Marked Presence of Methicillin-Resistant Staphylococcus aureus in Wild Lagomorphs in Valencia, Spain

 , , , and
, , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling, Isolation and Characterization of S. aureus Isolates
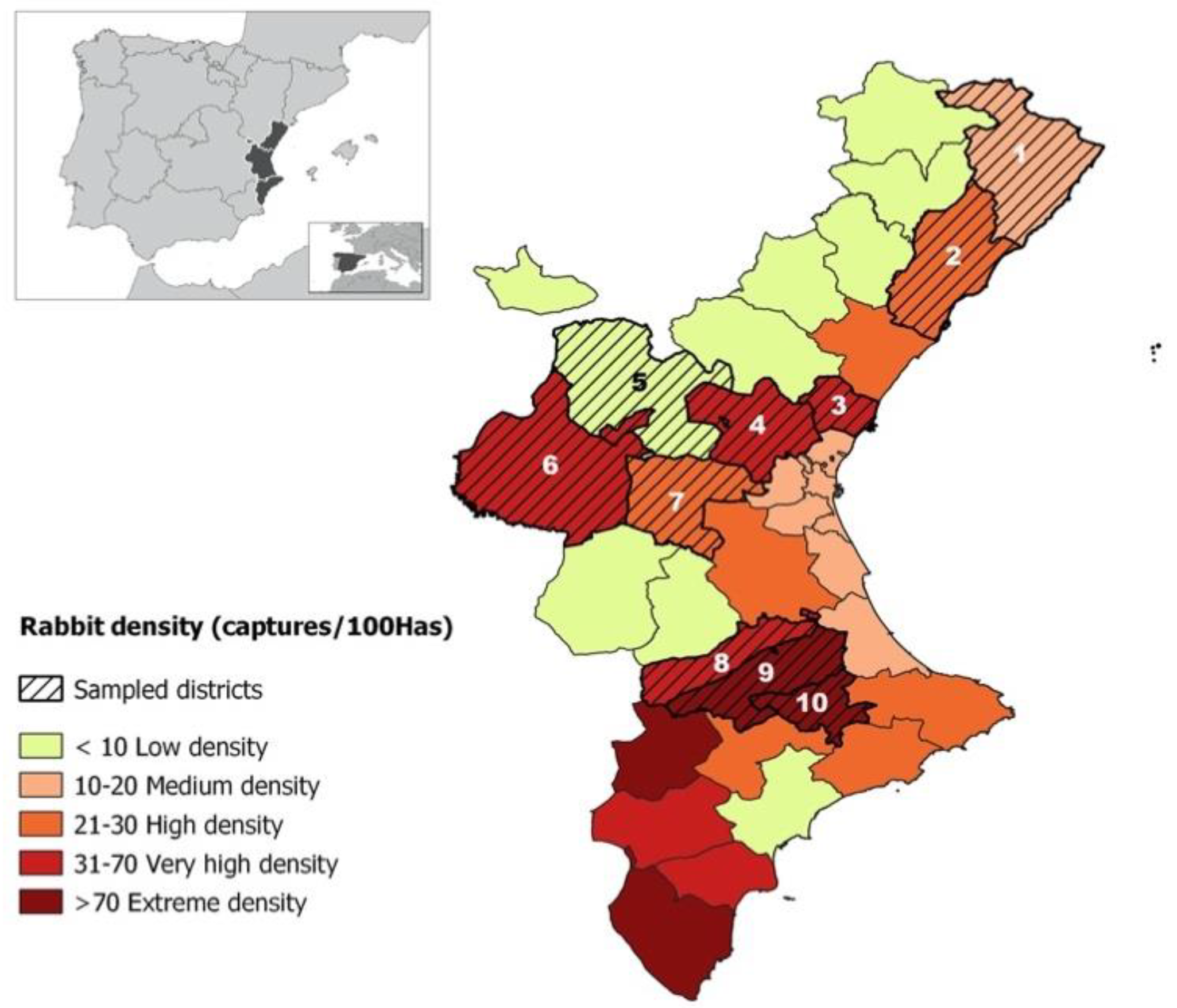
2.1.1. Animals and Geographical Locations
2.1.2. Microbiological Studies
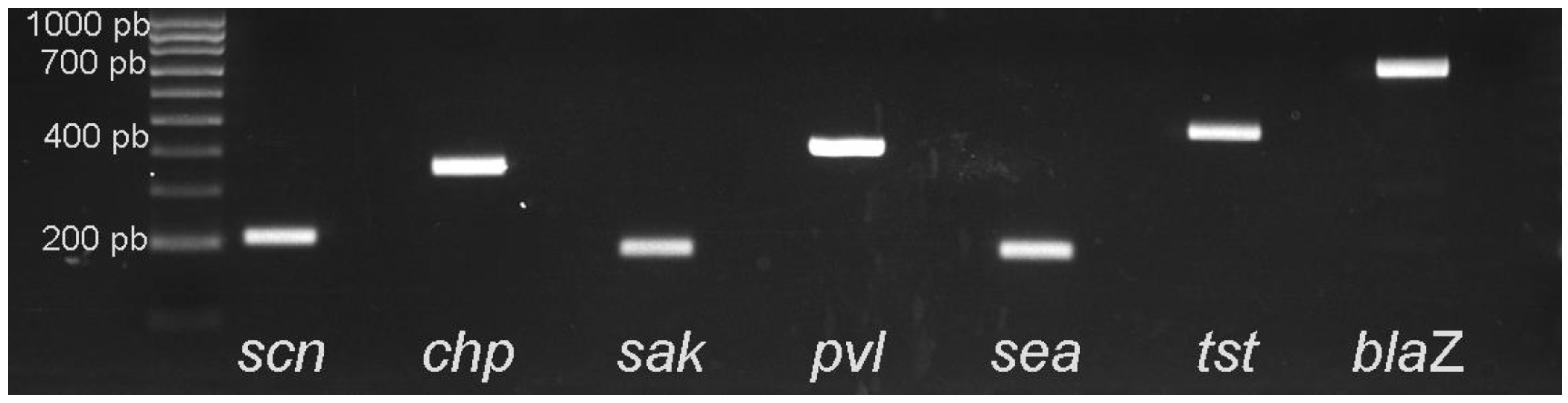
2.1.3. Detection of chp, sak, scn, blaZ, tst, sea, and PVL-encoding Genes, and agr Typing
2.1.4. Antibiotic Susceptibility Testing
2.2. Statistical Analysis
3. Results
3.1. Identification of S. aureus, MRSA and Lesions
3.2. Characterization of S. aureus Isolates
3.3. Antibiotic Resistance Profile
3.4. Detection of the IEC Cluster (scn, chp, sak and sea), blaZ, tst and the PVL Genes among MRSA Isolates
4. Discussion
5. Conclusion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kloos, W.E. Natural populations of the genus Staphylococcus. Annu. Rev. Microbiol. 1980, 34, 559–592. [Google Scholar] [CrossRef]
- Wertheim, H.F.L.; Melles, D.C.; Vos, M.C.; van Leeuwen, W.; van Belkum, A.; Verbrugh, H.A.; Nouwen, J.L. The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect. Dis. 2005, 5, 751–762. [Google Scholar] [CrossRef]
- Burton, S.; Reid-Smith, R.; McClure, J.T.; Weese, J.S. Staphylococcus aureus colonization in healthy horses in Atlantic Canada. Can. Vet. J. 2008, 49, 797–799. [Google Scholar]
- Vautor, E.; Abadie, G.; Guibert, J.-M.; Chevalier, N.; Pépin, M. Nasal carriage of Staphylococcus aureus in dairy sheep. Vet. Microbiol. 2005, 106, 235–239. [Google Scholar] [CrossRef] [Green Version]
- Hermans, K.; De Herdt, P.; Devriese, L.A.; Godard, C.; Haesebrouck, F. Colonisation of rabbits with Staphylococcus aureus after experimental infection with high and low virulence strains. Vet. Microbiol. 2000, 72, 277–284. [Google Scholar] [CrossRef]
- Selva, L.; Viana, D.; Corpa, J.M. Staphylococcus aureus nasal carriage could be a risk for development of clinical infections in rabbits. World Rabbit. Sci. 2015, 23, 181–184. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.N. Global epidemiology of antimicrobial resistance among community-acquired and nosocomial pathogens: A five-year summary from the SENTRY Antimicrobial Surveillance Program (1997–2001). Semin. Respir. Crit. Care Med. 2003, 24, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Francois, P.; Huyghe, A.; Charbonnier, Y.; Bento, M.; Herzig, S.; Topolski, I.; Fleury, B.; Lew, D.; Vaudaux, P.; Harbarth, S.; et al. Use of an automated multiple-locus, variable-number tandem repeat-based method for rapid and high-throughput genotyping of Staphylococcus aureus isolates. J. Clin. Microbiol. 2005, 43, 3346–3355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura, P.; Martinez, J.; Peris, B.; Selva, L.; Viana, D.; Penadés, J.R.; Corpa, J.M. Staphylococcal infections in rabbit does on two industrial farms. Vet. Rec. 2007, 160, 869–872. [Google Scholar] [CrossRef]
- Rosell, J.M.; de la Fuente, L.F. Culling and mortality in breeding rabbits. Prev. Vet. Med. 2009, 88, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Vancraeynest, D.; Haesebrouck, F.; Deplano, A.; Denis, O.; Godard, C.; Wildemauwe, C.; Hermans, K. International dissemination of a high virulence rabbit Staphylococcus aureus clone. J. Vet. Med. B Infect. Dis. Vet. Public Health 2006, 53, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Viana, D.; Selva, L.; Callanan, J.J.; Guerrero, I.; Ferrian, S.; Corpa, J.M. Strains of Staphylococcus aureus and pathology associated with chronic suppurative mastitis in rabbits. Vet. J. 2011, 190, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Pantosti, A.; Sanchini, A.; Monaco, M. Mechanisms of antibiotic resistance in Staphylococcus aureus. Fut. Microbiol. 2007, 2, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.J. Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiol. Rev. 2017, 41, 430–449. [Google Scholar] [CrossRef]
- Cuny, C.; Wieler, L.; Witte, W. Livestock-associated MRSA: The impact on humans. Antibiotics 2015, 4, 521–543. [Google Scholar] [CrossRef]
- Armand-Lefevre, L.; Ruimy, R.; Andremont, A. Clonal comparison of Staphylococcus aureus isolates from healthy pig farmers, human controls, and pigs. Emerg. Infect. Dis. 2005, 11, 711–714. [Google Scholar] [CrossRef]
- Graveland, H.; Duim, B.; van Duijkeren, E.; Heederik, D.; Wagenaar, J.A. Livestock-associated methicillin-resistant Staphylococcus aureus in animals and humans. Int. J. Med. Microbiol. 2011, 301, 630–634. [Google Scholar] [CrossRef]
- Frana, T.S.; Beahm, A.R.; Hanson, B.M.; Kinyon, J.M.; Layman, L.L.; Karriker, L.A.; Ramirez, A.; Smith, T.C. Isolation and characterization of methicillin-resistant Staphylococcus aureus from pork farms and visiting veterinary students. PLoS ONE 2013, 8, e53738. [Google Scholar] [CrossRef] [Green Version]
- Aires-de-Sousa, M. Methicillin-resistant Staphylococcus aureus among animals: Current overview. Clin. Microbiol. Infect. 2017, 23, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Angen, Ø.; Feld, L.; Larsen, J.; Rostgaard, K.; Skov, R.; Madsen, A.M.; Larsen, A.R. Transmission of methicillin-resistant Staphylococcus aureus to human volunteers visiting a swine farm. Appl. Environ. Microbiol. 2017, 83, e01489-17. [Google Scholar] [CrossRef] [Green Version]
- Agnoletti, F.; Mazzolini, E.; Bacchin, C.; Bano, L.; Berto, G.; Rigoli, R.; Muffato, G.; Coato, P.; Tonon, E.; Drigo, I. First reporting of methicillin-resistant Staphylococcus aureus (MRSA) ST398 in an industrial rabbit holding and in farm-related people. Vet. Microbiol. 2014, 170, 172–177. [Google Scholar] [CrossRef]
- Moreno-Grúa, E.; Pérez-Fuentes, S.; Muñoz-Silvestre, A.; Viana, D.; Fernández-Ros, A.B.; Sanz-Tejero, C.; Corpa, J.M.; Selva, L. Characterization of livestock-associated methicillin-resistant Staphylococcus aureus isolates obtained from commercial rabbitries located in the Iberian Peninsula. Front. Microbiol. 2018, 9, 1812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantosti, A. Methicillin-resistant Staphylococcus aureus associated with animals and its relevance to human health. Front. Microbiol. 2012, 3, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monecke, S.; Slickers, P.; Ellington, M.J.; Kearns, A.M.; Ehricht, R. High diversity of Panton-Valentine leukocidin-positive, methicillin-susceptible isolates of Staphylococcus aureus and implications for the evolution of community-associated methicillin-resistant S. aureus. Clin. Microbiol. Infect. 2014, 13, 1157–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mat Azis, N.; Pung, H.P.; Abdul Rachman, A.R.; Amin Nordin, S.; Sarchio, S.N.E.; Suhaili, Z.; Mohd Desa, M.N. A persistent antimicrobial resistance pattern and limited methicillin-resistance-associated genotype in a short-term Staphylococcus aureus carriage isolated from a student population. J. Infect. Public Health 2017, 10, 156–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowder, B.V.; Guinane, C.M.; Ben Zakour, N.L.; Weinert, L.A.; Conway-Morris, A.; Cartwright, R.A.; Simpson, A.J.; Rambaut, A.; Nübel, U.; Fitzgerald, J.R. Recent human-to-poultry host jump, adaptation, and pandemic spread of Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2009, 106, 19545–19550. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.C. Livestock-associated Staphylococcus aureus: The United States experience. PLoS Pathog. 2015, 11, e1004564. [Google Scholar] [CrossRef]
- Dolejska, M.; Cizek, A.; Literak, I. High prevalence of antimicrobial-resistant genes and integrons in Escherichia coli isolates from Black-headed Gulls in the Czech Republic. J. Appl. Microbiol. 2007, 103, 11–19. [Google Scholar] [CrossRef]
- Ruiz Ripa, L.; Alcalá, L.; Simón, C.; Gómez, P.; Mama, O.M.; Rezusta, A.; Zarazaga, M.; Torres, C. Diversity of Staphylococcus aureus clones in wild mammals in Aragon, Spain, with detection of MRSA ST130-mecCin wild rabbits. J. Appl. Microbiol. 2019, 127, 284–291. [Google Scholar] [CrossRef]
- Loncaric, I.; Kubber-Heiss, A.; Posautz, A.; Stalder, G.L.; Hoffmann, D.; Rosengarten, R.; Walzer, C. Characterization of methicillin-resistant Staphylococcus spp. carrying the mecC gene, isolated from wildlife. J. Antimicrob. Chemother. 2013, 2. [Google Scholar] [CrossRef] [Green Version]
- Monecke, S.; Gavier-Widén, D.; Hotzel, H.; Peters, M.; Guenther, S.; Lazaris, A.; Loncaric, I.; Müller, E.; Reissig, A.; Ruppelt-Lorz, A.; et al. Diversity of Staphylococcus aureus isolates in European wildlife. PLoS ONE 2016, 11, e0168433. [Google Scholar] [CrossRef] [Green Version]
- DOGV (Diario Oficial Comunidad Valenciana [Valencian Region Official Gazette]). Resolution of November 20, 2018, of the Director General of Natural Environment and Environmental Assessment, which Updates the Annex of the Order of June 11, 2009, of the Ministry of Environment, Water, Urban Planning and Housing, which Approves Extraordinary Guidelines for the Use, Management and Control of the Mountain Rabbit in the Valencian Region, Relative to the List of Municipal Terms Affected by Overpopulation of Mountain Rabbit. DOGV No. 8437 from 12.04.2018. 2018. Available online: http://www.dogv.gva.es/portal/ficha_disposicion.jsp?L=1&sig=010983%2F2018 (accessed on 25 February 2020).
- Torres, C.; Esteve, C.; Nieto, M.P.; Burgui, J.M. Estadísticas cinegéticas de la Comunitat Valenciana. Memoria 2018. Temporada 2017/2018 [Hunting statistics of the Valencian Community. Annual Report 2018. 2017/2018 Season]. Hunting and Fishing Service; Valencia Region Government: Valencia, Spain, 2019. [Google Scholar]
- Devriese, L.A.; Hendrickx, W.; Godard, C.; Okerman, L.; Haesebrouck, F. A new pathogenic Staphylococcus aureus type in commercial rabbits. Zentralbl Veterinarmed B 1996, 43, 313–315. [Google Scholar] [CrossRef] [PubMed]
- Viana, D.; Selva, L.; Segura, P.; Penadés, J.R.; Corpa, J.M. Genotypic characterization of Staphylococcus aureus strains isolated from rabbit lesions. Vet. Microbiol. 2007, 121, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geha, D.J.; Uhl, J.R.; Gustaferro, C.A.; Persing, D.H. Multiplex PCR for identification of methicillin-resistant staphylococci in the clinical laboratory. J. Clin. Microbiol. 1994, 32, 1768–1772. [Google Scholar] [CrossRef] [Green Version]
- Khairalla, A.S.; Wasfi, R.; Ashour, H.M. Carriage frequency, phenotypic, and genotypic characteristics of methicillin-resistant Staphylococcus aureus isolated from dental health-care personnel, patients, and environment. Sci. Rep. 2017, 7, 7390. [Google Scholar] [CrossRef] [PubMed]
- Viana, D.; Selva, L.; Penadés, M.; Corpa, J.M. Screening of virulence genes in Staphylococcus aureus isolates from rabbits. World Rabbit Sci. 2015, 23, 185. [Google Scholar] [CrossRef] [Green Version]
- van Wamel, W.J.B.; Rooijakkers, S.H.M.; Ruyken, M.; van Kessel, K.P.M.; van Strijp, J.A.G. The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on β-hemolysin-converting bacteriophages. J. Bacteriol. 2006, 188, 1310–1315. [Google Scholar] [CrossRef] [Green Version]
- Schnellmann, C.; Gerber, V.; Rossano, A.; Jaquier, V.; Panchaud, Y.; Doherr, M.G.; Thomann, A.; Straub, R.; Perreten, V. Presence of new mecA and mph(C) variants conferring antibiotic resistance in Staphylococcus spp. isolated from the skin of horses before and after clinic admission. J. Clin. Microbiol. 2006, 44, 4444–4454. [Google Scholar] [CrossRef] [Green Version]
- Hermans, K.; De Herdt, P.; Devriese, L.A.; Hendrickx, W.; Godard, C.; Haesebrouck, F. Colonization of rabbits with Staphylococcus aureus in flocks with and without chronic staphylococcosis. Vet. Microbiol. 1999, 67, 37–46. [Google Scholar] [CrossRef]
- Corpa, J.M.; Hermans, K.; Haesebrouck, F. Main pathologies associated with Staphylococcus aureus infections in rabbits: A review. World Rabbit Sci. 2010, 17, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, I.; Ferrian, S.; Penadés, M.; García-Quirós, A.; Pascual, J.J.; Selva, L.; Viana, D.; Corpa, J.M. Host responses associated with chronic staphylococcal mastitis in rabbits. Vet. J. 2015, 204, 338–344. [Google Scholar] [CrossRef]
- Ariza-Miguel, J.; Hernández, M.; Fernández-Natal, I.; Rodríguez-Lázaro, D. Methicillin-resistant Staphylococcus aureus harboring mecC in livestock in Spain. J. Clin. Microbiol. 2014, 52, 4067–4069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacinti, G.; Carfora, V.; Caprioli, A.; Sagrafoli, D.; Marri, N.; Giangolini, G.; Amoruso, R.; Iurescia, M.; Stravino, F.; Dottarelli, S.; et al. Prevalence and characterization of methicillin-resistant Staphylococcus aureus carrying mecA or mecC and methicillin-susceptible Staphylococcus aureus in dairy sheep farms in central Italy. J. Dairy Sci. 2017, 100, 7857–7863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paterson, G.K.; Larsen, A.R.; Robb, A.; Edwards, G.E.; Pennycott, T.W.; Foster, G.; Mot, D.; Hermans, K.; Baert, K.; Peacock, S.J.; et al. The newly described mecA homologue, mecALGA251, is present in methicillin-resistant Staphylococcus aureus isolates from a diverse range of host species. J. Antimicrob. Chemother. 2012, 67, 2809–2813. [Google Scholar] [CrossRef]
- Meemken, D.; Blaha, T.; Hotzel, H.; Strommenger, B.; Klein, G.; Ehricht, R.; Monecke, S.; Kehrenberg, C. Genotypic and phenotypic characterization of Staphylococcus aureus isolates from wild boars. Appl. Environ. Microbiol. 2013, 79, 1739–1742. [Google Scholar] [CrossRef] [Green Version]
- Porrero, M.C.; Mentaberre, G.; Sánchez, S.; Fernández-Llario, P.; Casas-Díaz, E.; Mateos, A.; Vidal, D.; Lavín, S.; Fernández-Garayzábal, J.-F.; Domínguez, L. Carriage of Staphylococcus aureus by free-living wild animals in Spain. Appl. Environ. Microbiol. 2014, 80, 4865–4870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Álvarez, L.; Holden, M.T.G.; Lindsay, H.; Webb, C.R.; Brown, D.F.J.; Curran, M.D.; Walpole, E.; Brooks, K.; Pickard, D.J.; Teale, C.; et al. Meticillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: A descriptive study. Lancet Infect. Dis. 2011, 11, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Porrero, M.C.; Valverde, A.; Fernández-Llario, P.; Díez-Guerrier, A.; Mateos, A.; Lavín, S.; Cantón, R.; Fernández-Garayzábal, J.-F.; Domínguez, L. Staphylococcus aureus carrying mecC gene in animals and urban wastewater, Spain. Emerg. Infect. Dis. 2014, 20, 899–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merz, A.; Stephan, R.; Johler, S. Genotyping and DNA microarray based characterization of Staphylococcus aureus isolates from rabbit carcasses. Meat Sci. 2016, 112, 86–89. [Google Scholar] [CrossRef]
- Cuny, C.; Layer, F.; Strommenger, B.; Witte, W. Rare occurrence of methicillin-resistant Staphylococcus aureus CC130 with a novel mecA homologue in humans in Germany. PLoS ONE 2011, 6, e24360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurent, F.; Chardon, H.; Haenni, M.; Bes, M.; Reverdy, M.-E.; Madec, J.-Y.; Lagier, E.; Vandenesch, F.; Tristan, A. MRSA harboring mecA variant gene mecC, France. Emerg. Infect. Dis. 2012, 18, 1465–1467. [Google Scholar] [CrossRef] [PubMed]
- García-Garrote, F.; Cercenado, E.; Marín, M.; Bal, M.; Trincado, P.; Corredoira, J.; Ballesteros, C.; Pita, J.; Alonso, P.; Vindel, A. Methicillin-resistant Staphylococcus aureus carrying the mecC gene: Emergence in Spain and report of a fatal case of bacteraemia. J. Antimicrob. Chemother. 2014, 69, 45–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, P.; González-Barrio, D.; Benito, D.; García, J.T.; Viñuela, J.; Zarazaga, M.; Ruiz-Fons, F.; Torres, C. Detection of methicillin-resistant Staphylococcus aureus (MRSA) carrying the mecC gene in wild small mammals in Spain. J. Antimicrob. Chemother. 2014, 69, 2061–2064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witte, W.; Strommenger, B.; Stanek, C.; Cuny, C. Methicillin-resistant Staphylococcus aureus ST398 in humans and animals, Central Europe. Emerg. Infect. Dis. 2007, 13, 255–258. [Google Scholar] [CrossRef]
- Bardiau, M.; Yamazaki, K.; Duprez, J.-N.; Taminiau, B.; Mainil, J.G.; Ote, I. Genotypic and phenotypic characterization of methicillin-resistant Staphylococcus aureus (MRSA) isolated from milk of bovine mastitis. Lett. Appl. Microbiol. 2013, 57, 181–186. [Google Scholar] [CrossRef]
- Paterson, G.K.; Morgan, F.J.E.; Harrison, E.M.; Cartwright, E.J.P.; Török, M.E.; Zadoks, R.N.; Parkhill, J.; Peacock, S.J.; Holmes, M.A. Prevalence and characterization of human mecC methicillin-resistant Staphylococcus aureus isolates in England. J. Antimicrob. Chemother. 2014, 69, 907–910. [Google Scholar] [CrossRef] [Green Version]
- Drougka, E.; Foka, A.; Posantzis, D.; Giormezis, N.; Anastassiou, E.D.; Petinaki, E.; Spiliopoulou, I. Human Staphylococcus aureus lineages among Zoological Park residents in Greece. Open Vet. J. 2015, 5, 148–153. [Google Scholar]
- Viana, D.; Comos, M.A.; McAdam, P.R.; Ward, M.J.; Selva, L.; Guinane, C.M.; Gonzalez-Munoz, B.M.; Tristan, A.; Foster, S.J.; Fitzgerald, J.R.; et al. A single natural nucleotide mutation alters bacterial pathogen host tropism. Nat. Genet. 2015, 47, 361–366. [Google Scholar] [CrossRef]
- Harrison, E.M.; Coll, F.; Toleman, M.S.; Blane, B.; Brown, N.M.; Török, M.E.; Parkhill, J.; Peacock, S.J. Genomic surveillance reveals low prevalence of livestock-associated methicillin-resistant Staphylococcus aureus in the East of England. Sci. Rep. 2017, 7, 7406–7407. [Google Scholar] [CrossRef] [Green Version]
- Traversa, A.; Gariano, G.R.; Gallina, S.; Bianchi, D.M.; Orusa, R.; Domenis, L.; Cavallerio, P.; Fossati, L.; Serra, R.; Decastelli, L. Methicillin resistance in Staphylococcus aureus strains isolated from food and wild animal carcasses in Italy. Food Microbiol. 2015, 52, 154–158. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Primer | Sequence (5′–3′) | References | |
|---|---|---|---|
| mecA | mecA-1m | GTAGAAATGACTGAACGTCCGATAA | [37,38] |
| mecA-2c | CCAATTCCACATTGTTTCGGTCTAA | ||
| mecC | mecC-1m | CTACACGTTCCATACCATTAG | [37,38] |
| mecC-2c | CGCCTCTATGATAAACAATTGC | ||
| lukF/S-PV | lukF/S-PV-1m | ATCATTAGGTAAAATGTCTGGACATGATCCA | [39] |
| lukF/S-PV-2c | GCATCAAGTGTATTGGATAGCAAAAGC | ||
| sea | sea-1m | AAAGTCCCGATCAATTTATGGCTA | [39] |
| sea-2c | GTAATTAACCGAAGGTTCTGTAGA | ||
| tst | tst-1m | CTAATCAAATAATCAAAACTGC | [39] |
| tst-2c | TTTCCAATAACCACCCGTTT | ||
| agr type I | agr type I-1m | GTCACAAGTACTATAAGCTGCGAT | [39] |
| agr type I-2c | ATGCACATGGTGCACATGC | ||
| agr type II | agr type II-1m | TATTACTAATTGAAAAGTGGCCATAGC | [39] |
| agr type II-2c | ATGCACATGGTGCACATGC | ||
| agr type III | agr type III-1m | GTAATGTAATAGCTTGTATAATAATACCCAG | [39] |
| agr type III-2c | ATGCACATGGTGCACATGC | ||
| agr type IV | agr type IV-1m | CGATAATGCCGTAATACCCG | [39] |
| agr type IV-2c | ATGCACATGGTGCACATGC | ||
| sak | sak-1m | AAGGCGATGACGCGAGTTAT | [40] |
| sak-2c | GCGCTTGGATCTAATTCAAC | ||
| chp | chp-1m | TTTACTTTTGAACCGTTTCCTAC | [40] |
| chp-2c | CGTCCTGAATTCTTAGTATGCATATTCATTAG | ||
| scn | scn-1m | ACTTTAGCAATCGTTTTAGC | [40] |
| scn-2c | CTGAAATTTTTATAGTTCGC | ||
| blaZ | blaZ-1m | CAGTTCACATGCCAAAGAG | [41] |
| blaZ-2c | TACACTCTTGGCGGTTTC | ||
| Id | District | Town | No. of Animals Tested | No. of Animals S. aureus Positive 1 | p2 | Animals mecC+ | Animals mecA+ | mec (%) |
|---|---|---|---|---|---|---|---|---|
| 1 | El Baix Maestrat | Vinaroz | 1 | 1 | 0.4132 | 1 | 0 | 100 |
| 2 | La Plana Alta | Cabanes | 120 | 18 | 0.0001 | 10 | 0 | 55.6 |
| 3 | Camp de Morvedre | Faura | 31 | 9 | 0.1826 | 1 | 0 | 11.1 |
| 4 | El Camp de Túria | Llíria | 1 | 0 | 1 | 0 | 0 | 0 |
| 4 | El Camp de Túria | Vilamarxant | 2 | 1 | 1 | 1 | 0 | 100 |
| 5 | Serranos | Pedralba | 39 | 37 | 0.0001 | 32 | 0 | 86.5 |
| 6 | La Plana Utiel-Requena | Requena | 2 | 0 | 0.5137 | 0 | 0 | 0 |
| 6 | La Plana Utiel-Requena | Utiel | 32 | 6 | 0.0077 | 0 | 0 | 0 |
| 7 | Hoya de Buñol | Cheste | 53 | 13 | 0.0098 | 8 | 1 | 69.2 |
| 7 | Hoya de Buñol | Godelleta | 15 | 11 | 0.0143 | 4 | 0 | 36.4 |
| 8 | La Costera | Montesa | 1 | 0 | 1 | 0 | 0 | 0 |
| 9 | La Vall d’Albaida | Castelló de Rugat | 28 | 26 | 0.0001 | 15 | 0 | 57.7 |
| 9 | La Vall d’Albaida | Montaverner | 18 | 9 | 0.4695 | 3 | 0 | 33.3 |
| 10 | El Comtat | Alfafara | 20 | 19 | 0.0001 | 19 | 0 | 100 |
| Total | 363 | 150 | 94 | 1 | 63.3 | |||
| Sampling Localization | Samples | S. aureus | (%) | MRSA | (%) |
|---|---|---|---|---|---|
| Nasal cavity | 363 | 70 | 19.3 | 34 | 48.6 |
| Ears | 363 | 121 | 33.3 | 80 | 66.1 |
| Perineum | 363 | 45 | 12.4 | 20 | 44.4 |
| Lesions | 11 | 8 | 72.7 | 6 | 75 |
| Dermatitis | 6 | 5 | 83.3 | 3 | 60 |
| Hepatic abscesses | 2 | 2 | 100 | 2 | 100 |
| Conjunctivitis | 3 | 1 | 33.3 | 1 | 100 |
| Total | 1100 | 244 | 22.2 | 140 | 57.4 |
| MLST | CC | MRSA | MSSA | Total |
|---|---|---|---|---|
| ST1945 | CC130 | 136 | 41 | 177 |
| ST425 | CC425 | - | 42 | 42 |
| ST121 | CC121 | - | 7 | 7 |
| ST5821 | CC425 | - | 4 | 4 |
| ST5826 | singleton | - | 3 | 3 |
| ST398 | CC398 | 1 | 2 | 3 |
| ST5 | CC5 | - | 2 | 2 |
| ST5822 | CC130 | 1 | - | 1 |
| ST5823 | CC130 | 1 | - | 1 |
| ST5824 | singleton | 1 | - | 1 |
| ST5825 | CC130 | - | 1 | 1 |
| ST5844 | CC425 | - | 1 | 1 |
| ST5845 | CC130 | - | 1 | 1 |
| Total | 140 | 104 | 244 |
| Lesions | Rabbits | CC Lesion | CC Nostril | CC Ear | CC Perineum |
|---|---|---|---|---|---|
| Dermatitis | OC19006 | CC130/CC425 | CC425 | - | CC425 |
| Conjunctivitis | OC19190 | CC130 | CC130 | CC130 | CC130 |
| Hepatitis | OC19275 | CC130 | CC130 | CC130 | CC130 |
| Hepatitis | OC19276 | CC130 | CC130 | CC130 | CC130 |
| Strain | Town | Anatomical Location | Molecular Typing | SCCmec | IEC | Antimicrobial Resistance Phenotype | Antimicrobial Resistance Genes | |||
|---|---|---|---|---|---|---|---|---|---|---|
| coa/spa | MLST | CC | agr | |||||||
| 1439 | Alfafara | Ear | B4 I1 | ST1945 | CC130 | III | XI | scn-sak (group E) | PEN-FOX | mecC-SCCmecXI |
| 1490 | Cabanes | Dermatitis | B4 I1 | ST5822 | CC130 | III | XI | - | PEN | mecC-SCCmecXI |
| 1564 | Cabanes | Nostril | B4 I1 | ST1945 | CC130 | III | XI | - | Susceptible | mecC-SCCmecXI |
| 1768 | Castelló de Rugat | Ear | B4 I1 | ST1945 | CC130 | III | XI | - | PEN-FOX | mecC-SCCmecXI |
| 1657 | Cheste | Ear | D1 I6 | ST398 | CC398 | I | V | - | PEN-FOX-ENO-TET | mecA-blaZ-SCCmecV |
| 1660 | Cheste | Ear | B4 I1 | ST5823 | CC130 | III | XI | - | PEN-FOX | mecC-SCCmecXI |
| 1651 | Cheste | Perineum | B4 I1 | ST1945 | CC130 | III | XI | - | PEN-FOX | mecC-SCCmecXI |
| 1867 | Faura | Ear | B4 II4 | ST1945 | CC130 | III | XI | - | FOX | mecC-SCCmecXI |
| 1707 | Godelleta | Nostril | B4 I1 | ST1945 | CC130 | III | XI | - | PEN-FOX | mecC-SCCmecXI |
| 1712 | Godellerta | Ear | A1 IV1 | ST5824 | singleton | II | XI | - | PEN-FOX | mecC-SCCmecXI |
| 1620 | Montaverner | Ear | B4 I1 | ST1945 | CC130 | III | XI | - | PEN-FOX | mecC-SCCmecXI |
| 1468 | Pedralba | Perineum | B4 II4 | ST1945 | CC130 | III | XI | - | PEN-FOX | mecC-SCCmecXI |
| 1802 | Vilamarxant | Nostril | B4 I1 | ST1954 | CC130 | III | XI | - | PEN-FOX | mecC-SCCmecXI |
| 1272 | Vinaròs | Dermatitis | B4 I1 | ST1945 | CC130 | III | XI | scn-sak (group E) | PEN-FOX | mecC-SCCmecXI |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Grúa, E.; Pérez-Fuentes, S.; Viana, D.; Cardells, J.; Lizana, V.; Aguiló, J.; Selva, L.; Corpa, J.M. Marked Presence of Methicillin-Resistant Staphylococcus aureus in Wild Lagomorphs in Valencia, Spain. Animals 2020, 10, 1109. https://doi.org/10.3390/ani10071109
Moreno-Grúa E, Pérez-Fuentes S, Viana D, Cardells J, Lizana V, Aguiló J, Selva L, Corpa JM. Marked Presence of Methicillin-Resistant Staphylococcus aureus in Wild Lagomorphs in Valencia, Spain. Animals. 2020; 10(7):1109. https://doi.org/10.3390/ani10071109
Chicago/Turabian StyleMoreno-Grúa, Elena, Sara Pérez-Fuentes, David Viana, Jesús Cardells, Víctor Lizana, Jordi Aguiló, Laura Selva, and Juan M. Corpa. 2020. "Marked Presence of Methicillin-Resistant Staphylococcus aureus in Wild Lagomorphs in Valencia, Spain" Animals 10, no. 7: 1109. https://doi.org/10.3390/ani10071109
APA StyleMoreno-Grúa, E., Pérez-Fuentes, S., Viana, D., Cardells, J., Lizana, V., Aguiló, J., Selva, L., & Corpa, J. M. (2020). Marked Presence of Methicillin-Resistant Staphylococcus aureus in Wild Lagomorphs in Valencia, Spain. Animals, 10(7), 1109. https://doi.org/10.3390/ani10071109