Effect of a Diet Supplemented with the Moringa oleifera Seed Powder on the Performance, Egg Quality, and Gene Expression in Japanese Laying Quail under Heat-Stress



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Housing, and Experimental Design
2.2. Productive and Reproductive Measurements
2.3. Sample Collection and Egg Quality Measurements
2.4. Total RNA Isolation and cDNA Synthesis
2.5. Quantitative Real-Time Polymerase Chain Reaction (PCR)
2.6. Statistical Analysis
3. Results
3.1. Productive and Laying Performance
3.2. Egg Quality Measurements
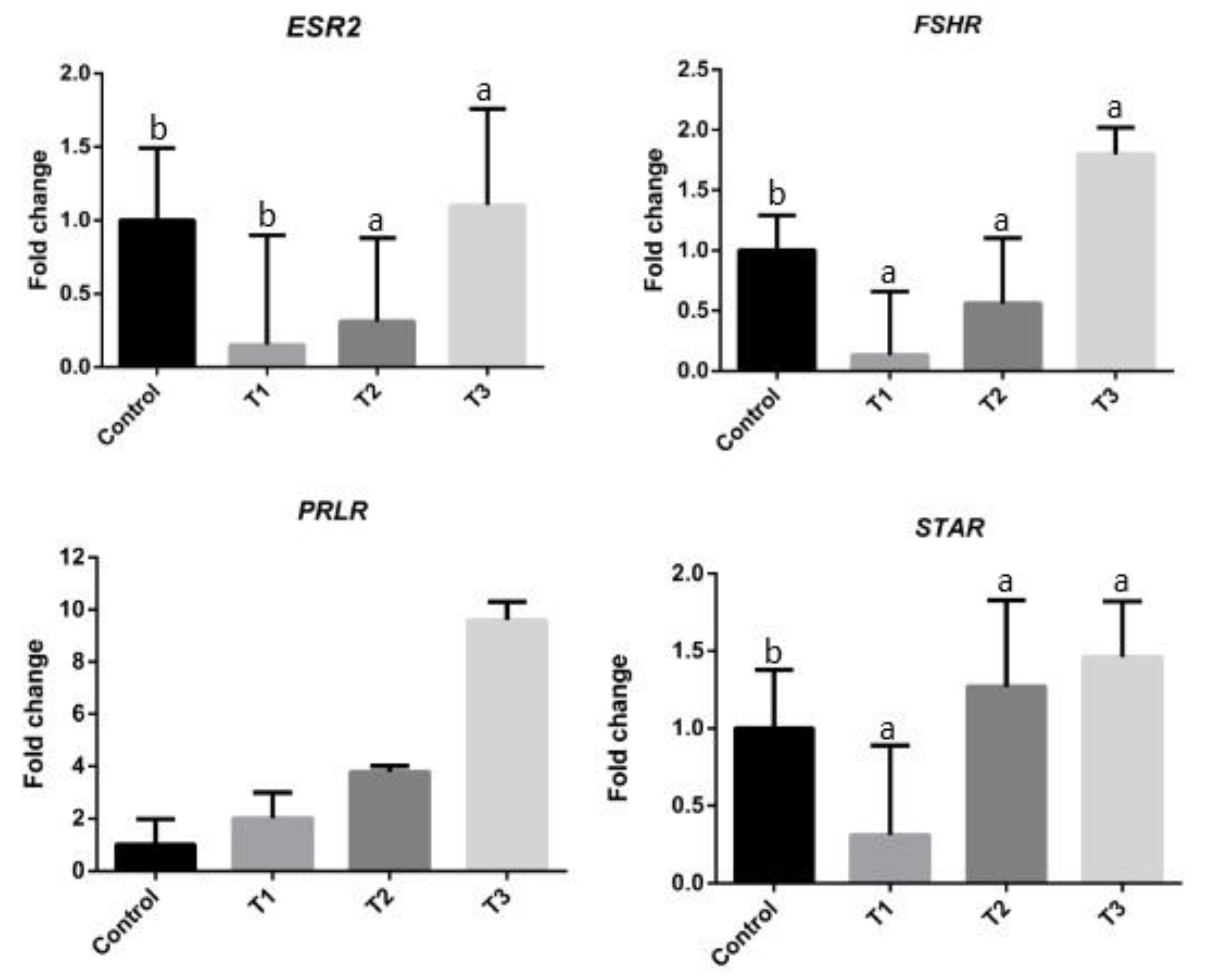
3.3. Expression Profiles of ESR2, FSHR, STAR, and PRLR
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tunsaringkarn, T.; Tungjaroenchai, W.; Siriwong, W. Nutrient benefits of quail (Coturnix coturnix japonica) eggs. Int. J. Sci. Res. Publ. 2013, 3, 1–8. [Google Scholar]
- Tolik, D.; Poawska, E.; Charuta, A.; Nowaczewski, S.; Cooper, R. Characteristics of egg parts, chemical composition and nutritive value of Japanese quail eggs–a review. Folia Biol. 2014, 62, 287–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atakisi, E.; Atakisi, O.; Yaman, H.; Arslan, I. Omega-3 fatty acid application reduces yolk and plasma cholesterol levels in Japanese quails. Food Chem. Toxicol. 2009, 47, 2590–2593. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; Alagawany, M.; Elrys, A.S.; Desoky, E.S.M.; Tolba, H.; Elnahal, A.S.; Swelum, A.A. Effect of forage Moringa oleifera L. (moringa) on animal health and nutrition and its beneficial applications in soil, plants and water purification. Agriculture 2018, 8, 145. [Google Scholar] [CrossRef] [Green Version]
- Saeed, M.; Abbas, G.; Alagawany, M.; Kamboh, A.A.; Abd El-Hack, M.E.; Khafaga, A.F.; Chao, S. Heat stress management in poultry farms: A comprehensive overview. J. Therm. Biol. 2019, 84, 414–425. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; Abdelnour, S.A.; Taha, A.E.; Khafaga, A.F.; Arif, M.; Ayasan, T.; Swelum, A.A.; Abukhalil, H.H.; Alkahtani, S.; Aleya, L.; et al. Herbs as thermoregulatory agents in poultry: An overview. Sci. Total Environ. 2019, 134399. [Google Scholar] [CrossRef]
- Deng, W.; Dong, X.; Tong, J.; Zhang, Q. The probiotic Bacillus licheniformis ameliorates heat stress-induced impairment of egg production, gut morphology, and intestinal mucosal immunity in laying hens. Poult. Sci. 2012, 91, 575–582. [Google Scholar] [CrossRef]
- Rozenboim, I.; Tako, E.; Gal-Garber, O.; Proudman, J.; Uni, Z. The effect of heat stress on ovarian function of laying hens. Poult. Sci. 2007, 86, 1760–1765. [Google Scholar] [CrossRef]
- Abdelnour, S.A.; Abd El-Hack, M.E.; Khafaga, A.F.; Arif, M.; Taha, A.E.; Noreldin, A.E. Stress biomarkers and proteomics alteration to thermal stress in ruminants: A review. J. Therm. Biol. 2019, 79, 120–134. [Google Scholar] [CrossRef]
- Haugen, M.J.; Johnson, A.L. Bone morphogenetic protein 2 inhibits FSH responsiveness in hen granulosa cells. Reproduction 2010, 140, 551. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Lu, Y.; Liu, X.; Xie, X.; Wang, K.; Yu, D. Identification of chicken FSHR gene promoter and the correlations between polymorphisms and egg production in Chinese native hens. Reprod. Domest. Anim. 2019, 54, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Sun, H.; Qu, L.; Ma, M.; Dou, T.; Lu, J.; Guo, J.; Hu, Y.; Wang, X.; Li, Y. Genetic architecture and candidate genes identified for follicle number in chicken. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephens, C.; Johnson, P. Bone morphogenetic protein 15 may promote follicle selection in the hen. Gen. Comp. Endocrinol. 2016, 235, 170–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Xiao, H.; Liang, Z.; Pan, A.; Shen, J.; Pi, J.; Pu, Y.; Du, J.; Chen, Z. Differential expression profiling of estrogen receptor in the ovaries of two egg duck (Anas platyrhynchos) breeds. Czech J. Anim. Sci 2014, 59, 238–243. [Google Scholar] [CrossRef] [Green Version]
- Asli, M.M.; Hosseini, S.A.; Lotfollahian, H.; Shariatmadari, F. Effect of probiotics, yeast, vitamin E and vitamin C supplements on performance and immune response of laying hen during high environmental temperature. Int. J. Poult. Sci. 2007, 6, 895–900. [Google Scholar] [CrossRef]
- Balnave, D.; Brake, J. Nutrition and management of heat-stressed pullets and laying hens. World’s Poult. Sci. J. 2005, 61, 399–406. [Google Scholar] [CrossRef]
- Al-Sagheer, A.A.; Abd El-Hack, M.E.; Alagawany, M.; Naiel, M.A.; Mahgoub, S.A.; Badr, M.M.; Hussein, E.O.; Alowaimer, A.N.; Swelum, A.A. Paulownia leaves as a new feed resource: Chemical composition and effects on growth, carcasses, digestibility, blood biochemistry, and intestinal bacterial populations of growing rabbits. Animals 2019, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Dehshahri, S.; Wink, M.; Afsharypuor, S.; Asghari, G.; Mohagheghzadeh, A. Antioxidant activity of methanolic leaf extract of Moringa peregrina (Forssk.) Fiori. Res. Pharm. Sci. 2012, 7, 111. [Google Scholar]
- Naiel, M.A.; Ismael, N.E.; Abd El-hameed, S.A.; Amer, M.S. The antioxidative and immunity roles of chitosan nanoparticle and vitamin C-supplemented diets against imidacloprid toxicity on Oreochromis niloticus. Aquaculture 2020, 523, 735219. [Google Scholar] [CrossRef]
- Mbikay, M. Therapeutic potential of Moringa oleifera leaves in chronic hyperglycemia and dyslipidemia: A review. Front. Pharm. 2012, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- Arif, M.; Hayat, Z.; Abd El-Hack, M.E.; Saeed, M.; Imran, H.M.; Alowaimer, A.N.; Saadeldin, I.M.; Taha, A.E.; Swelum, A.A. Impacts of supplementing broiler diets with a powder mixture of black cumin, Moringa and chicory seeds. S. Afr. J. Anim. Sci. 2019, 49, 564–572. [Google Scholar] [CrossRef]
- Moyo, B.; Oyedemi, S.; Masika, P.; Muchenje, V. Polyphenolic content and antioxidant properties of Moringa oleifera leaf extracts and enzymatic activity of liver from goats supplemented with Moringa oleifera leaves/sunflower seed cake. Meat Sci. 2012, 91, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.; Shetty, N.; Prakash, M.; Giridhar, P. Effect of dehydration methods on retention of carotenoids, tocopherols, ascorbic acid and antioxidant activity in Moringa oleifera leaves and preparation of a RTE product. J. Food Sci. Technol. 2014, 51, 2176–2182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, N.F.; Sadek, K.M.; Soliman, M.K.; Khalil, R.H.; Khafaga, A.F.; Ajarem, J.S.; Maodaa, S.N.; Allam, A.A. Moringa Oleifera Leaf Extract Repairs the Oxidative Misbalance following Sub-Chronic Exposure to Sodium Fluoride in Nile Tilapia Oreochromis niloticus. Animals 2020, 10, 626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreelatha, S.; Padma, P. Antioxidant activity and total phenolic content of Moringa oleifera leaves in two stages of maturity. Plant Foods Human Nutr. 2009, 64, 303. [Google Scholar] [CrossRef]
- Ashour, E.A.; El-Kholy, M.S.; Alagawany, M.; Abd El-Hack, M.E.; Mohamed, L.A.; Taha, A.E.; Sheikh, A.I.E.; Laudadio, V.; Tufarelli, V. Effect of Dietary Supplementation with Moringa oleifera Leaves and/or Seeds Powder on Production, Egg Characteristics, Hatchability and Blood Chemistry of Laying Japanese Quails. Sustainability 2020, 12, 2463. [Google Scholar] [CrossRef] [Green Version]
- Thu Lan, L.T.; Trong Ngu, N.; Thai Hung, L.; Nguyen Han, L.V.; Hong Nhan, N.T. Moringa oleifera and Calliandra calothyrsus leaf powder as feed supplement in the diet of laying Japanese quails. Livest. Res. 2019, 31. [Google Scholar]
- A.O.A.C. Official Methods of Analysis; Patricia, C. Association of Official Analytical Chemists (AOAC), 16th ed.; A.O.A.C. Official Methods of Analysis: Arlington, VA, USA, 1995. [Google Scholar]
- Reddy, P.; Reddy, V.; Reddy, C.; Rao, P. Egg weight, shape index and hatachability in Khaki Campbell duck eggs. Indian J. Poult. Sci. 1979, 14, 26–32. [Google Scholar]
- Romanoff, A.L.; Romanoff, A.J. The Avian Egg; John Wiley and Sons: New York, NY, USA, 1949. [Google Scholar]
- Cotta, T. Chicken Reproduction and Egg Production; UFLA-Faepe: Lavras, Brazil, 1997; pp. 81–92. [Google Scholar]
- Abd El Naby, W.S.H.; Basha, H.A. Influence of Lighting Systems on Some Muscle Development Related Genes and Production Traits in Japanese quail. AJAVA 2018, 13, 295–300. [Google Scholar] [CrossRef] [Green Version]
- Rao, D.D.; Senzer, N.; Wang, Z.; Kumar, P.; Jay, C.M.; Nemunaitis, J. Bifunctional short hairpin RNA (bi-shRNA): Design and pathway to clinical application. In siRNA Des.; Springer: Clifton, NJ, USA, 2013; pp. 259–278. [Google Scholar]
- Kim, H.-Y. Statistical notes for clinical researchers: Assessing normal distribution (2) using skewness and kurtosis. Rest. Dent. Endod. 2013, 38, 52–54. [Google Scholar]
- Duncans, D. Multiple range and multiple F. test. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- EL-Shoukary, R.D.; Darwish, M.H.; Abdel-Rahman, M.A. Behavioral, performance, carcass traits and hormonal changes of heat stressed broilers feeding black and coriander seeds. J. of Adv. Vet. Res. 2014, 4, 97–101. [Google Scholar]
- Hassan, H.; El-Moniary, M.; Hamouda, Y.; El-Daly, E.F.; Youssef, A.W.; El-Azeem, N.A. Effect of different levels of Moringa oleifera leaves meal on productive performance, carcass characteristics and some blood parameters of broiler chicks reared under heat stress conditions. Asian J. Anim. Vet. Adv. 2016, 11, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Khalique, A.; Pasha, T.; Mehmood, S.; Hussain, K.; Shaheen, M.; Naeem, M.; Shafiq, M. Effect of Moringa oleifera (Lam.) pods as feed additive on egg antioxidants, chemical composition and performance of commercial layers. South African J. of Anim. Sci. 2017, 47, 864–874. [Google Scholar] [CrossRef] [Green Version]
- David, L.; Vidanarachchi, J.; Samarasinghe, K.; Cyril, H.; Dematawewa, C. Effects of moringa based feed additives on the growth performance and carcass quality of broiler chicken. Trop. Agric. Res. 2012, 24, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Opel, H.; Proudman, J.A. Effects of repeated handling and blood sampling on plasma prolactin levels in young turkeys. Poult. Sci. 1982, 61, 1390–1398. [Google Scholar]
- Johnson, A.L. Comparison of three serial blood sampling techniques on plasma hormone concentrations in the laying hen. Poult. Sci. 1981, 60, 2322–2327. [Google Scholar] [CrossRef]
- El Halawani, M.; Silsby, J.; Fehrer, S.; Behnke, E. The influence of acute or repeated immobilization on plasma prolactin levels in the turkey (Meleagris gallopavo). Gen. Comp. Endocrinol. 1985, 59, 410–415. [Google Scholar] [CrossRef]
- Arif, M.; Iram, A.; Bhutta, M.A.; Naiel, M.A.; Abd El-Hack, M.E.; Othman, S.I.; Allam, A.A.; Amer, M.S.; Taha, A.E. The Biodegradation Role of Saccharomyces cerevisiae against Harmful Effects of Mycotoxin Contaminated Diets on Broiler Performance, Immunity Status, and Carcass characteristics. Animals 2020, 10, 238. [Google Scholar] [CrossRef] [Green Version]
- El-Shall, N.A.; Awad, A.M.; Abd El-Hack, M.E.; Naiel, M.A.; Othman, S.I.; Allam, A.A.; Sedeik, M.E. The Simultaneous Administration of a Probiotic or Prebiotic with Live Salmonella Vaccine Improves Growth Performance and Reduces Fecal Shedding of the Bacterium in Salmonella-Challenged Broilers. Animals 2020, 10, 70. [Google Scholar] [CrossRef] [Green Version]
- Berger, M.; Habs, M.; Jahn, S.; Schmahl, D. Toxicological assessment of seeds from Moringa oleifera and Moringa stenopetala, two highly efficient primary coagulants for domestic water treatment of tropical raw waters. East African Med. J. 1984, 61, 712. [Google Scholar]
- Sultana, B.; Anwar, F. Flavonols (kaempeferol, quercetin, myricetin) contents of selected fruits, vegetables and medicinal plants. Food Chem. 2008, 108, 879–884. [Google Scholar] [CrossRef] [PubMed]
- Gombart, A.F.; Pierre, A.; Maggini, S. A Review of Micronutrients and the Immune System-Working in Harmony to Reduce the Risk of Infection. Nutrients 2020, 12, 236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naiel, M.A.; Ismael, N.E.; Shehata, S.A. Ameliorative effect of diets supplemented with rosemary (Rosmarinus officinalis) on aflatoxin B1 toxicity in terms of the performance, liver histopathology, immunity and antioxidant activity of Nile Tilapia (Oreochromis niloticus). Aquaculture 2019, 511, 734264. [Google Scholar] [CrossRef]
- Gopalakrishnan, L.; Doriya, K.; Kumar, D.S. Moringa oleifera: A review on nutritive importance and its medicinal application. Food Sci. Human Wellness 2016, 5, 49–56. [Google Scholar] [CrossRef] [Green Version]
- George, J.W.; Dille, E.A.; Heckert, L.L. Current concepts of follicle-stimulating hormone receptor gene regulation. Biol. Reprod. 2011, 84, 7–17. [Google Scholar] [CrossRef] [Green Version]
- Riry, F.; Elkloub, K.; Moustafa, M.; MAM, M.; Hanan, A.; Youssef, S. Effect of partial replacement of soybean meal by moringa oleifera seed meal on japanese quail performance during laying period. Egy. Poult. Sci. J. 2018, 38, 255–267. [Google Scholar]
- Knapczyk, K.; Duda, M.; Durlej, M.; Galas, J.; Koziorowski, M.; Slomczynska, M. Expression of estrogen receptor α (ERα) and estrogen receptor β (ERβ) in the ovarian follicles and corpora lutea of pregnant swine. Domest. Anim. Endocrinol. 2008, 35, 170–179. [Google Scholar] [CrossRef]
- Rangel, P.; Rodríguez, A.; Rojas, S.; Sharp, P.; Gutierrez, C. Testosterone stimulates progesterone production and STAR, P450 cholesterol side-chain cleavage and LH receptor mRNAs expression in hen (Gallus domesticus) granulosa cells. Reprod. 2009, 138, 961–969. [Google Scholar] [CrossRef]
- Aroyo, A.; Yavin, S.; Arav, A.; Roth, Z. Maternal hyperthermia disrupts developmental competence of follicle-enclosed oocytes: In vivo and ex vivo studies in mice. Theriogenology 2007, 67, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Wolfenson, D.; Lew, B.; Thatcher, W.; Graber, Y.; Meidan, R. Seasonal and acute heat stress effects on steroid production by dominant follicles in cows. Anim. Reprod. Sci. 1997, 47, 9–19. [Google Scholar] [CrossRef]
- Abdel-Azeem, A.; Mohamed, F.; El-Shiekh, S.; Hessin, A. Maximizing Productivity of Lohmann Chickens by Feeding Diets Inclusion Different Levels of Moringa oleifera Leaf Powder as a Safe Feed Additive. J. Anim. Poult. Prod. 2017, 8, 319–328. [Google Scholar] [CrossRef]


{kind=link}
| Parameter | Amount |
|---|---|
| Moisture (g) | 8 |
| Crude protein (g) | 36 |
| Crude fat(g) | 37.2 |
| Carbohydrate (g) | 7.9 |
| Fiber (g) | 3.1 |
| Ash (g) | 14.2 |
| Calcium (mg) | 45 |
| Magnesium (mg) | 598 |
| Phosphorus (mg) | 73 |
| Potassium (mg) | 7 |
| Copper (mg) | 4.2 |
| Iron (mg) | 2 |
| Sulfur (mg) | 0.05 |
| Vitamin E (mg) | 766 |
| Vitamin C (mg) | 6.5 |
| Total amino acids(g) | 20.2 |
| PUFA (mg) | 73 |
| Items | Amount |
|---|---|
| Ingredients, % | |
| Yellow corn | 56 |
| Soybean meal (44%) | 29.29 |
| Corn gluten meal | 3.8 |
| Vegetable oil | 3.5 |
| Salt | 0.31 |
| Dicalcium phosphate 1 | 1.1 |
| Premix 2 | 0.25 |
| Limestone | 5.6 |
| DL-Methionine 3 | 0.11 |
| L-lysine | 0.04 |
| Total | 100.00 |
| Energy and Nutrient level | |
| ME (Kcal /kg) | 2979 |
| Crude protein % | 19.9 |
| Ether extract % | 5.9 |
| Crude fiber % | 3.3 |
| Calcium % | 2.461 |
| Available phosphorus% | 0.33 |
| Methionine % | 0.454 |
| Lysine | 1.002 |
| Methionine + cysteine | 0.788 |
| Product Length (bp) | Primer Sequence (5′–3′) | Gene and ID |
|---|---|---|
| ESR2 (XM_015865466.1) | F: TCACCTGCTGGTATTGGCTC R: GGGATGTAGACTGGACTGTGT | 83 |
| FSHR (XM_015856889.1) | F: CTCTCGGTCTACACGCTGAC R: TTGCGGTTAAGTTGCATGGC | 77 |
| STAR (XM_015883089.1) | F: ATCCCAGCGTCAAAGAGGTG R: CCCAATGATGTTCCCAGGCA | 98 |
| PRLR (NM_001323231.1) | F: AAATGCTGGCGTGAGGTACA R: CCACTGGGGATCAGAATCCG | 99 |
| ß-actin (NM-205518.1) | F: ATGGCTCCGGTATGTGCAA R: TGTCTTTCTGGCCCATACCAA | 120 |
| Item 1 | Dietary Treatments 2 | Sig. | ||||
|---|---|---|---|---|---|---|
| Weeks | CNT | T1 | T2 | T3 | ||
| Egg weight (g/egg) | 7th–11th | 11.41 ± 0.25 | 11.66 ± 0.25 | 11.73 ± 0.26 | 11.55 ± 0.25 | N.S. |
| 11th-15th | 11.27 ± 0.27 c | 12.07 ± 0.27 b | 12.91 ± 0.27 a | 12.29 ± 0.28 ab | ** | |
| Egg production (%) | 7th–11th | 50.51 ± 1.81 c | 52.08 ± 1.76 c | 59.37 ± 0.76 b | 62.20 ± 1.44 a | ** |
| 11th–15th | 48.38 ± 1.95 c | 58.97 ± 1.87 b | 57.52 ± 1.11 b | 65.06 ± 1.91 a | *** | |
| Feed intake (g/bird) | 7th–11th | 34.75 ± 0.85 | 33.75 ± 0.63 | 34.50 ± 1.71 | 32.00 ± 1.63 | N.S. |
| 11th–15th | 33.75 ± 0.25 b | 38.50 ± 1.19 a | 29.75 ± 1.25 c | 30.75 ± 0.63 c | * | |
| Feed conversion ratio | 7th–11th | 3.08 ± 0.10 a | 2.65 ± 0.18 b | 2.80 ± 0.20 b | 2.70 ± 0.17 b | * |
| 11th–15th | 3.11 ± 0.09 a | 2.81 ± 0.10 b | 2.49 ± 0.09 c | 2.47 ± 0.99 c | ** | |
| Item | Dietary Treatments 1 | Sig. | ||||
|---|---|---|---|---|---|---|
| Week | CNT | T1 | T2 | T3 | ||
| Shell Thickness (mm) | 4th | 0.36 ± 0.22 | 0.38 ± 0.15 | 0.42 ± 0.20 | 0.33 ± 0.23 | N.S. |
| 8th | 0.30 ± 0.2 b | 0.39 ± 0.1 a | 0.42 ± 0.25 a | 0.39 ± 0.17 a | ** | |
| Shell% | 4th | 15.27 ± 0.40 | 14.26 ± 0.50 | 15.34 ± 0.5 | 14.08 ± 0.46 | N.S. |
| 8th | 14.63 ± 0.55 b | 14.27 ± 0.45 b | 16.91 ± 0.35 a | 14.25 ± 0.55 b | *** | |
| Albumin% | 4th | 46.49 ± 2.02 | 47.91 ± 2.27 | 50.77 ± 1.75 | 49.84 ± 2.02 | N.S. |
| 8th | 54.88 ± 2.50 b | 54.17 ± 2.48 b | 55.05 ± 2.05 b | 57.46 ± 1.99 a | ** | |
| Yolk % | 4th | 30.38 ± 0.91 b | 32.06 ± 0.97 a | 33.30 ± 0.74 a | 32.88 ± 0.85 a | * |
| 8th | 29.30 ± 1.40 b | 30.31 ± 1.61 ab | 31.97 ± 1.40 a | 30.11 ± 0.99 ab | ** | |
| Yolk Index | 4th | 0.32 ± 0.012 | 0.32 ± 0.014 | 0.32 ± 0.015 | 0.32 ± 0.010 | N.S. |
| 8th | 0.38 ± 0.012 b | 0.39 ± 0.009 b | 0.38 ± 0.013 b | 0.45 ± 0.015 a | ** | |
| Egg Shape Index | 4th | 77.20 ± 0.81 | 77.75 ± 0.82 | 77.50 ± 0.78 | 78.52 ± 0.80 | N.S. |
| 8th | 77.40 ± 0.77 | 76.75 ± 0.92 | 78.45 ± 0.90 | 79.05 ± 0.80 | N.S. | |
| Haugh Unit | 4th | 83.08 ± 1.18 | 84.61 ± 1.16 | 83.45 ± 1.07 | 82.50 ± 0.98 | N.S. |
| 8th | 81.18 ± 1.32 | 81.14 ± 1.27 | 81.54 ± 1.19 | 82.31 ± 1.22 | N.S. | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abou-Elkhair, R.; Abdo Basha, H.; Slouma Hamouda Abd El Naby, W.; Ajarem, J.S.; Maodaa, S.N.; Allam, A.A.; Naiel, M.A.E. Effect of a Diet Supplemented with the Moringa oleifera Seed Powder on the Performance, Egg Quality, and Gene Expression in Japanese Laying Quail under Heat-Stress. Animals 2020, 10, 809. https://doi.org/10.3390/ani10050809
Abou-Elkhair R, Abdo Basha H, Slouma Hamouda Abd El Naby W, Ajarem JS, Maodaa SN, Allam AA, Naiel MAE. Effect of a Diet Supplemented with the Moringa oleifera Seed Powder on the Performance, Egg Quality, and Gene Expression in Japanese Laying Quail under Heat-Stress. Animals. 2020; 10(5):809. https://doi.org/10.3390/ani10050809
Chicago/Turabian StyleAbou-Elkhair, Reham, Heba Abdo Basha, Walaa Slouma Hamouda Abd El Naby, Jamaan S. Ajarem, Saleh N. Maodaa, Ahmed A. Allam, and Mohammed A. E. Naiel. 2020. "Effect of a Diet Supplemented with the Moringa oleifera Seed Powder on the Performance, Egg Quality, and Gene Expression in Japanese Laying Quail under Heat-Stress" Animals 10, no. 5: 809. https://doi.org/10.3390/ani10050809
APA StyleAbou-Elkhair, R., Abdo Basha, H., Slouma Hamouda Abd El Naby, W., Ajarem, J. S., Maodaa, S. N., Allam, A. A., & Naiel, M. A. E. (2020). Effect of a Diet Supplemented with the Moringa oleifera Seed Powder on the Performance, Egg Quality, and Gene Expression in Japanese Laying Quail under Heat-Stress. Animals, 10(5), 809. https://doi.org/10.3390/ani10050809