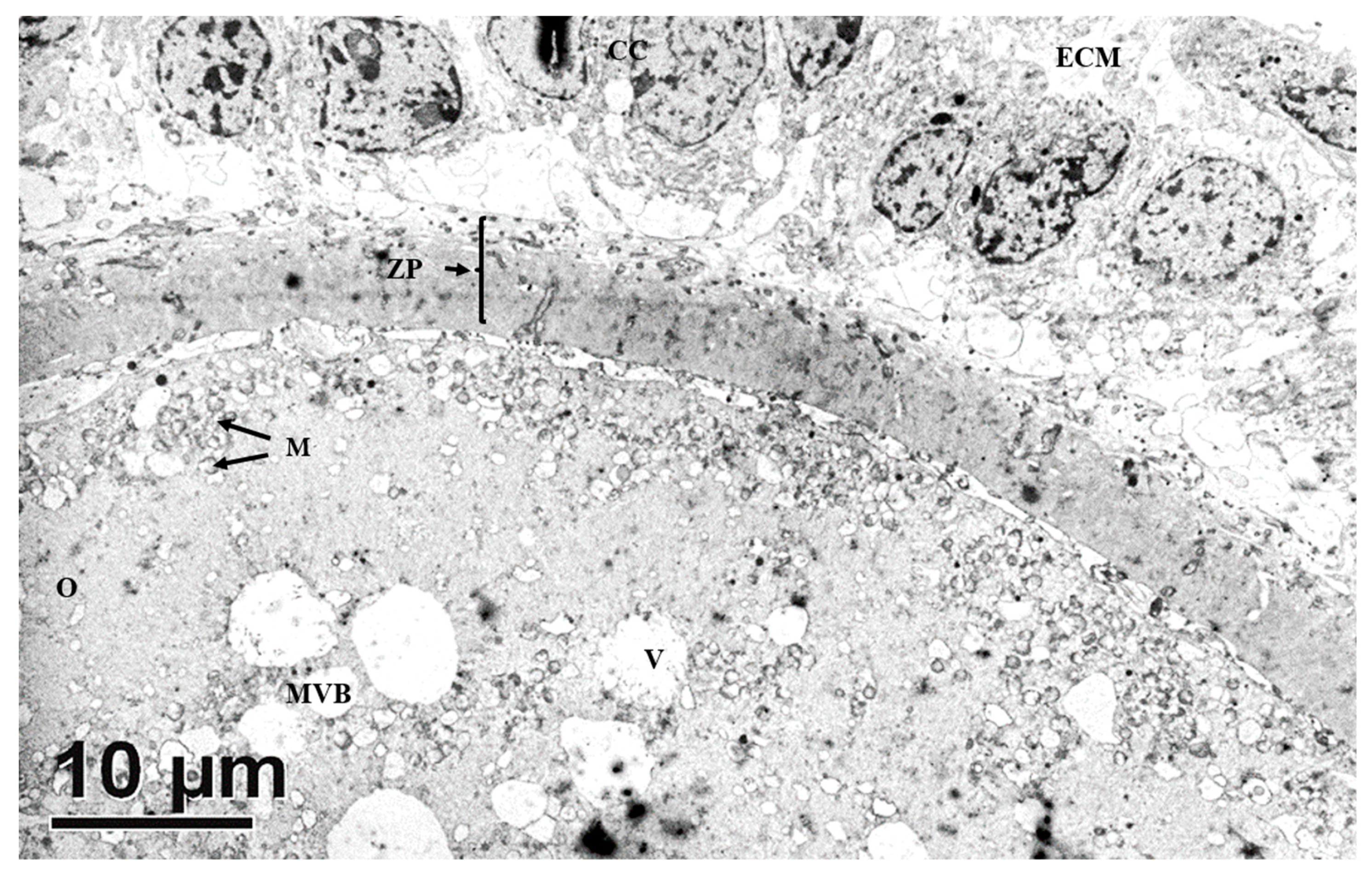
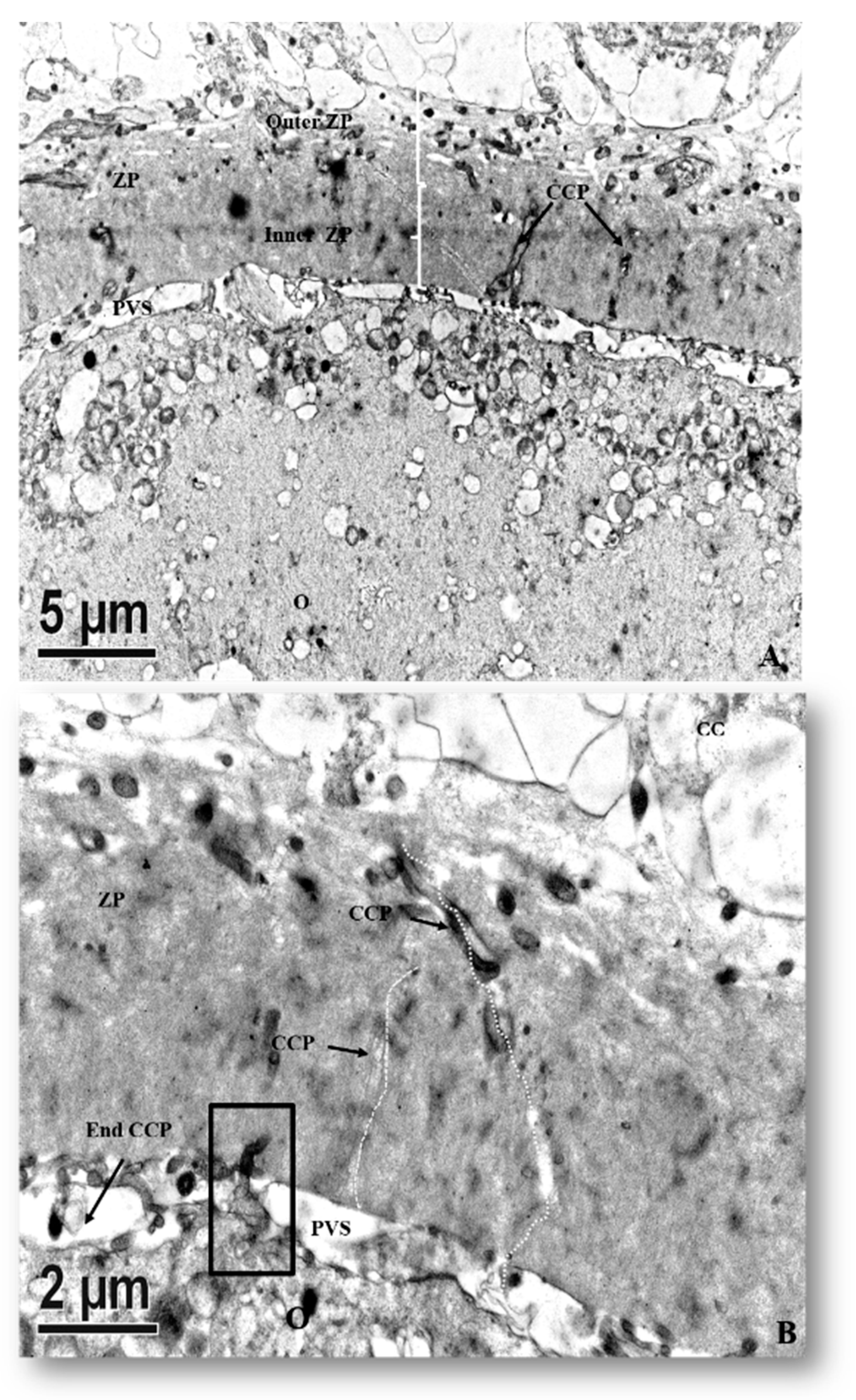
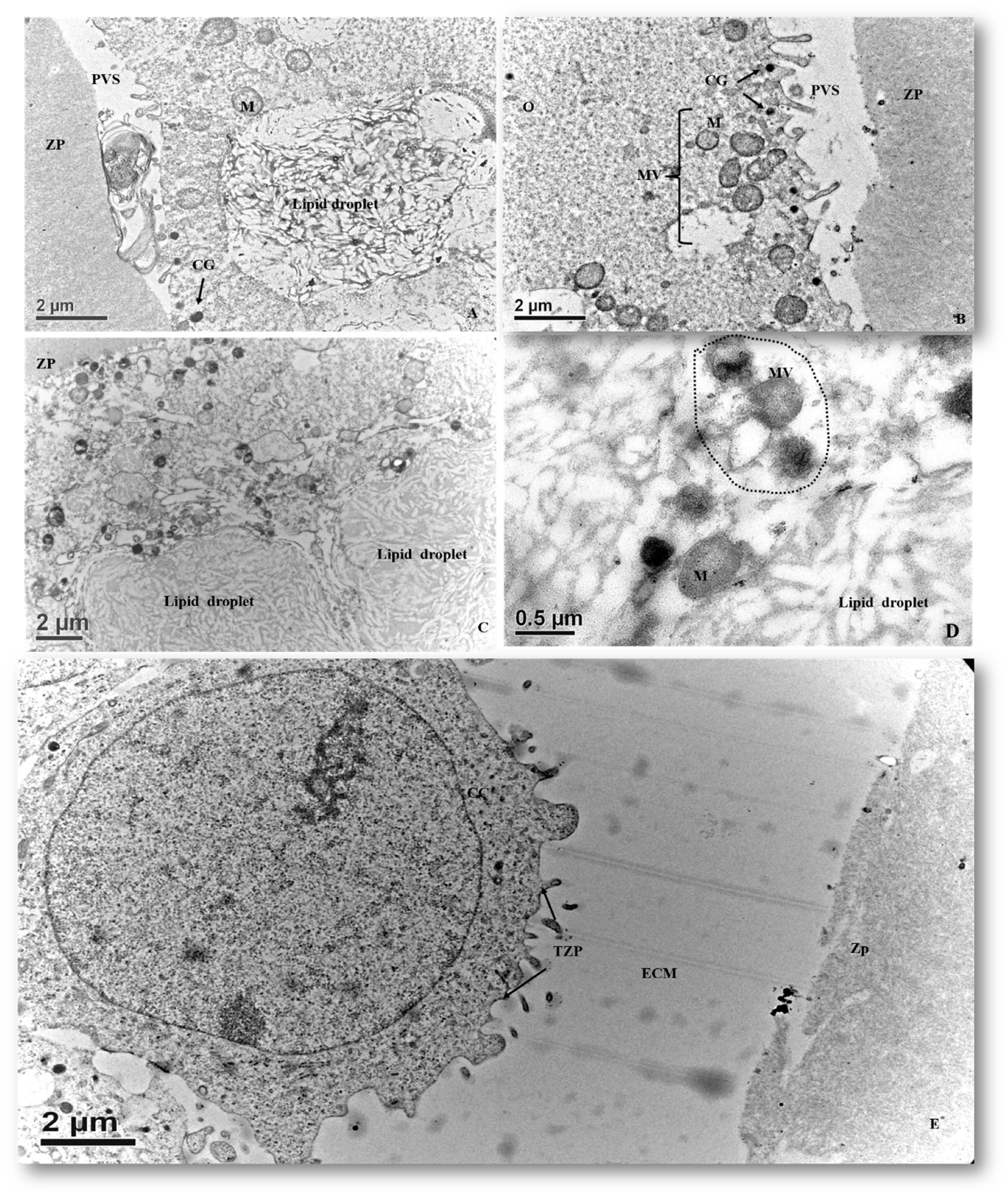
3.2.5. In Vitro Matured Oocytes
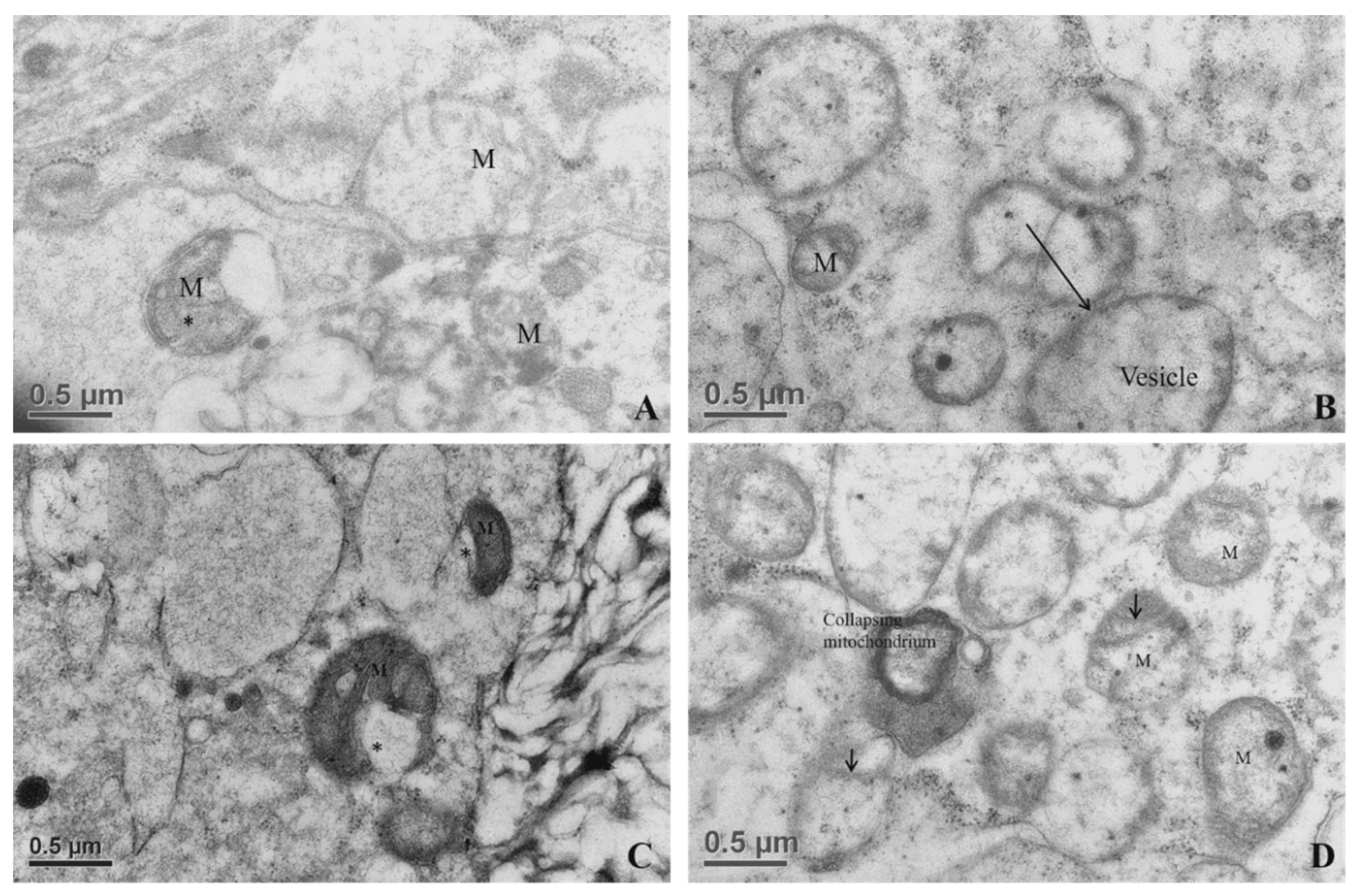
3.2.6. Mitochondrial Morphometry
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Motta, P.M.; Nottola, S.A.; Makabe, S.; Heyn, R. Mitochondrial morphology in human fetal and adult female germ cells. Hum. Reprod. 2000, 15, 129–147. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Rui, R.; Dai, J.; Zhang, C.; Ju, S.; Xie, B.; Lu, X.; Zheng, X. Effects of cryopreservation on the developmental competence, ultrastructure and cytoskeletal structure of porcine oocytes. Mol. Reprod. Dev. 2006, 73, 1454–1462. [Google Scholar] [CrossRef] [PubMed]
- Hajkova, P.; Erhardt, S.; Lane, N.; Haaf, T.; El-Maarri, O.; Reik, W.; Walter, J.; Surani, A. Epigenetic reprogramming in mouse primordial germ cells. Mech. Dev. 2002, 117, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Yokota, S. Nuage proteins: Their localization in subcellular structures of spermatogenic cells as revealed by immunoelectron microscopy. Histochem. Cell Boil. 2012, 138, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.; Arkov, A.L. Next generation organelles: Structure and role of germ granules in the germ line. Mol. Reprod. Dev. 2013, 80, 610–623. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Xun, W.; Zhou, H.; Hou, G.; Yue, W.; Zhang, C.; Ren, Y.; Yang, R. Ultrastructure of germ cells, Sertoli cells and mitochondria during spermatogenesis in mature testis of the Chinese Taihang black goats (Capra hircus). Micron 2013, 50, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Black, J.L.; Erickson, B.H. Oogenesis and ovarian development in the prenatal pig. Anat. Rec. Adv. Integr. Anat. Evol. Boil. 1968, 161, 45–55. [Google Scholar] [CrossRef]
- Anderson, E.; Albertini, D.F. Gap junctions between the oocyte and companion follicle cells in the mammalian ovary. J. Cell Boil. 1976, 71, 680–686. [Google Scholar] [CrossRef] [Green Version]
- Sathananthan, A.H. Ultrastructure of human gametes, fertilization and embryos in assisted reproduction: A personal survey. Micron 2013, 44, 1–20. [Google Scholar] [CrossRef]
- Kafi, M.; Mesbah, F.; Nili, H.; Khalili, A. Chronological and ultrastructural changes in camel (Camelus dromedarius) oocytes during in vitro maturation. Theriogenology 2005, 63, 2458–2470. [Google Scholar] [CrossRef]
- Pepling, M.E. From primordial germ cell to primordial follicle: Mammalian female germ cell development. Genesis 2006, 44, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Dumollard, R.; Rossbach, A.; Lai, F.A.; Swann, K. Redistribution of mitochondria leads to bursts of ATP production during spontaneous mouse oocyte maturation. J. Cell. Physiol. 2010, 224, 672–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, J.B.; Badik, J.R.; Ryan, E.L.; Gleason, T.J.; Broadaway, K.A.; Epstein, M.P.; Fridovich-Keil, J.L. Modifiers of ovarian function in girls and women with classic galactosemia. J. Clin. Endocrinol. Metab. 2013, 98, E1257–E1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, N.; Abdelhafez, F.; Ali, M.Y.; Sayed, E.H.; Falcone, T.; Goldfarb, J.; Abu-Alhassan, A.M. Mouse ovarian follicle cryopreservation using vitrification or slow programmed cooling: Assessment of in vitro development, maturation, ultra-structure and meiotic spindle organization. J. Obstet. Gynaecol. Res. 2011, 37, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kéré, M.; Siriboon, C.; Liao, J.; Lo, N.; Chiang, H.; Fan, Y.; Kastelic, J.P.; Ju, J.-C. Vascular endothelial growth factor A improves quality of matured porcine oocytes and developing parthenotes. Domest. Anim. Endocrinol. 2014, 49, 60–69. [Google Scholar] [CrossRef]
- Cran, D.G. Qualitative and quantitative structural changes during pig oocyte maturation. J. Reprod. Fertil. 1985, 74, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Bielańska-Osuchowska, Z. Oogenesis in pig ovaries during the prenatal period: Ultrastructure and morphometry. Reprod. Boil. 2006, 6, 161–193. [Google Scholar]
- Silva, R.C.; Báo, S.N.; Jivago, J.L.P.; Lucci, C.M. Ultrastructural characterization of porcine oocytes and adjacent follicular cells during follicle development: Lipid component evolution. Theriogenology 2011, 76, 1647–1657. [Google Scholar] [CrossRef] [Green Version]
- Kere, M.; Siriboon, C.; Lo, N.-W.; Nguyen, N.T.; Ju, J.-C. Ascorbic Acid Improves the Developmental Competence of Porcine Oocytes After Parthenogenetic Activation and Somatic Cell Nuclear Transplantation. J. Reprod. Dev. 2012, 59, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Fair, T.; Hulshof, S.C.J.; Hyttel, P.; Greve, T.; Boland, M. Oocyte ultrastructure in bovine primordial to early tertiary follicles. Brain Struct. Funct. 1997, 195, 327–336. [Google Scholar] [CrossRef]
- Fair, T. Follicular oocyte growth and acquisition of developmental competence. Anim. Reprod. Sci. 2003, 78, 203–216. [Google Scholar] [CrossRef]
- Hyttel, P.; Xu, K.P.; Smith, S.; Greve, T. Ultrastructure of in-vitro oocyte maturation in cattle. J. Reprod. Fertil. 1986, 78, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Hyttel, P.; Callesen, H.; Greve, T. Ultrastructural features of preovulatory oocyte maturation in superovulated cattle. J. Reprod. Fertil. 1986, 76, 645–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondadori, R.; Santin, T.R.; Fidelis, A.A.G.; Porfírio, E.P.; Báo, S.N.; Mondadori, R. Buffalo (Bubalus bubalis) Pre-antral Follicle Population and Ultrastructural Characterization of Antral Follicle Oocyte. Reprod. Domest. Anim. 2010, 45, 33–37. [Google Scholar] [CrossRef]
- Santos, S.; Ferreira, M.A.P.; Lima, M.; Sampaio, R.; Cordeiro, M.; Silva, T.; Costa, N.; Miranda, M.D.S.; Ohashi, O. Quantification, Morphology and Ultrastructure of Preantral Follicles of Buffalo (Bubalus bubalis) Foetuses. Reprod. Domest. Anim. 2011, 46, 17–22. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, P.; Liu, Y.; Bian, X.; Ullah, S.; Zhang, Q.; Chen, W.; Le, Y.; Chen, B.; Lin, J.; et al. Pre-spermiogenic initiation of flagellar growth and correlative ultrastructural observations on nuage, nuclear and mitochondrial developmental morphology in the zebrafish Danio rerio. Micron 2014, 66, 1–8. [Google Scholar] [CrossRef]
- Liu, P.-C. Establishment of a soft shell turtle, Pelodiscus sinensis, embryo primary cell culture for studies of soft shell turtle poxvirus-like virus replication and characteristics. Afr. J. Microbiol. Res. 2012, 6, 960–967. [Google Scholar]
- Makabe, S.; Van Blerkom, J. Atlas of Human Female Reproductive Function: Ovarian Development to Early Embryogenesis after In-Vitro Fertilization, 1st ed.; Informa Healthcare: London, UK, 2006. [Google Scholar]
- Andrade, E.R.; Maddox-Hyttel, P.; Landim-Alvarenga, F.D.C.; Silva, J.R.V.; Alfieri, A.F.; Seneda, M.; Figueiredo, J.; Toniolli, R. Ultrastructure of Sheep Primordial Follicles Cultured in the Presence of Indol Acetic Acid, EGF, and FSH. Veter-Med. Int. 2010, 2011, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kacinskis, M.Á.; Lucci, C.M.; Luque, M.C.A.; Báo, S.N. Morphometric and ultrastructural characterization of Bos indicus preantral follicles. Anim. Reprod. Sci. 2005, 87, 45–57. [Google Scholar] [CrossRef] [Green Version]
- Paulini, F.; Silva, R.; Rôlo, J.L.J.D.P.; Lucci, C.M. Ultrastructural changes in oocytes during folliculogenesis in domestic mammals. J. Ovarian Res. 2014, 7, 102. [Google Scholar] [CrossRef] [Green Version]
- Dvorak, M.; Tesarik, J. Ultrastructure of human ovarian follicles. In Biology of the Ovary; Springer: Berlin, Germany, 1980; pp. 121–137. [Google Scholar]
- Senger, P.L.; Saacke, R.G. Unusual Mitochondria of the Bovine Oocyte. J. Cell Boil. 1970, 46, 405–408. [Google Scholar] [CrossRef] [PubMed]
- Fleming, W.N.; Saacke, R.G. Fine structure of the bovine oocyte from the mature graafian follicle. J. Reprod. Fertil. 1972, 29, 203–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assey, R.; Hyttel, P.; Kanuya, N. Oocyte structure in dominant and subordinate follicles in zebu cattle (Bos indicus). Brain Struct. Funct. 1994, 190, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Rüsse, I. Oogenesis in cattle and sheep. Bibl. Anat. 1983, 24, 77–92. [Google Scholar]
- Sun, Q.Y.; Wu, G.M.; Lai, L.; Park, K.W.; Cabot, R.; Cheong, H.T.; Day, B.N.; Prather, R.S.; Schatten, H. Translocation of active mitochondria during pig oocyte maturation, fertilization and early embryo development in vitro. Reproduction 2001, 122, 155–163. [Google Scholar] [CrossRef]
- Adams, E.C.; Hertig, A.T. Studies on guinea pig oocytes. Electron microscope observations on the development of cytoplasmic organelles in oocyte of primordial and primary follicles. J. Cell. Biol. 1964, 21, 397–427. [Google Scholar] [CrossRef] [Green Version]
- Nicosia, S.V. Rabbit Ovarian Follicles. I. Isolation Technique and Characterization at Different Stages of Development. Boil. Reprod. 1975, 13, 423–447. [Google Scholar] [CrossRef]
- Himelstein-Braw, R.; Byskov, A.G.; Peters, H.; Faber, M. Follicular atresia in the infant human ovary. J. Reprod. Fertil. 1976, 46, 55–59. [Google Scholar] [CrossRef] [Green Version]
- Oakberg, E.F. Folicular growth and atresia in the mouse. In vitro 1979, 15, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Nottola, S.; Cecconi, S.; Bianchi, S.; Motta, C.; Rossi, G.; Continenza, M.A.; Macchiarelli, G. Ultrastructure of isolated mouse ovarian follicles cultured in vitro. Reprod. Boil. Endocrinol. 2011, 9, 3. [Google Scholar] [CrossRef] [Green Version]
- Carrijo, O.A., Jr.; Marinho, A.; Campos, A.; Amorim, C.A.; Báo, S.N.; Lucci, C.M. Morphometry, Estimation and Ultrastructure of Ovarian Preantral Follicle Population in Queens. Cells Tissues Organs 2010, 191, 152–160. [Google Scholar] [CrossRef] [PubMed]
- De Lesegno, C.V.; Reynaud, K.; Péchoux, C.; Thoumire, S.; Chastant-Maillard, S. Ultrastructure of canine oocytes during in vivo maturation. Mol. Reprod. Dev. 2007, 75, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Turathum, B.; Saikhun, J.; Sangsuwan, P.; Kitiyanant, Y. Effects of vitrification on nuclear maturation, ultrastructural changes and gene expression of canine oocytes. Reprod. Boil. Endocrinol. 2010, 8, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, H.; Jeong, B.-S.; Yang, X. Dynamic changes of cumulus-oocyte cell communication during in vitro maturation of porcine oocytes. Boil. Reprod. 2000, 63, 723–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, L.A.; Donoghue, A.M.; Igo, W.; Simmons, L.G.; Wildt, D.E.; Rieffenberger, J. Oocyte recovery and maturation in the American black bear (Ursus americanus): A model for endangered ursids. J. Exp. Zool. 1994, 269, 53–61. [Google Scholar] [CrossRef]
- Isachenko, V.; Selman, H.; Isachenko, E.; Montag, M.; El-Danasouri, I.; Nawroth, F. Modified vitrification and cooling of human pronuclear oocytes: Efficacy and effect on ultrastructure. Reprod. Biomed. Online 2003, 7, 211–216. [Google Scholar] [CrossRef]
- Van Blerkom, J. Microtubule mediation of cytoplasmic and nuclear maturation during the early stages of resumed meiosis in cultured mouse oocytes. Proc. Natl. Acad. Sci. USA 1991, 88, 5031–5035. [Google Scholar] [CrossRef] [Green Version]
- Van Blerkom, J.; Sinclair, J.; Davis, P. Mitochondrial transfer between oocytes: Potential applications of mitochondrial donation and the issue of heteroplasmy. Hum. Reprod. 1998, 13, 2857–2868. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Follicular/Oocytes Stages | Primordial Follicle | Primary Follicle | Secondary Follicle | Tertiary Follicle | Matured Oocyte |
|---|---|---|---|---|---|
| Mean ± SEM | 0.60 ± 0.03 ab | 0.51 ± 0.03 a | 0.73 ± 0.03 b | 0.59 ± 0.03 ab | 0.69 ± 0.03 ab |
| Minimum | 0.340 | 0.223 | 0.416 | 0.276 | 0.272 |
| Maximum | 0.859 | 0.803 | 1.047 | 0.926 | 1.014 |
| No. of mitochondria measured | 28 | 28 | 33 | 29 | 31 |
| No. of oocytes evaluated by TEM | 9 | 11 | 7 | 11 | 12 |
| No. of oocytes used for mitochondria evaluation | 5 | 5 | 4 | 4 | 4 |
| No. oocyte hooded mitochondria | 0 | 0 | 0 | 3 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kere, M.; Liu, P.-C.; Chen, Y.-K.; Chao, P.-C.; Tsai, L.-K.; Yeh, T.-Y.; Siriboon, C.; Intawicha, P.; Lo, N.-W.; Chiang, H.-I.; et al. Ultrastructural Characterization of Porcine Growing and In Vitro Matured Oocytes. Animals 2020, 10, 664. https://doi.org/10.3390/ani10040664
Kere M, Liu P-C, Chen Y-K, Chao P-C, Tsai L-K, Yeh T-Y, Siriboon C, Intawicha P, Lo N-W, Chiang H-I, et al. Ultrastructural Characterization of Porcine Growing and In Vitro Matured Oocytes. Animals. 2020; 10(4):664. https://doi.org/10.3390/ani10040664
Chicago/Turabian StyleKere, Michel, Pan-Chen Liu, Yuh-Kun Chen, Pei-Chi Chao, Li-Kuang Tsai, Ting-Yu Yeh, Chawalit Siriboon, Payungsuk Intawicha, Neng-Wen Lo, Hsing-I Chiang, and et al. 2020. "Ultrastructural Characterization of Porcine Growing and In Vitro Matured Oocytes" Animals 10, no. 4: 664. https://doi.org/10.3390/ani10040664
APA StyleKere, M., Liu, P. -C., Chen, Y. -K., Chao, P. -C., Tsai, L. -K., Yeh, T. -Y., Siriboon, C., Intawicha, P., Lo, N. -W., Chiang, H. -I., Fan, Y. -K., & Ju, J. -C. (2020). Ultrastructural Characterization of Porcine Growing and In Vitro Matured Oocytes. Animals, 10(4), 664. https://doi.org/10.3390/ani10040664