Size Matters: Zoo Data Analysis Shows that the White Blood Cell Ratio Differs between Large and Small Felids

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
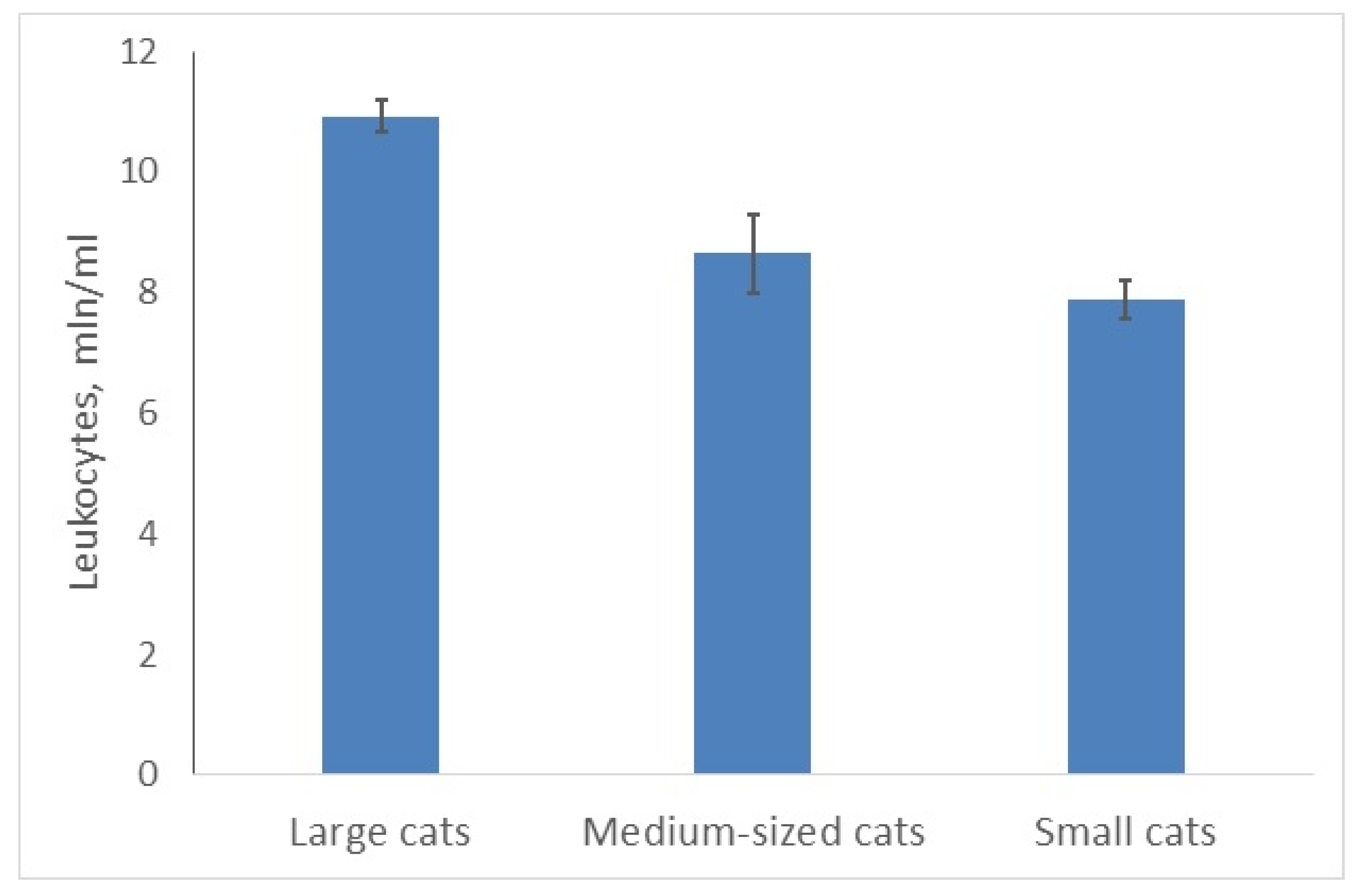
3.1. Larger Cats Have More WBCs Than Smaller Ones
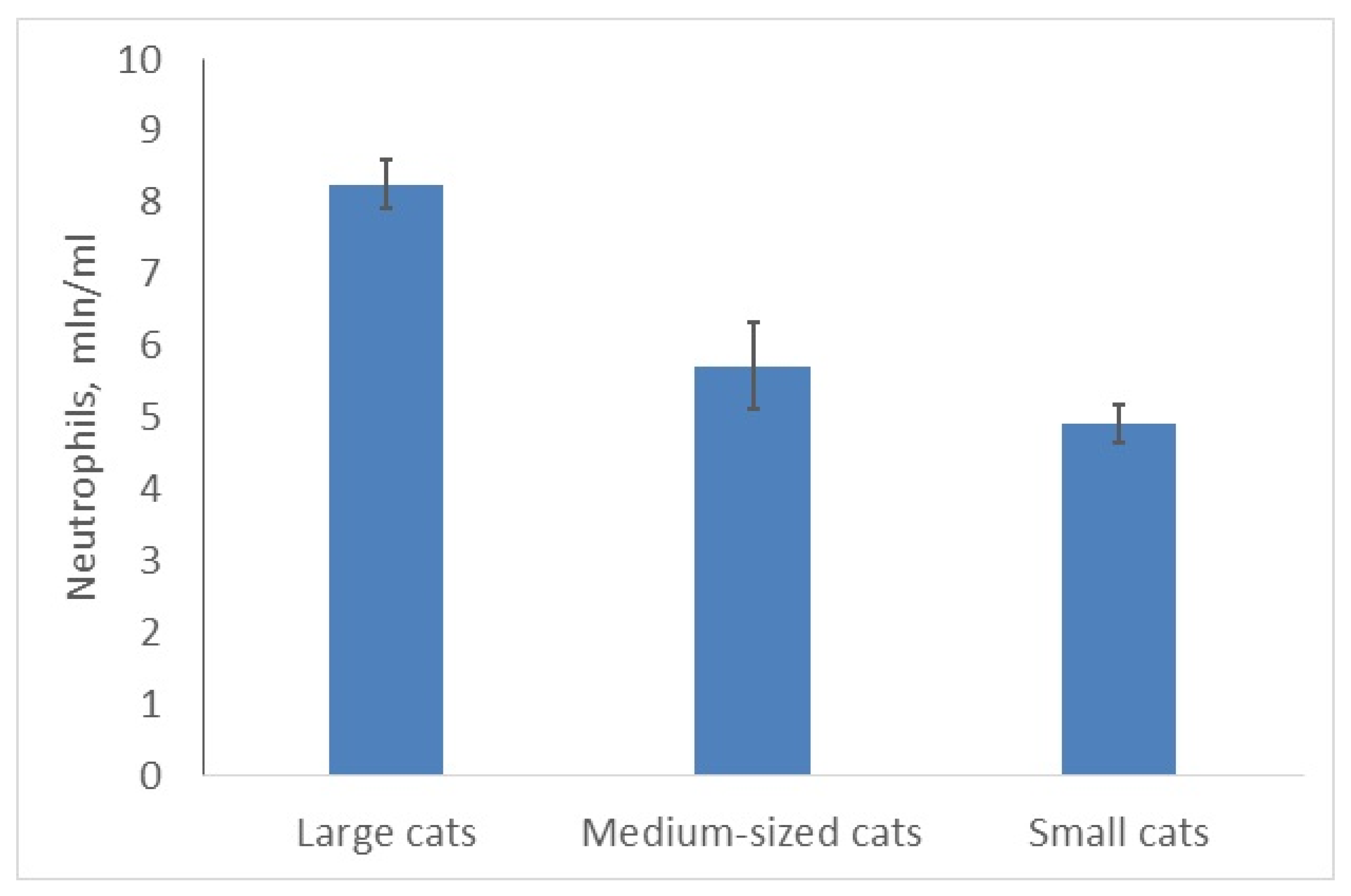
3.2. Larger Cats Have More Neutrophils and Monocytes Than Smaller Felids
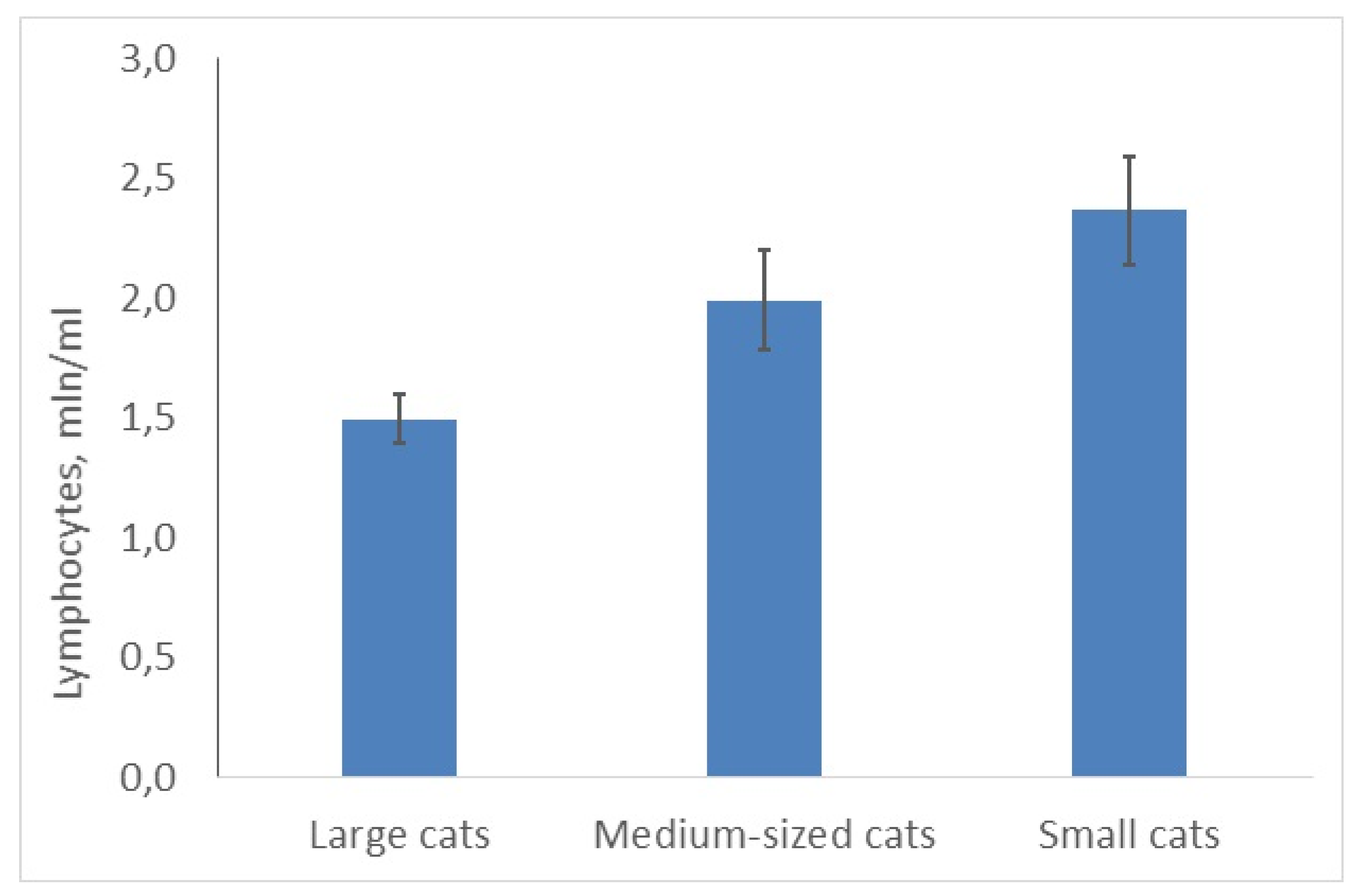
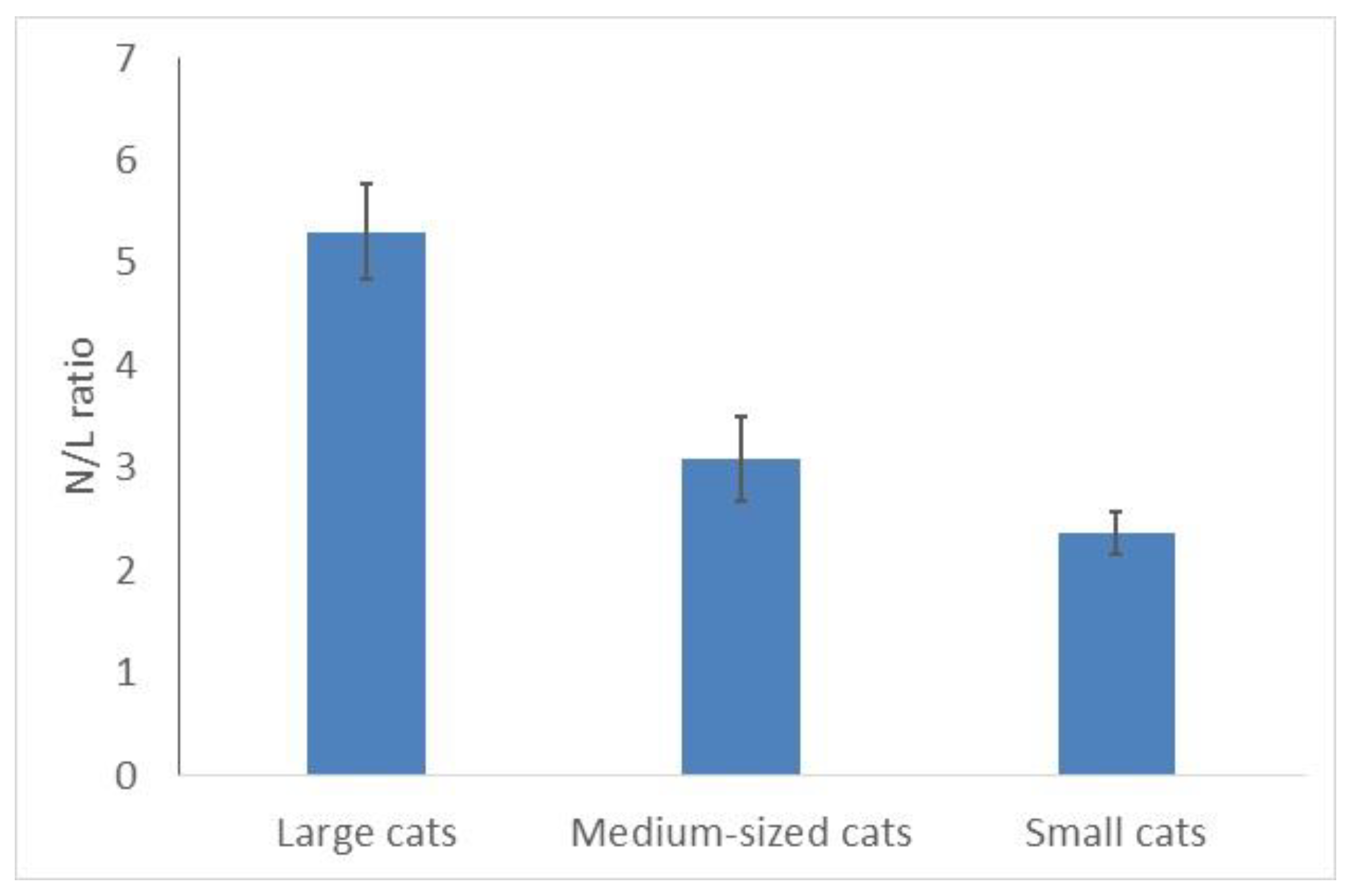
3.3. Small Felids Have More Lymphocytes and a Lower N/L Ratio Than Big Cats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Long, C.A. Evolution of function and form in camelid erythrocytes. In Proceedings of the 2007 WSEAS International Conference on Cellular and Molecular Biology—Biophysics and Bioengineering, Athens, Greece, 26–28 August 2007; pp. 18–24. [Google Scholar]
- Bordes, F.; Blumstein, D.T.; Morand, S. Rodent sociality and parasite diversity. Biol. Lett. 2007, 3, 692–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordes, F.; Morand, S. Coevolution between multiple helminth infestations and basal immune investment in mammals: Cumulative effects of polyparasitism? Parasitol. Res. 2009, 106, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Nunn, C.L.; Gittleman, J.L.; Antonovics, J. Promiscuity and the primate immune system. Science 2000, 290, 1168–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunn, C.L.; Altizer, S.M. Sexual selection, behaviour and sexually transmitted diseases. In Sexual Selection in Primates: New and Comparative Perspectives; Kappeler, P.M., Schaik, C.P., Eds.; Cambridge University Press: Cambridge, UK, 2004; pp. 117–130. [Google Scholar]
- Nunn, C.L.; Gittleman, J.L.; Antonovics, J. A comparative study of white blood cell counts and disease risk in carnivores. Proc. R. Soc. Lond. 2004, 270, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Courtiol, A.; Schneeberger, K.; Greenwood, A.D.; Czirjak, G.A. Circulating white blood cell counts in captive and wild rodents are influenced by body mass rather than testes mass, a correlate of mating promiscuity. Funct. Ecol. 2017, 29, 823–829. [Google Scholar] [CrossRef]
- Schneeberger, K.; Czirjak, G.A.; Voigt, C.C. Measures of the constitutive immune system are linked to diet and roosting habits of neotropical bats. PLoS ONE 2013, 8, e54023. [Google Scholar] [CrossRef]
- Heptner, V.G.; Sludskii, A.A. Mammals of the Soviet Union. Carnivores: Hyenas and Felids, Part 2; Vysshaya Shkola: Moscow, Russia, 1972; Volume 2. (In Russian) [Google Scholar]
- Larivière, S.; Ferguson, S.H. Evolution of induced ovulation in North American carnivores. J. Mammal. 2003, 84, 937–947. [Google Scholar] [CrossRef] [Green Version]
- Naidenko, S.V.; Erofeeva, M.N. Reproduction of the Eurasian lynx, Lynx lynx (Felidae, Carnivora), and the traits of female reproductive strategy. Zool. Zh 2004, 83, 261–269. [Google Scholar]
- Bekoff, M.; Diamond, J.; Mitton, J.B. Life-history patterns and sociality in canids: Body size, reproduction, and behavior. Oecologia 1981, 50, 386–390. [Google Scholar] [CrossRef]
- Iossa, G.; Soulsbury, C.D.; Baker, P.J.; Harris, S. Body mass, territory size, and life-history tactics in a socially monogamous canid, the red fox Vulpes vulpes. J. Mammal. 2008, 89, 1481–1490. [Google Scholar] [CrossRef] [Green Version]
- Kitchener, A.C.; Breitenmoser-Wursten, C.; Eizirik, E.; Gentry, A.; Werdelin, L.; Wilting, A.; Yamaguchi, N.; Abramov, A.V.; Christiansen, P.; Driscoll, C.; et al. A revised taxonomy of the Felidae. The final report of the cat classification task force of the IUCN/SSC cat specialist group. Cat News 2017, 11, 1–80. [Google Scholar]
- Grinell, J.; Packer, C.; Pusey, A.E. Cooperation in male lions: Kinship, reciprocity or mutualism? Anim. Behav. 1995, 49, 95–105. [Google Scholar] [CrossRef]
- Eaton, R.L. Group interaction, spacing and territoriality in cheetahs. Z. für Tierpsychol. 1970, 27, 481–491. [Google Scholar] [CrossRef]
- Chiang, P.J.; Allen, M.L. A review of our current knowledge of clouded leopards (Neofelis nebulosa). Int. J. Avian Wildl. Biol. 2017. [Google Scholar] [CrossRef] [Green Version]
- LaRosa, D.F.; Orange, J.S. Lymphocytes. J. Allergy Clin. Immun. 2008, 121, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Sharma, A. Neutrophils: Cinderella of innate immune system. Inter. Immunopharm. 2010, 10, 1234–1325. [Google Scholar] [CrossRef] [PubMed]
- Dale, D.C.; Boxer, L.; Liles, W.C. The phagocytes: Neutrophils and monocytes. Blood 2008, 112, 935–945. [Google Scholar] [CrossRef]
- Wen, T.; Rothenberg, M.E. The regulatory function of eosinophils. Microbiol. Spectr. 2016. [Google Scholar] [CrossRef] [Green Version]
- Marone, G.; Borriello, F.; Varricchi, G.; Genovese, A.; Granata, F. Basophils: Historical reflections and perspectives. In History of Allergy. Chemical Immunology Allergy; Bergmann, C.R., King, J., Eds.; Karger: Basel, Switzerland, 2014; Volume 100, pp. 172–192. [Google Scholar]
- Smuts, G.L.; Robinson, G.A.; Whyte, I.J. Comparative growth of wild male and female lions (Panthera leo). J. Zool. Lond. 1980, 190, 365–373. [Google Scholar] [CrossRef]
- De Azevedo, F.C.C.; Murray, D.L. Spatial organization and food habits of jaguars (Panthera onca) in a floodplain forest. Biol. Conserv. 2007, 137, 391–402. [Google Scholar] [CrossRef]
- Kitchener, A. The Natural History of the Wild Cats; Christopher Helm Publishers: London, UK, 1991. [Google Scholar]
- Hudson, P.E.; Corr, S.A.; Payne-Davis, R.C.; Clancy, S.N.; Lane, E.; Wilson, A.M. Functional anatomy of the cheetah (Acinonyx jubatus) hindlimb. J. Anat. 2011, 218, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Boast, L.K.; Houser, A.M.; Good, K.; Gusset, M. Regional variation in body size of the cheetah (Acinonyx jubatus). J. Mammal. 2013, 94, 1293–1297. [Google Scholar] [CrossRef]
- Johansson, O.; Malmsten, J.; Mishra, C.; Lkhagvajav, P.; McCarthy, T. Reversible immobilization of free-ranging snow leopards (Panthera uncia) with a combination of medetomidine and tiletamine-zolazepam. J. Wldl. Dis. 2013, 49, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Kitchener, A.C.; Driscoll, C.A.; Yamaguchi, N. What is a snow leopard? Taxonomy, morphology, and phylogeny. In Biodiversity of the World, Conservation from Genes to Landscapes: Snow Leopards; McCarthy, T., Mallon, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 3–11. [Google Scholar]
- De Torre, J.V.; Rivero, M. A morphological comparison of jaguars and pumas in southern Mexico. Therya 2017, 8, 117–122. [Google Scholar] [CrossRef]
- Naidenko, S.V. The Traits of Reproduction and Postnatal Development of Eurasian Lynx; KMK: Moscow, Russia, 2005. (In Russian) [Google Scholar]
- Sunquist, M.; Sunquist, F. Wild Cats of the World; University of Chicago Press: Chicago, IL, USA, 2002. [Google Scholar]
- Sunquist, F. Two species, one design. Int Wildl. 1996, 26, 28–34. [Google Scholar]
- Stuart, C.T. Aspects of the Biology of the Caracal (Felis caracal, Schreber, 1776) in the Cape Province, South Africa. Master’ Thesis, University of Natal, Pietermaritzburg, South Africa, 1992. [Google Scholar]
- Quinn, N.W.S.; Parker, G. Lynx. In Wild Furbearer Management and Conservation in North America; Novak, M., Ed.; Ontario Trappers Association: North Bay, ON, Canada, 1987; pp. 683–694. [Google Scholar]
- Poole, K.G. A review of the Canada Lynx, Lynx canadensis, in Canada. Can. Field Nat. 2003, 117, 360–376. [Google Scholar] [CrossRef] [Green Version]
- Lariviere, S.; Walton, L.R. Lynx rufus. Mammal. Sp. 1997, 563, 1–8. [Google Scholar] [CrossRef]
- Caso, A. Spatial Differences and Local Avoidance of Ocelot (Leopardus pardalis) and Jaguarundi (Puma yagouaroundi) in Northeast Mexico. Ph.D. Thesis, College of Graduate Studies Texas, A&M University, Kingsville, TX, USA, 2013. [Google Scholar]
- Mukherjee, S.; Goyal, S.P.; Johnsingh, A.J.T.; Pitman, M.R.P.L. The importance of rodents in the diet of jungle cat (Felis chaus), caracal (Caracal caracal) and golden jackal (Canis aureus) in Sariska Tiger Reserve, Rajasthan, India. J. Zool. 2004, 262, 405–411. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Krishnan, A.; Tamma, K.; Home, C.R.N.; Joseph, S.; Das, A.; Ramakrishnan, U. Ecology driving genetic variation: A comparative phylogeography of jungle cat (Felis chaus) and leopard cat (Prionailurus bengalensis) in India. PLoS ONE 2010, 5, e13724. [Google Scholar] [CrossRef]
- Miththapala, S. The ecology of the wild cats of Sri Lanka. In Fauna of Sri Lanka: Status of Taxonomy, Research and Conservation; Bambaradeniya, C.N.B., Ed.; IUCN and the Ministry of Environment: Colombo, Sri Lanka, 2004; pp. 224–240. [Google Scholar]
- Drew, S.J.; Perpiñán, D.; Baily, J. Concurrent transitional meningioma and ceruminous gland adenocarcinoma in a Scottish wildcat hybrid (Felis silvestris). J. Comp. Pathol. 2016, 154, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Lucherini, M. Body mass variation in the Geoffroy’s cat (Oncifelis geoffroyi). Rev. Chil. Hist. Nat. 2006, 79, 169–174. [Google Scholar] [CrossRef]
- De Oliveira, T.G. Leopardus wiedi. Mammal. Sp. 1998, 579, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Naidenko, S.V.; Pavlova, E.V.; Kirilyuk, V.E. Detection of seasonal weight loss and a serologic survey of potential pathogens in wild Pallas’ (Felis [Otocolobus] manul) of the Daurian steppe, Russia. J. Wildl. Dis. 2014, 50, 188–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sliwa, A. Home range size and social organization of black-footed cats (Felis nigripes). Mammal. Biol. 2014, 69, 96–107. [Google Scholar] [CrossRef]
- Sliwa, A. Seasonal and sex-specific prey composition of black-footed cats Felis nigripes. Acta Theriol. 2006, 51, 195–204. [Google Scholar]
- Philips, W.A.A. Family: Felidae: Cats. In Manual of the Mammals of Sri Lanka, Part 3, 2nd ed.; Wildlife and Nature Protection Society of Sri Lanka: Colombo, Sri Lanka, 1984; pp. 268–389. [Google Scholar]
- Erofeeva, M.N.; Naidenko, S.V. Spatial organization of felids populations and some traits of their reproductive strategies. Zhurnal Obs. Biol. 2011, 72, 284–297. (In Russian) [Google Scholar]
- Hernandez-Blanco, J.A.; Naidenko, S.V.; Chistopolova, M.D.; Lukarevskiy, V.S.; Kostyrya, A.; Rybin, A.; Sorokin, P.A.; Litvinov, M.N.; Kotlyar, A.K.; Miquelle, D.G.; et al. Social structure and space use of Amur tigers (Panthera tigris altaica) in Southern Russian far east based on GPS telemetry data. Integr. Zool. 2015, 10, 365–375. [Google Scholar] [CrossRef]
- Naidenko, S.V.; Ivanov, E.A.; Mordvintsev, I.N.; Platonov, N.G.; Ershov, R.V.; Rozhnov, V.V. Seropositivity for different pathogens in polar bears (Ursus maritimus) from Barents Sea islands. Biol. Bull. 2013, 40, 779–782. [Google Scholar] [CrossRef]
- Valiathan, R.; Ashman, M.; Asthana, D. Effects of ageing on the immune system: Infants to elderly. Scand. J. Immunol. 2016, 83, 255–266. [Google Scholar] [CrossRef] [Green Version]
- Esan, A.J. Hematological differences in newborn and aging: A review study. Hematol. Transf. Int. J. 2016, 3, 178–190. [Google Scholar]
- Gwak, M.S.; Choi, S.J.; Kim, J.A.; Ko, J.S.; Kim, T.H.; Lee, S.M.; Park, J.A.; Kim, M.H. Effects of gender on white blood cell populations and neutrophil-lymphocyte ratio following gastrectomy in patients with stomach cancer. J. Korean Med. Sci. 2007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doeing, D.C.; Borowicz, J.L.; Crockett, E.T. Gender dimorphism in differential peripheral blood leukocyte counts in mice using cardiac, tail, foot, and saphenous vein puncture methods. BMC Clin. Path. 2003, 3, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemeth, N.; Kiss, F.; Furka, I.; Miko, I. Gender differences of blood rheological parameters in laboratory animals. Clin. Hemorheol. Microcircul. 2010, 45, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Hofmann-Lehmann, R.; Holznagel, E.; Lutz, H. Female cats have lower rates of apoptosis in peripheral blood lymphocytes than male cats: Correlation with estradiol-17b, but not with progesterone blood levels. Vet. Immunol. Immunopathol. 1998, 65, 151–160. [Google Scholar] [CrossRef]
- Naidenko, S.V.; Klyuchnikova, P.S.; Kirilyuk, V.E.; Alekseeva, G.S. Effect of population density on number of leukocytes in domestic cats. Nature Conserv. Res. 2020, 5, 89–96. [Google Scholar] [CrossRef]
- Gatti, L.; Tenconi, P.M.; Guarneri, D.; Bertulessi, C.; Ossola, M.W.; Bosco, P.; Gianotti, G.A. Hemostatic parameters and platelet activation by flow-cytometry in normal pregnancy: A longitudinal study. Intern. J. Clin. Lab. Res. 1994, 24, 217–219. [Google Scholar] [CrossRef]
- Edlestam, G.; Lowbeer, C.; Kral, G.; Gustafsson, S.A.; Venge, P. New reference values for routine blood samples and human neutrophilic lipocalin during third trimester pregnancy. Scand. J. Clin. Lab. Invest. 2001, 61, 583–592. [Google Scholar] [CrossRef]
- Chandra, S.; Tripathi, A.K.; Mishra, S.; Amzarul, M.; Vaish, A.K. Indian physiological changes in hematological parameters during pregnancy. J. Hematol. Blood Transf. 2012, 28, 144–146. [Google Scholar] [CrossRef] [Green Version]
- Roy, S.; Roy, M.; Mishra, S. Hematological and biochemical profile during gestation period in Sahiwal cows. Vet. World 2010, 3, 26–28. [Google Scholar]
- Faas, M.M.; Bouman, A.; Moes, H.; Heineman, M.J.; de Leij, L.; Schuiling, G. The immune response during the luteal phase of the ovarian cycle: A Th2- type response? Fert. Steril. 2000, 74, 1008–1013. [Google Scholar] [CrossRef]
- Dimco, E.; Abeshi, J.; Lika, E.; Dhamo, G. Effect of pregnancy in hematological profile of dogs. Alb. J. Agricul. Sci. 2013, 12, 159–162. [Google Scholar]
- Şimşek, Ö.; Arıkan, Ş.; Çınar, M. Reference values for selected hematological and biochemical blood parameters from prepregnancy to advanced gestation in Angora cats. Turk. J. Vet. Anim. Sci. 2015, 39, 29–33. [Google Scholar] [CrossRef]
- Berman, E. Hemogram of the cat during pregnancy and lactation and after lactation. Am. J. Vet. Res. 1974, 35, 457–460. [Google Scholar]
- Osbaldiston, G.W. Haematological values in healthy cats. Brit. Vet. J. 1978, 134, 524–536. [Google Scholar] [CrossRef]
- Davis, A.K.; Maney, D.L.; Maerz, J.C. The use of leukocyte profiles to measure stress in vertebrates: A review for ecologists. Funct Ecol. 2008, 22, 760–772. [Google Scholar] [CrossRef]
- Pavlova, E.V.; Alekseeva, G.S.; Erofeeva, M.N.; Vasilieva, N.A.; Tchabovsky, A.V.; Naidenko, S.V. The method matters: The effect of handling time on cortisol level and blood parameters in wild cats. J. Exp. Zool. 2018, 329, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Carlstead, K.; Brown, J.L.; Seidensticker, J. Behavioral and adrenocortical responses to environmental changes of leopard cats (Felis bengalensis). Zoo Biol. 1993, 12, 321–331. [Google Scholar] [CrossRef]
- Wielebnowski, N.C.; Fletchall, N.; Carlstead, K.; Busso, J.M.; Brown, J.L. Noninvasive assessment of adrenal activity associated with husbandry and behavioral factors in the North American clouded leopard population. Zoo Biol. 2002, 21, 77–98. [Google Scholar] [CrossRef]
- Naidenko, S.V.; Ivanov, E.A.; Lukarevskii, V.S.; Hernandez-Blanco, J.A.; Sorokin, P.A.; Litvinov, M.N.; Kotlyar, A.K.; Rozhnov, V.V. Activity of the hypothalamo-pituitary-adrenals axis in the Siberian tiger (Panthera tigris altaica) in captivity and in the wild, and its dynamics throughout the year. Biol. Bull. 2011, 338, 301–305. [Google Scholar] [CrossRef]
- Labelle, P.; Mikaelian, I.; Martineau, D.; Beaudin, S.; Blanchette, N.; Lafond, R.; St-Onge, S. Seroprevalence of leptospirosis in lynx and bobcats from Quebec. Can. Vet. J. 2000, 41, 319–320. [Google Scholar]
- Chomel, B.B.; Kikuchi, Y.; Martenson, J.S.; Roelke-Parker, M.E.; Chang, C.C.; Kasten, R.W.; Foley, J.E.; Laudre, J.; Murphy, K.; Swift, P.K.; et al. Seroprevalence of Bartonella infection in American free-ranging and captive pumas (Felis concolor) and bobcats (Lynx rufus). Vet. Res. 2004, 35, 233–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naidenko, S.V.; Hernandez-Blanco, J.A.; Pavlova, E.V.; Erofeeva, M.N.; Sorokin, P.A.; Litvinov, M.N.; Kotlyar, K.; Sulikhan, N.S.; Rozhnov, V.V. Primary study of seroprevalence to virus pathogens in wild felids of South Primorie, Russia. Can. J. Zool. 2018, 96, 839–846. [Google Scholar] [CrossRef] [Green Version]
- Naidenko, S.V.; Hernandez-Blanco, J.A.; Erofeeva, M.N.; Litvinov, M.N.; Rozhnov, V.V. Primary study of seroprevalence to non-viral pathogens in wild felids of South Primorie, Russia. Nat. Conserv. Res. 2019, 4, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Hayward, M.W.; Kerley, G.I.H. Prey preferences of the lion (Panthera leo). J. Zool. 2005, 267, 309–322. [Google Scholar] [CrossRef] [Green Version]
- Hayward, M.W.; Jędrzejewski, W.; Jędrzejewska, B. Prey preferences of the tiger Panthera tigris. J. Zool. 2012, 286, 221–231. [Google Scholar] [CrossRef]
- Clements, H.S.; Tambling, C.J.; Hayward, M.W.; Kerley, G.I.H. An objective approach to determining the weight ranges of prey preferred by and accessible to the five large African carnivores. PLoS ONE 2014, 9, e101054. [Google Scholar] [CrossRef] [Green Version]
- Miller, C.S.; Hebblewhite, M.; Petrunenko, Y.K.; Seryodkin, I.V.; DeCesare, N.J.; Goodrich, J.M.; Miquelle, D.G. Estimating Amur tiger (Panthera tigris altaica) kill rates and potential consumption rates using global positioning system collars. J. Mammal. 2013, 94, 845–855. [Google Scholar] [CrossRef] [Green Version]
- Rozhnov, V.V.; Chistopolova, M.D.; Lukarevskii, V.S.; Hernandez-Blanco, J.A.; Naidenko, S.V.; Sorokin, P.A. Home range structure and space use of a female Amur leopard, Panthera pardus orientalis (Carnivora, Felidae). Biol. Bull. 2015, 42, 821–830. [Google Scholar] [CrossRef]
- DeVault, T.L.; Rhodes, O.E., Jr.; Shivik, J.A. Scavenging by vertebrates: Behavioral, ecological, and evolutionary perspectives on an important energy transfer pathway in terrestrial ecosystems. Oikos 2003, 102, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Werdelin, L.; Yamaguchi, N.; Johnson, W.E.; O’Brien, S.J. Phylogeny and evolution of cats (Felidae). In Biology and Conservation of Wild Felids; Macdonald, D.W., Loveridge, A.J., Eds.; Oxford University Press: Oxford, UK, 2010; pp. 59–82. [Google Scholar]
- Naidenko, S.V.; Hernandez-Blanko, J.A.; Seryodkin, I.V.; Miquelle, D.G.; Blidchenko, E.Y.; Litvinov, M.N.; Rozhnov, V.V. Serum prevalence of the bears in the Russian far east to different pathogens. Biol. Bull. 2019, 46, 960–965. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Number of Tests * | Number of Animals * | Average Body Mass of Adult Individual | References for Body Mass |
|---|---|---|---|---|
| Large cats | ||||
| Tiger (Panthera tigris) | 3485 | 848 | 170.0 | [9] |
| Lion (Panthera leo) | 2943 | 851 | 155.5 | [23] |
| Jaguar (Panthera onca) | 779 | 252 | 80.6 | [24] |
| Leopard (Panthera pardus) | 1008 | 283 | 50.0 | [9,25] |
| Cheetah (Acinonyx jubatus) | 3554 | 751 | 38.0 | [26,27] |
| Snow leopard (Panthera uncia) | 1642 | 438 | 35.0 | [28,29] |
| Medium sized cats | ||||
| Mountain lion (Puma concolor) | 1148 | 316 | 26.3 | [30] |
| Eurasian lynx (Lynx lynx) | 133 | 50 | 20.0 | [31] |
| Clouded leopard (Neofelis nebulosa) | 623 | 194 | 17.0 | [32] |
| Serval (Leptailurus serval) | 653 | 197 | 14.0 | [32] |
| Fishing cat (Prionailurus viverrinus) | 511 | 144 | 12.0 | [33] |
| Caracal (Caracal caracal) | 422 | 111 | 11.5 | [34] |
| Asiatic golden cat (Catopuma temminckii) | 73 | 31 | 10.7 | [33] |
| Canadian lynx (Lynx canadensis) | 313 | 115 | 10.5 | [35,36] |
| Small cats | 70 | |||
| Bobcat (Lynx rufus) | 839 | 193 | 8.2 | [37] |
| Ocelot (Leopardus pardalis) | 758 | 153 | 7.8 | [38] |
| Jungle cat (Felis chaus) | 19 | 6 | 5.0 | [39,40,41] |
| Bengal cat (Prionailurus bengalensis) | 44 | 18 | 5.0 | [40,41] |
| Jaguarundi (Puma yagouaroundi) | 52 | 22 | 4.9 | [38] |
| European wildcat (Felis silvestris) | 238 | 40 | 4.5 | [42] |
| Geoffroy’s cat (Leopardus geoffroyi) | 47 | 17 | 4.3 | [43] |
| Margay (Leopardus wiedi) | 109 | 32 | 3.9 | [44] |
| Pallas’ cat (Otocolobus manul) | 581 | 157 | 3.7 | [45] |
| Black-footed cat (Felis nigripes) | 499 | 106 | 1.6 | [46,47] |
| Sand cat (Felis margarita) | 392 | 105 | 1.5 | [9] |
| Rusty-spotted cat (Prionailurus rubiginosus) | 15 | 7 | 1.3 | [48] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naidenko, S.V.; Alshinetskiy, M.V. Size Matters: Zoo Data Analysis Shows that the White Blood Cell Ratio Differs between Large and Small Felids. Animals 2020, 10, 940. https://doi.org/10.3390/ani10060940
Naidenko SV, Alshinetskiy MV. Size Matters: Zoo Data Analysis Shows that the White Blood Cell Ratio Differs between Large and Small Felids. Animals. 2020; 10(6):940. https://doi.org/10.3390/ani10060940
Chicago/Turabian StyleNaidenko, Sergey V., and Mikhail V. Alshinetskiy. 2020. "Size Matters: Zoo Data Analysis Shows that the White Blood Cell Ratio Differs between Large and Small Felids" Animals 10, no. 6: 940. https://doi.org/10.3390/ani10060940
APA StyleNaidenko, S. V., & Alshinetskiy, M. V. (2020). Size Matters: Zoo Data Analysis Shows that the White Blood Cell Ratio Differs between Large and Small Felids. Animals, 10(6), 940. https://doi.org/10.3390/ani10060940