Characterization of Endolysin LyJH307 with Antimicrobial Activity against Streptococcus bovis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Identification, Cloning, and Overexpression of Recombinant LyJH307
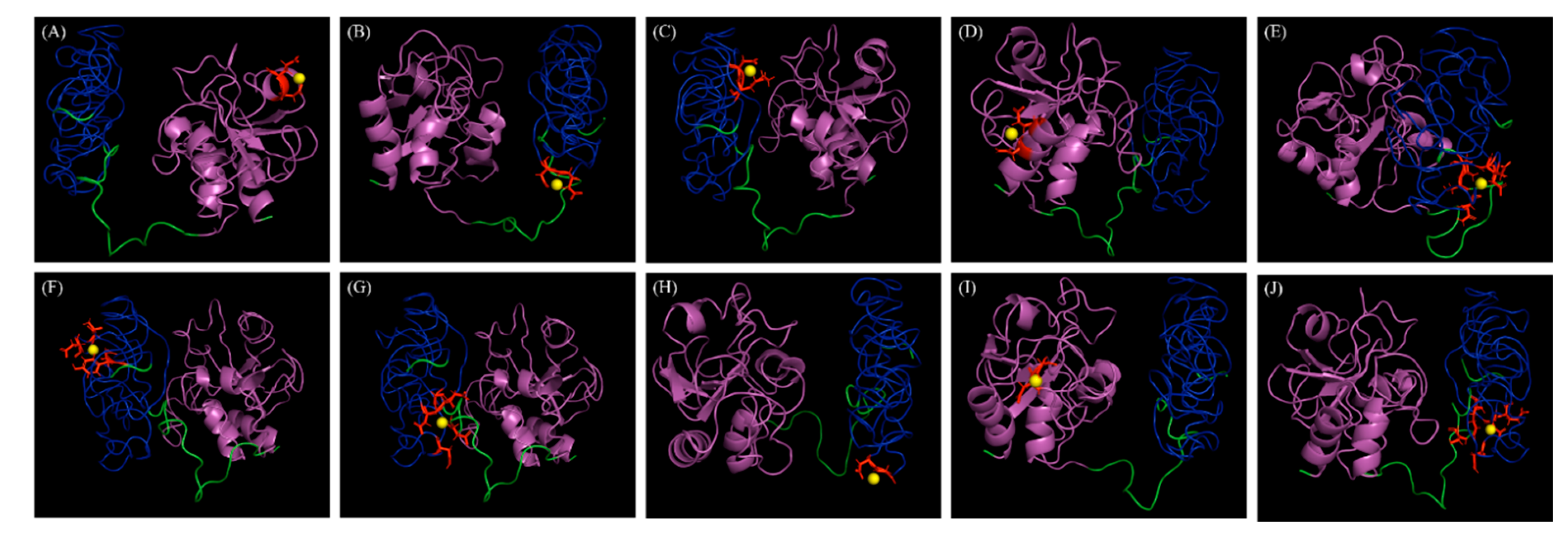
2.3. Structure and Metal Docking Site Prediction of LyJH307
2.4. Characterization of LyJH307
2.5. The Spectrum of Lytic Activity
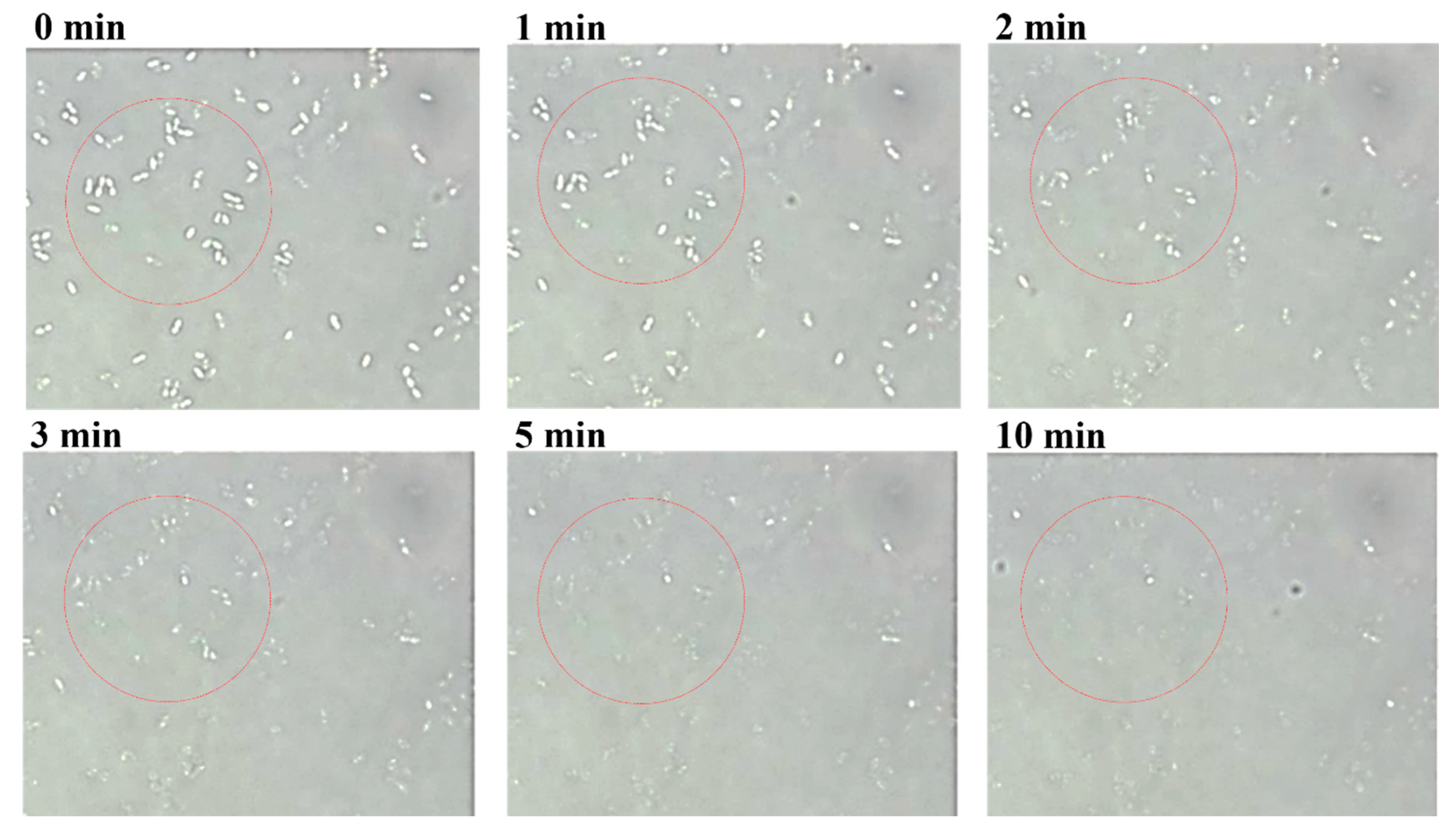
2.6. Optical Microscopy
2.7. Statistical Analysis
3. Results
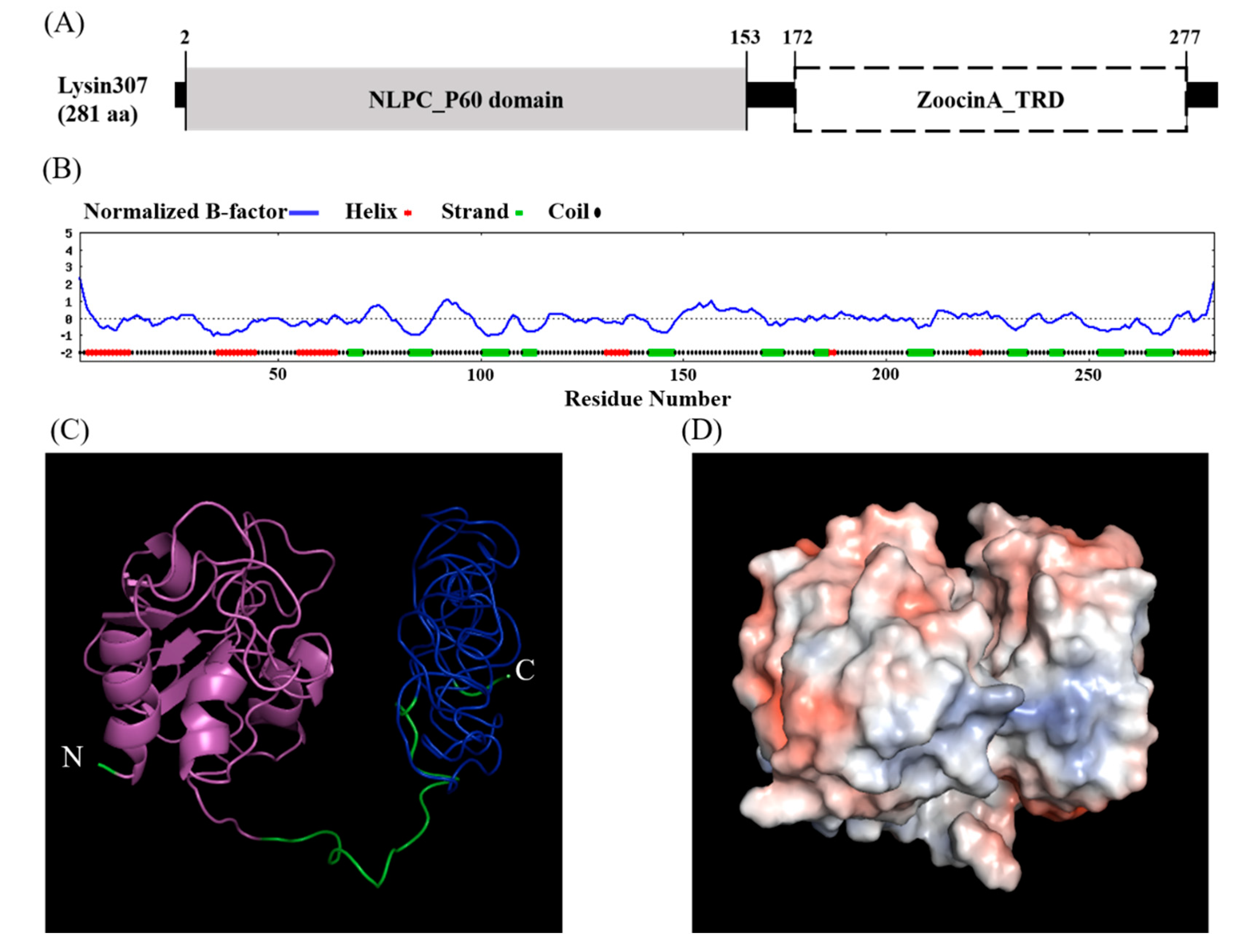
3.1. Sequence Analysis and Overexpression of LyJH307
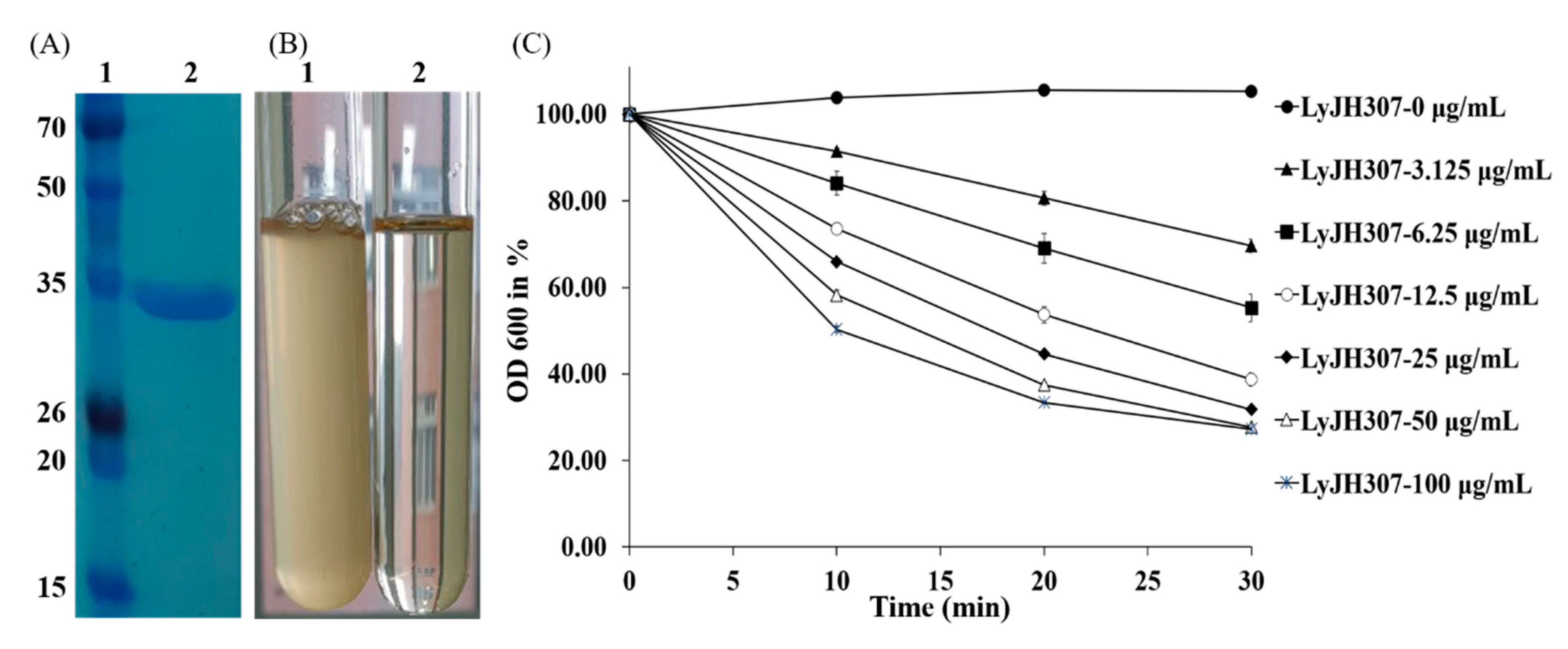
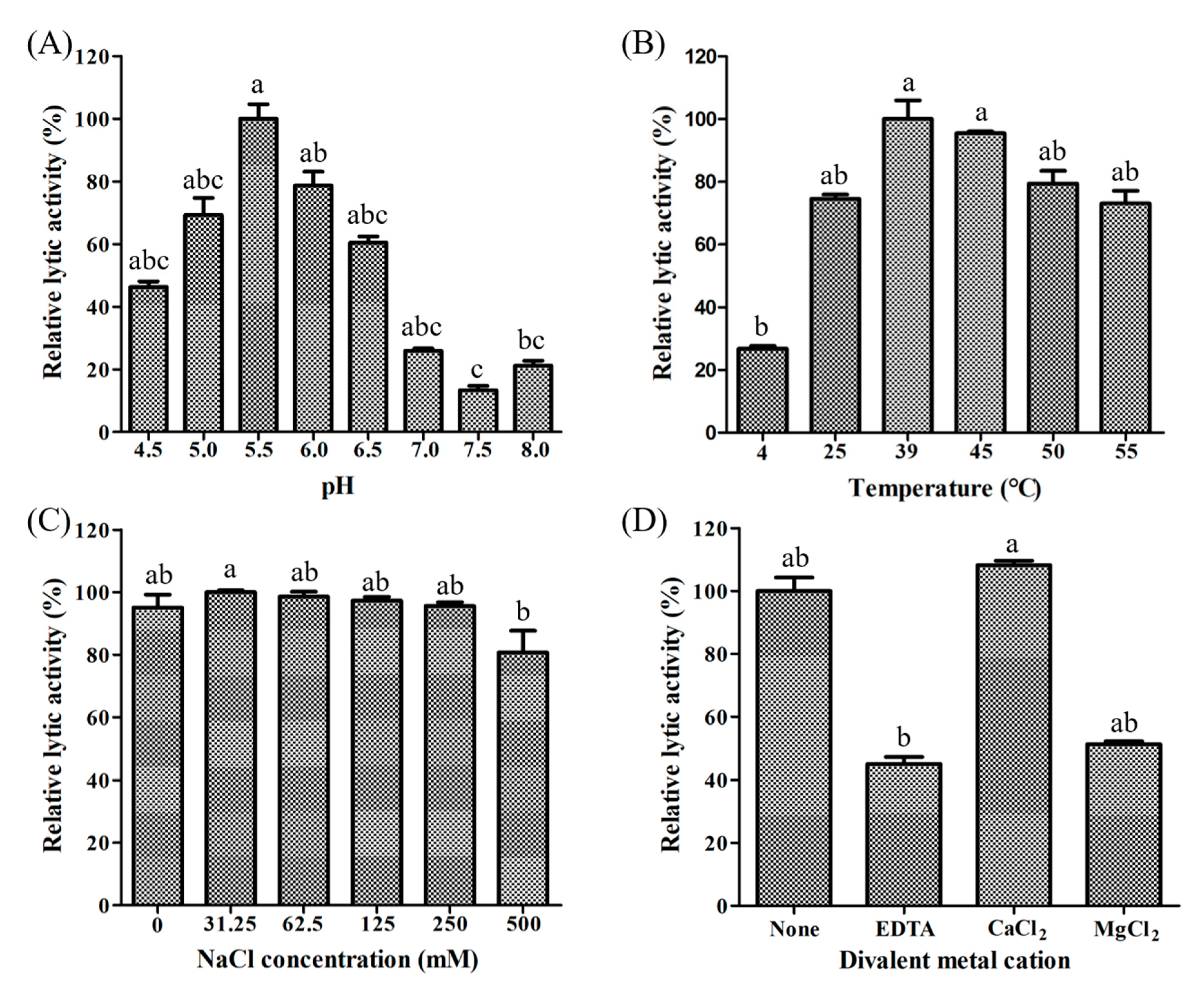
3.2. Characterization of Recombinant LyJH307
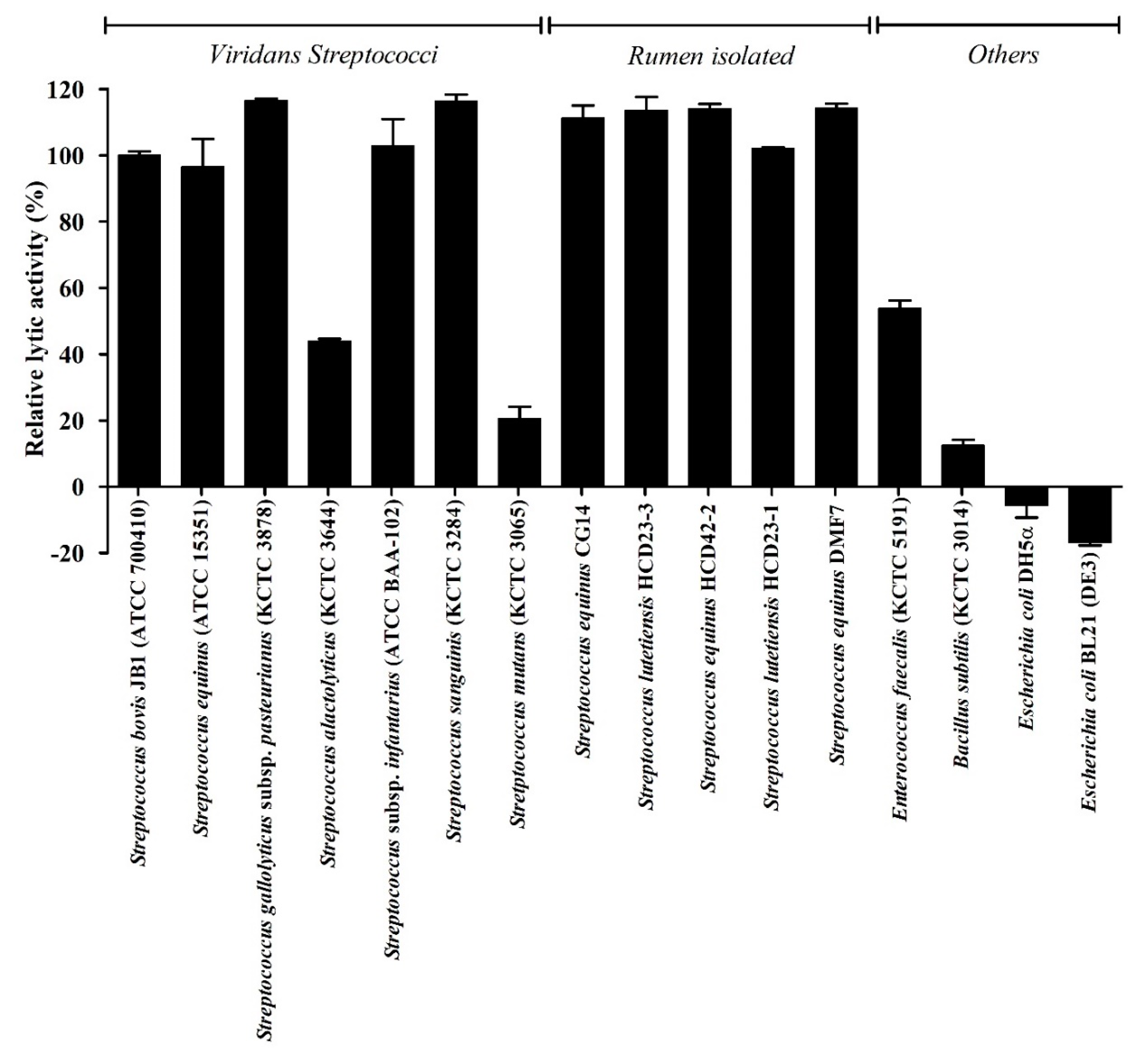
3.3. Antimicrobial Spectrum of LyJH307
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nagaraja, T.; Titgemeyer, E. Ruminal acidosis in beef cattle: The current microbiological and nutritional outlook1, 2. J. Dairy Sci. 2007, 90, E17–E38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enemark, J.M.D.; Jorgensen, R.; Enemark, P.S. Rumen acidosis with special emphasis on diagnostic aspects of subclinical rumen acidosis: A review. Vet. Zootech. 2002, 20, 16–29. [Google Scholar]
- Humer, E.; Petri, R.; Aschenbach, J.; Bradford, B.; Penner, G.; Tafaj, M.; Südekum, K.-H.; Zebeli, Q. Invited review: Practical feeding management recommendations to mitigate the risk of subacute ruminal acidosis in dairy cattle. J. Dairy Sci. 2018, 101, 872–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asanuma, N.; Hino, T. Regulation of fermentation in a ruminal bacterium, Streptococcus bovis, with special reference to rumen acidosis. Anim. Sci. J. 2002, 73, 313–325. [Google Scholar] [CrossRef]
- Russell, J.B.; Hino, T. Regulation of lactate production in Streptococcus bovis: A spiraling effect that contributes to rumen acidosis. J. Dairy Sci. 1985, 68, 1712–1721. [Google Scholar] [CrossRef]
- Hungate, R.E. Evolution of a microbial ecologist. Annu. Rev. Microbiol. 1979, 33, 1–21. [Google Scholar] [CrossRef]
- Snyder, E.; Credille, B. Diagnosis and Treatment of Clinical Rumen Acidosis. Vet. Clin. N. Am. Food Anim. Pract. 2017, 33, 451–461. [Google Scholar] [CrossRef]
- Wang, H.; Pan, X.; Wang, C.; Wang, M.; Yu, L. Effects of different dietary concentrate to forage ratio and thiamine supplementation on the rumen fermentation and ruminal bacterial community in dairy cows. Anim. Prod. Sci. 2015, 55, 189–193. [Google Scholar] [CrossRef] [Green Version]
- Dawson, K.; Rasmussen, M.; Allison, M. Digestive disorders and nutritional toxicity. In The Rumen Microbial Ecosystem; Springer: Dordrecht, The Netherlands, 1997; pp. 633–660. [Google Scholar]
- Owens, F.; Secrist, D.; Hill, W.; Gill, D. Acidosis in cattle: A review. J. Anim. Sci. 1998, 76, 275–286. [Google Scholar] [CrossRef]
- Coe, M.L.; Nagaraja, T.G.; Sun, Y.D.; Wallace, N.; Towne, E.G.; Kemp, K.E.; Hutcheson, J.P. Effect of virginiamycin on ruminal fermentation in cattle during adaptation to a high concentrate diet and during an induced acidosis. J. Anim. Sci. 1999, 77, 2259–2268. [Google Scholar] [CrossRef] [Green Version]
- Nagaraja, T.; Avery, T.; Galitzer, S.; Harmon, D. Effect of ionophore antibiotics on experimentally induced lactic acidosis in cattle. Am. J. Vet. Res. 1985, 46, 2444–2452. [Google Scholar] [PubMed]
- Nagaraja, T.G.; Avery, T.B.; Bartley, E.E.; Roof, S.K.; Dayton, A.D. Effect of Lasalocid, Monensin or Thiopeptin on Lactic Acidosis in Cattle. J. Anim. Sci. 1982, 54, 649–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, J.B.; Houlihan, A.J. Ionophore resistance of ruminal bacteria and its potential impact on human health. FEMS Microbiol. Rev. 2003, 27, 65–74. [Google Scholar] [CrossRef]
- Donovan, D.M. Bacteriophage and Peptidoglycan Degrading Enzymes with Antimicrobial Applications. Recent Pat. Biotechnol. 2007, 1, 113–122. [Google Scholar] [CrossRef]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [Green Version]
- Arends, J.; Zanen, H. Meningitis caused by Streptococcus suis in humans. Rev. Infect. Dis. 1988, 10, 131–137. [Google Scholar] [CrossRef]
- Cole, J.N.; Barnett, T.C.; Nizet, V.; Walker, M.J. Molecular insight into invasive group A streptococcal disease. Nat. Rev. Microbiol. 2011, 9, 724. [Google Scholar] [CrossRef]
- Maestro, B.; Sanz, J.M. Novel approaches to fight Streptococcus pneumoniae. Recent Pat. Antiinfect. Drug Discov. 2007, 2, 188–196. [Google Scholar] [CrossRef]
- Altermann, E.; Schofield, L.R.; Ronimus, R.S.; Beattie, A.K.; Reilly, K. Inhibition of rumen methanogens by a novel archaeal lytic enzyme displayed on tailored bionanoparticles. Front. Microbiol. 2018, 9, 2378. [Google Scholar] [CrossRef] [Green Version]
- Swift, S.M.; Waters, J.J.; Rowley, D.T.; Oakley, B.B.; Donovan, D.M. Characterization of two glycosyl hydrolases, putative prophage endolysins, that target Clostridium perfringens. FEMS Microbiol. Lett. 2018, 365, fny179. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.-F.; Cheng, C.-W.; Shih, C.-S.; Hwang, J.-K.; Yu, C.-S.; Lu, C.-H. MIB: Metal ion-binding site prediction and docking server. J. Chem. Inf. Model. 2016, 56, 2287–2291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, B.; Yun, J.; Lim, J.-A.; Shin, H.; Heu, S.; Ryu, S. Characterization of LysB4, an endolysin from the Bacillus cereus-infecting bacteriophage B4. BMC Microbiol. 2012, 12, 33. [Google Scholar] [CrossRef] [Green Version]
- Linden, S.B.; Zhang, H.; Heselpoth, R.D.; Shen, Y.; Schmelcher, M.; Eichenseher, F.; Nelson, D.C. Biochemical and biophysical characterization of PlyGRCS, a bacteriophage endolysin active against methicillin-resistant Staphylococcus aureus. Appl. Microbiol. Biotechnol. 2015, 99, 741–752. [Google Scholar] [CrossRef]
- Mónico, A.; Martínez-Senra, E.; Zorrilla, S.; Pérez-Sala, D. Drawbacks of dialysis procedures for removal of EDTA. PLoS ONE 2017, 12, e0169843. [Google Scholar] [CrossRef]
- Guruprasad, K.; Reddy, B.B.; Pandit, M.W. Correlation between stability of a protein and its dipeptide composition: A novel approach for predicting in vivo stability of a protein from its primary sequence. Protein Eng. Des. Sel. 1990, 4, 155–161. [Google Scholar] [CrossRef]
- Ji, W.; Huang, Q.; Sun, L.; Wang, H.; Yan, Y.; Sun, J. A novel endolysin disrupts Streptococcus suis with high efficiency. FEMS Microbiol. Lett. 2015, 362, fnv205. [Google Scholar] [CrossRef] [Green Version]
- Schmelcher, M.; Powell, A.M.; Camp, M.J.; Pohl, C.S.; Donovan, D.M. Synergistic streptococcal phage λSA2 and B30 endolysins kill streptococci in cow milk and in a mouse model of mastitis. Appl. Microbiol. Biotechnol. 2015, 99, 8475–8486. [Google Scholar] [CrossRef]
- Celia, L.K.; Nelson, D.; Kerr, D.E. Characterization of a bacteriophage lysin (Ply700) from Streptococcus uberis. Vet. Microbiol 2008, 130, 107–117. [Google Scholar] [CrossRef]
- Anantharaman, V.; Aravind, L. Evolutionary history, structural features and biochemical diversity of the NlpC/P60 superfamily of enzymes. Genome Biol. 2003, 4, R11. [Google Scholar] [CrossRef] [Green Version]
- Ohnishi, R.; Ishikawa, S.; Sekiguchi, J. Peptidoglycan hydrolase LytF plays a role in cell separation with CwlF during vegetative growth of Bacillus subtilis. J. Bacteriol. 1999, 181, 3178–3184. [Google Scholar] [CrossRef] [Green Version]
- Simmonds, R.; Naidoo, J.; Jones, C.; Tagg, J. The streptococcal bacteriocin-like inhibitory substance, zoocin A, reduces the proportion of Streptococcus mutans in an artificial plaque. Microb. Ecol. Health Dis. 1995, 8, 281–292. [Google Scholar] [CrossRef] [Green Version]
- Akesson, M.; Dufour, M.; Sloan, G.L.; Simmonds, R.S. Targeting of streptococci by zoocin A. FEMS Microbiol. Lett. 2007, 270, 155–161. [Google Scholar] [CrossRef]
- Janda, W.M. The genus Streptococcus–part I: Emerging pathogens in the “Pyogenic Cocci” and the “Streptococcus bovis” groups. Clin. Microbiol. Newsl. 2014, 36, 157–166. [Google Scholar] [CrossRef]
- Pompilio, A.; Di Bonaventura, G.; Gherardi, G. An overview on Streptococcus bovis/Streptococcus equinus complex isolates: Identification to the species/subspecies level and antibiotic resistance. Int. J. Mol. Sci. 2019, 20, 480. [Google Scholar] [CrossRef] [Green Version]
- Lai, A.-Y.; Tran, S.; Simmonds, R. Functional characterization of domains found within a lytic enzyme produced by Streptococcus equi subsp. zooepidemicus. FEMS Microbiol. Lett. 2002, 215, 133–138. [Google Scholar] [CrossRef]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Lee, H.G.; Kwon, I.; Seo, J. Characterization of Endolysin LyJH307 with Antimicrobial Activity against Streptococcus bovis. Animals 2020, 10, 963. https://doi.org/10.3390/ani10060963
Kim H, Lee HG, Kwon I, Seo J. Characterization of Endolysin LyJH307 with Antimicrobial Activity against Streptococcus bovis. Animals. 2020; 10(6):963. https://doi.org/10.3390/ani10060963
Chicago/Turabian StyleKim, Hanbeen, Hyo Gun Lee, Inhyuk Kwon, and Jakyeom Seo. 2020. "Characterization of Endolysin LyJH307 with Antimicrobial Activity against Streptococcus bovis" Animals 10, no. 6: 963. https://doi.org/10.3390/ani10060963
APA StyleKim, H., Lee, H. G., Kwon, I., & Seo, J. (2020). Characterization of Endolysin LyJH307 with Antimicrobial Activity against Streptococcus bovis. Animals, 10(6), 963. https://doi.org/10.3390/ani10060963