Behavioral Diversity as a Potential Indicator of Positive Animal Welfare




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Behavioral Diversity and Behavioral Restriction
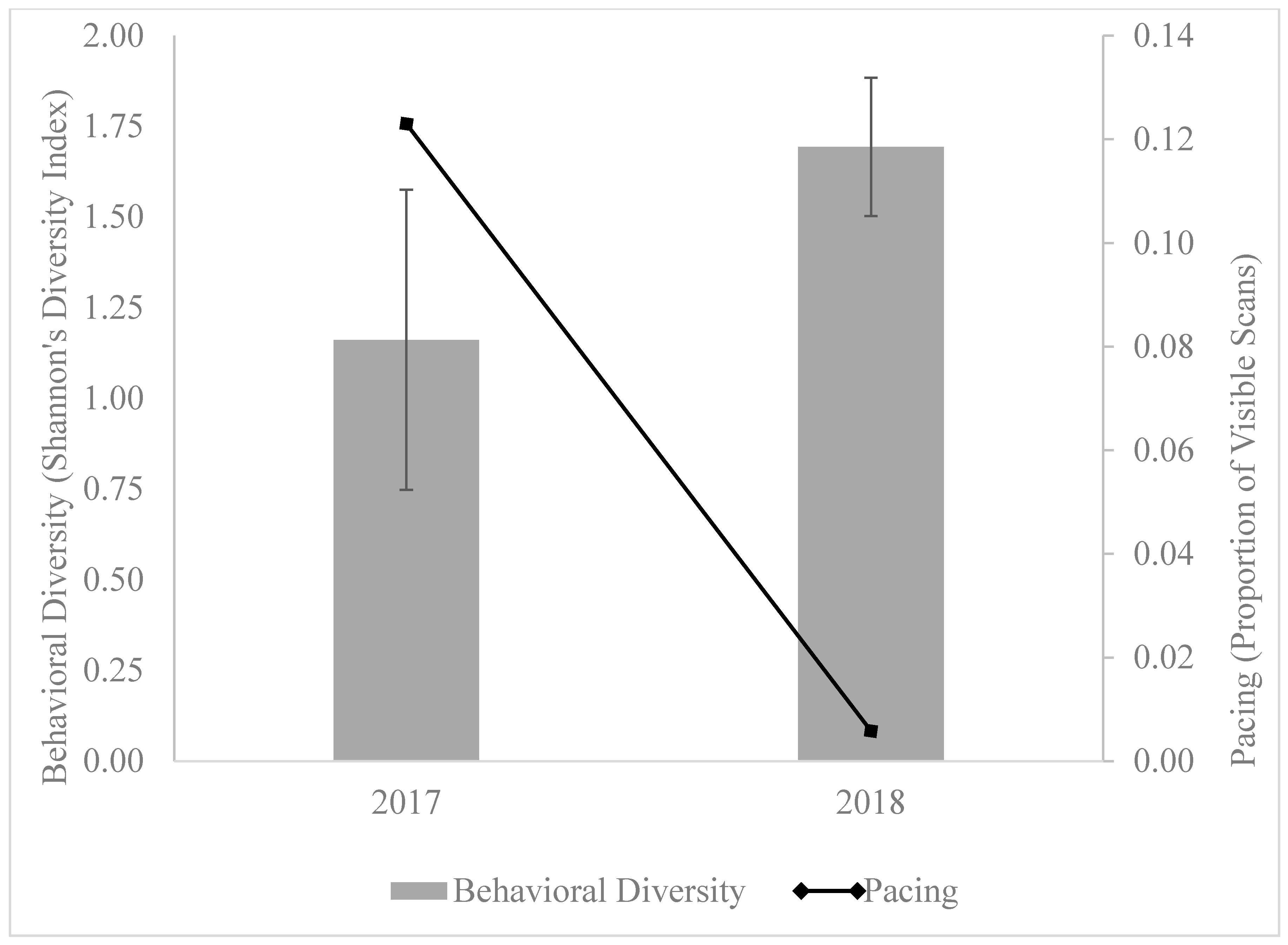
4. Behavioral Diversity and Stereotypic Behavior
5. Factors Affecting Behavioral Diversity
6. Behavioral Diversity and Physiological Indicators of Animal Welfare
7. Methods of Calculation
8. Behavioral Diversity Considerations
9. Shannon’s Behavioral Diversity Index Scores across Species
10. Application of Behavioral Diversity
- Does the animal express behavior at a frequency and diversity that is consistent with natural history?
- Does the animal express normal social behavior?
- Does the animal respond in a behaviorally appropriate way to challenges, problem solving, and environmental changes in a way that is consistent with natural history?
- Is the animal responsive to learning new skills and shows motivation to engage?
- Does the animal use the environment to acquire resources that benefit them?
11. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Melfi, V.A. There are big gaps in our knowledge, and thus approach, to zoo animal welfare: A case for evidence-based zoo animal management. Zoo Biol. 2009, 28, 574–588. [Google Scholar] [CrossRef]
- Webster, A.J.F. Animal Welfare: A Cool Eye Towards Eden; Blackwell Science: Oxford, UK, 1995. [Google Scholar]
- Anonymous. Five Freedoms; Farm Animal Welfare Council, the National Archives, 2012. Available online: http://webarchive.nationalarchives.gov.uk/20121007104210/http:/www.fawc.org.uk/freedoms.htm (accessed on 19 May 2020).
- Mellor, D.J.; Reid, C.S.W. Concepts of animal well-being and predicting the impact of procedures on experimental animals. In Improving the Well-Being of Animals in the Research Environment; Australian and New Zealand Council for the Care of Animals in Research and Teaching: Glen Osmond, Australia, 1994; pp. 3–18. [Google Scholar]
- Mellor, D.J.; Beausoleil, N.J. Extending the Five Domains model for animal welfare assessment to incorporate positive welfare states. Anim. Welf. 2015, 24, 241–253. [Google Scholar] [CrossRef]
- Vicino, G.; Miller, L.J. From Prevention of Cruelty to Optimizing Welfare: Opportunities to Thrive. In Proceedings of the International Ethological Conference, Cairns, Australia, 19 September 2015. [Google Scholar]
- Miller, L.J.; Pisacane, C.B.; Vicino, G.A. Relationship between behavioural diversity and faecal glucocorticoid metabolites: A case study with cheetahs (Acinonyx jubatus). Anim. Welf. 2016, 25, 325–329. [Google Scholar] [CrossRef]
- Mason, G.J.; Latham, N.R. Can’t stop, won’t stop: Is stereotypy a reliable animal welfare indicator? Anim. Welf. 2004, 13, 57–69. [Google Scholar]
- Benn, A.L.; McLelland, D.J.; Whittaker, A.L. A review of welfare assessment methods in reptiles, and preliminary application of the welfare quality® protocol to the pygmy blue-tongue skink, Tiliqua adelaidensis, using animal-based measures. Animals 2019, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- Brereton, J.E. Directions in animal enclosure use studies. J. Zoo Aquar. Res. 2020, 8, 1–9. [Google Scholar]
- Sherwen, S.L.; Hemsworth, L.M.; Beausoleil, N.J.; Embury, A.; Mellor, D.J. An animal welfare risk assessment process for zoos. Animals 2018, 8, 130. [Google Scholar] [CrossRef]
- Mason, G.J.; Burn, C.C. Behavioral restriction. In Animal Welfare, 2nd ed.; Appleby, M.C., Mench, J.A., Olson, L.A.S., Huges, B.O., Eds.; CAB International: Wallingford, UK, 2011; pp. 98–119. [Google Scholar]
- Spinka, M.; Wemelsfelder, F. Environmental challenge and animal agency. In Animal Welfare, 2nd ed.; Appleby, M.C., Mench, J.A., Olson, L.A.S., Huges, B.O., Eds.; CAB International: Wallingford, UK, 2011; pp. 27–44. [Google Scholar]
- Mason, G.J.; Cooper, J.; Clarebrough, C. Frustrations of fur-farmed mink. Nature 2001, 410, 35–36. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, S.; Van der Vegt, B.J.; Lawrence, A.B.; McLean, K.A.; Deans, L.A.; Chirnside, J.; Calvert, S.K. The effect of parity and environmental restriction on behavioural and physiological responses of pre-parturient pigs. Appl. Anim. Behav. Sci. 2001, 71, 203–216. [Google Scholar] [CrossRef]
- Sarrafchi, A.; Blokhuis, H.J. Equine stereotypic behaviors: Causation, occurrence, and prevention. J. Vet. Behav. 2013, 8, 386–394. [Google Scholar] [CrossRef]
- Würbel, H.; Stauffacher, M.; von Holst, D. Stereotypies in laboratory mice—Quantitative and qualitative description of the ontogeny of wire-gnawing and jumping in Zur: ICR and Zur: ICR nu. Ethology 1996, 102, 371–385. [Google Scholar] [CrossRef]
- Mason, G.J.; Turner, M.A. Mechanisms involved in the development and control of stereotypies. In Perspectives in Ethology; Bateson, P.P.G., Klopfer, P.H., Thompson, N.S., Eds.; Plenum Press: London, UK, 1993; pp. 53–85. [Google Scholar]
- Rushen, J.; Lawrence, A.B.; Terlouw, C.E.M. The motivational basis of stereotypies. In Stereotypic Animal Behavior: Fundamentals and Applications to Welfare; Lawrence, A.B., Rushen, J., Eds.; CAB International: Wallingford, UK, 1993; pp. 41–64. [Google Scholar]
- Meehan, C.L.; Garner, J.P.; Mench, J.A. Environmental enrichment and development of cage stereotypy in orange-winged Amazon parrots (Amazona amazonica). Dev. Psychobiol. 2004, 44, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Stolba, A.; Wood-Gush, D.G.; Baker, N. The characterisation of stereotyped behaviour in stalled sows by informational redundancy. Behaviour 1983, 87, 157–182. [Google Scholar] [CrossRef]
- Goswami, S.; Tyagi, P.C.; Malik, P.K.; Pandit, S.J.; Kadivar, R.F.; Fitzpatrick, M.; Mondol, S. Effects of personality and rearing-history on the welfare of captive Asiatic lions (Panthera leo persica). PeerJ 2020, 8, e8425. [Google Scholar] [CrossRef]
- Gunn, D.; Morton, D.B. Inventory of the behaviour of New Zealand White rabbits in laboratory cages. Appl. Anim. Behav. Sci. 1995, 45, 277–292. [Google Scholar] [CrossRef]
- Miller, L.J.; Lauderdale, L.K.; Bryant, J.L.; Mellen, J.D.; Walsh, M.T.; Granger, D.A. Behavioral diversity as a potential positive indicator of animal welfare in bottlenose dolphins. PLoS ONE. under review.
- Vickery, S.; Mason, G. Stereotypic behavior in Asiatic black and Malayan sun bears. Zoo Biol. 2004, 23, 409–430. [Google Scholar] [CrossRef]
- Shepherdson, D.J.; Carlstead, K.; Mellen, J.D.; Seidensticker, J. The influence of food presentation on the behavior of small cats in confined environments. Zoo Biol. 1993, 12, 203–216. [Google Scholar] [CrossRef]
- Antonenko, T.V.; Medvedeva, J.E.; Panchuk, K.A. The influence of olfactory stimulation on the welfare of big cats in captivity. Ukr. J. Ecol. 2017, 7, 134–138. [Google Scholar] [CrossRef]
- Swaisgood, R.R.; White, A.M.; Zhou, X.; Zhang, G.; Lindburg, D.G. How do giant pandas (Ailuropoda melanoleuca) respond to varying properties of enrichments? A comparison of behavioral profiles among five enrichment items. J. Comp. Psychol. 2005, 119, 325–334. [Google Scholar] [CrossRef]
- Swaisgood, R.R.; White, A.M.; Zhou, X.; Zhang, H.; Zhang, G.; Wei, R.; Hare, V.J.; Tepper, E.M.; Lindburg, D.G. A quantitative assessment of the efficacy of an environmental enrichment programme for giant pandas. Anim. Behav. 2001, 61, 447–457. [Google Scholar] [CrossRef]
- Renner, M.J.; Lussier, J.P. Environmental enrichment for the captive spectacled bear (Tremarctos ornatus). Pharm. Biochem Behav 2002, 73, 279–283. [Google Scholar] [CrossRef]
- Neal Webb, S.J.; Hau, J.; Schapiro, S.J. Captive chimpanzee (Pan troglodytes) behavior as a function of space per animal and enclosure type. Am. J. Primatol. 2018, 80, e22749. [Google Scholar] [CrossRef] [PubMed]
- Scott, N.L.; LaDue, C.A. The behavioral effects of exhibit size versus complexity in African elephants: A potential solution for smaller spaces. Zoo Biol. 2019, 38, 448–457. [Google Scholar] [CrossRef]
- Clyvia, A.; Faggioli, A.B.; Cipreste, C.F. Effects of environmental enrichment in a captive pair of Golden Parakeet (Guaruba guarouba, Psittacidae) with abnormal behaviors. Rev. Bras. Ornitol. 2015, 23, 309–314. [Google Scholar] [CrossRef]
- Garner, J.P.; Mason, G.J.; Smith, R. Stereotypic route-tracing in experimentally caged songbirds correlates with general behavioural disinhibition. Anim. Behav. 2003, 66, 711–727. [Google Scholar] [CrossRef]
- Lyon, M.; Robbins, T. The action of central nervous system stimulant drugs: A general theory concering amphetamine effects. Curr. Dev. Psychopharmacol. 1975, 2, 79–163. [Google Scholar]
- Catapani, M.L.; Pires, J.S.; Vasconcellos, A.D. Single-or pair-housed: Which is better for captive southern tamanduas? J. Appl. Anim. Welf. Sci. 2019, 22, 289–297. [Google Scholar] [CrossRef]
- Miller, L.J.; Mellen, J.; Greer, T.; Kuczaj, S.A. The effects of education programmes on Atlantic bottlenose dolphin (Tursiops truncatus) behaviour. Anim. Welf. 2011, 20, 159–172. [Google Scholar]
- Bashaw, M.J.; Gibson, M.D.; Schowe, D.M.; Kucher, A.S. Does enrichment improve reptile welfare? Leopard geckos (Eublepharis macularius) respond to five types of environmental enrichment. Appl. Anim. Behav. Sci. 2016, 184, 150–160. [Google Scholar] [CrossRef]
- Hogan, L.A.; Johnston, S.D.; Lisle, A.; Horsup, A.B.; Janssen, T.; Phillips, C.J. Stereotypies and environmental enrichment in captive southern hairy-nosed wombats, Lasiorhinus latifrons. Appl. Anim. Behav. Sci. 2010, 126, 85–95. [Google Scholar] [CrossRef]
- de Oliveira, C.G.; Miranda, J.G.; Japyassú, H.F.; El-Hani, C.N. Using Zipf–Mandelbrot law and graph theory to evaluate animal welfare. Phys. A 2018, 492, 285–295. [Google Scholar] [CrossRef]
- Kohari, D.; Sunada, A.; Matsui, Y.; Ootaki, A.; Hori, H. Behavioral restriction effects on activity motivation of a captive lion (Panthera leo persica). J. Vet. Behav. 2017, 17, 14–18. [Google Scholar] [CrossRef]
- Izzo, G.N.; Bashaw, M.J.; Campbell, J.B. Enrichment and individual differences affect welfare indicators in squirrel monkeys (Saimiri sciureus). J. Comp. Psychol. 2011, 125, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.J.; Ivy, J.A.; Vicino, G.A.; Schork, I.G. Impacts of natural history and exhibit factors on carnivore welfare. J. Appl. Anim. Welf. Sci. 2019, 22, 188–196. [Google Scholar] [CrossRef]
- Young, R.J. Environmental Enrichment for Captive Animals; Universities Federation for Animal Welfare: Wheathampstead, UK, 2003. [Google Scholar]
- Van Metter, J.E.; Harriger, M.D.; Bolen, R.H. Environmental enrichment utilizing stimulus objects for African lions (Panthera leo leo) and Sumatran tigers (Panthera tigris sumatrae). BIOS 2008, 79, 7–16. [Google Scholar] [CrossRef]
- Yu, S.; Jiang, Z.; Zhu, H.; Li, C.; Zhang, E.; Zhang, J.; Harrington, C. Effects of odors on behaviors of captive Amur leopards Panthera pardus orientalis. Curr. Zool. 2009, 55, 20–27. [Google Scholar] [CrossRef]
- Galhardo, L.; Correia, J.; Oliveira, R.F. The effect of substrate availability on behavioural and physiological indicators of welfare in the African cichlid (Oreochromis mossambicus). Anim. Welf. 2008, 17, 239–254. [Google Scholar]
- Haskell, M.; Wemelsfelder, F.; Mendl, M.T.; Calvert, S.; Lawrence, A.B. The effect of substrate-enriched and substrate-impoverished housing environments on the diversity of behaviour in pigs. Behaviour 1996, 133, 741–761. [Google Scholar]
- Hirt, H.; Wechsler, B. Behavioural diversity as a measure of welfare: A study in pigs. Appl. Anim. Behav. Sci. 1994, 40, 82–83. [Google Scholar] [CrossRef]
- Wemelsfelder, F.; Haskell, M.; Mendl, M.T.; Calvert, S.; Lawrence, A.B. Diversity of behaviour during novel object tests is reduced in pigs housed in substrate-impoverished conditions. Anim. Behav. 2000, 60, 385–394. [Google Scholar] [CrossRef]
- Kistler, C.; Hegglin, D.; Würbel, H.; König, B. Feeding enrichment in an opportunistic carnivore: The red fox. Appl. Anim. Behav. Sci. 2009, 116, 260–265. [Google Scholar] [CrossRef]
- Miller, L.J.; Lauderdale, L.K.; Mellen, J.D.; Walsh, M.T.; Granger, D.A. Relationships between animal management and habitat characteristics with two potential indicators of welfare for bottlenose dolphins under professional care. PLoS ONE. under review.
- Murray, A.J.; Waran, N.K.; Young, R.J. Environmental enrichment for Australian mammals. Anim. Welf. 1998, 7, 415–425. [Google Scholar]
- Wagman, J.D. The Effects of Feeding Enrichment on Behavioral Measures of Animal Welfare in Four Bear Species. Ph.D. Thesis, Case Western Reserve University, Cleveland, OH, USA, 2015. [Google Scholar]
- Renner, M.J.; Rosenzweig, M.R. Object interactions in juvenile rats (Rattus norvegicus): Effects of different experiential histories. J. Comp. Psychol. 1986, 100, 229–236. [Google Scholar] [CrossRef]
- Spain, M.S.; Fuller, G.; Allard, S.M. Effects of habitat modifications on behavioral indicators of welfare for Madagascar giant hognose snakes (Leioheterodon madagascariensis). Anim. Behav. Cogn. 2020, 7, 70–81. [Google Scholar] [CrossRef]
- Clark, F.E.; Melfi, V.A. Environmental enrichment for a mixed-species nocturnal mammal exhibit. Zoo Biol. 2012, 31, 397–413. [Google Scholar] [CrossRef]
- Frézard, A.; Pape, G.L. Contribution to the welfare of captive wolves (Canis lupus lupus): A behavioral comparison of six wolf packs. Zoo Biol. 2003, 22, 33–44. [Google Scholar] [CrossRef]
- Hacker, C.E.; Miller, L.J.; Schulte, B.A. Examination of enrichment using space and food for African elephants (Loxodonta africana) at the San Diego Zoo Safari Park. Anim. Welf. 2018, 27, 55–65. [Google Scholar] [CrossRef]
- Kistler, C.; Hegglin, D.; Würbel, H.; König, B. Preference for structured environment in zebrafish (Danio rerio) and checker barbs (Puntius oligolepis). Appl. Anim. Behav. Sci. 2011, 135, 318–327. [Google Scholar] [CrossRef]
- Kuczaj, S.; Lacinak, T.; Fad, O.; Trone, M.; Solangi, M.; Ramos, J. Keeping environmental enrichment enriching. Int. J. Comp. Psychol. 2002, 15, 127–137. [Google Scholar]
- Alados, C.L.; Escos, J.M.; Emlen, J.M. Fractal structure of sequential behaviour patterns: An indicator of stress. Anim. Behav. 1996, 51, 437–443. [Google Scholar] [CrossRef]
- Escós, J.M.; Alados, C.L.; Emlen, J.M. Fractal structures and fractal functions as disease indicators. Oikos 1995, 74, 310–314. [Google Scholar] [CrossRef]
- MacIntosh, A.J.; Alados, C.L.; Huffman, M.A. Fractal analysis of behaviour in a wild primate: Behavioural complexity in health and disease. J. R. Soc. Interface 2011, 8, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Menchetti, L.; Righi, C.; Guelfi, G.; Enas, C.; Moscati, L.; Mancini, S.; Diverio, S. Multi-operator qualitative behavioural assessment for dogs entering the shelter. Appl. Anim. Behav. Sci. 2019, 213, 107–116. [Google Scholar] [CrossRef]
- Kühl, H.S.; Boesch, C.; Kulik, L.; Haas, F.; Arandjelovic, M.; Dieguez, P.; Bocksberger, G.; McElreath, M.B.; Agbor, A.; Angedakin, S.; et al. Human impact erodes chimpanzee behavioral diversity. Science 2019, 363, 1453–1455. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Labs Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef]
- Hall, K.; Bryant, J.; Staley, M.; Whitham, J.; Miller, L.J. Behavioral diversity as a potential positive indicator of welfare: Examining various models and their relationships to physiological welfare indicators. Anim. Welf. under review.
- Guilliams, T.G.; Edwards, L. Chronic stress and the HPA axis. Standard 2010, 9, 1–12. [Google Scholar]
- Dantzer, B.; Fletcher, Q.E.; Boonstra, R.; Sheriff, M.J. Measures of physiological stress: A transparent or opaque window into the status, management and conservation of species? Conserv. Physiol. 2014, 2, cou023. [Google Scholar] [CrossRef] [PubMed]
- Wielebnowski, N. Stress and distress: Evaluating their impact for the well-being of zoo animals. J. Am. Vet. Med. Assoc. 2003, 223, 973–977. [Google Scholar] [CrossRef] [PubMed]
- Baird, B.A. Ambassador Animal Welfare: Using Behavioral and Physiological Indicators to Assess the Well-Being of Animals Used for Education Programs in Zoos. Ph.D. Thesis, Case Western Reserve University, Cleveland, OH, USA, 2018. [Google Scholar]
- Spiezio, C.; Valsecchi, V.; Sandri, C.; Regaiolli, B. Investigating individual and social behaviour of the Northern bald ibis (Geronticus eremita): Behavioural variety and welfare. PeerJ 2018, 6, e5436. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.E.; Brereton, J.E.; Croft, D.P. Measuring welfare in captive flamingos: Activity patterns and exhibit usage in zoo-housed birds. Appl. Anim. Behav. Sci. 2018, 205, 115–125. [Google Scholar] [CrossRef]
- Pielou, E.C. An Introduction to Mathematical Ecology; Wiley-Interscience: New York, NY, USA, 1969. [Google Scholar]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Gini, C. Variabilità e Mutabilità; Tipografia di Paolo Cuppini: Bologna, Italy, 1912. [Google Scholar]
- Greenacre, M.; Hastie, T. The geometric interpretation of correspondence analysis. J. Am. Stat. Assoc. 1987, 82, 437–447. [Google Scholar] [CrossRef]
- Lebart, L.; Morineau, A.; Warwick, K.M. Multivariate Descriptive Statistical Analysis; Wiley: New York, NY, USA, 1984. [Google Scholar]
- Zipf, G.K. Human Behaviour and the Principle of Least Effort; Hafner: New York, NY, USA, 1949. [Google Scholar]
- Zipf, G.K. Human Behaivour and the Principle of Least Effort, 2nd ed.; Hafner: New York, NY, USA, 1965. [Google Scholar]
- Mandelbrot, B. Information theory and psycho1inguistics: A theory of word frequencies. In Readings in Mathematical Social Science; Lazarsfeld, P.F., Henry, N.H., Eds.; Science Research Associates: Chicago, IL, USA, 1965. [Google Scholar]
- Butler, G.C.; Yamamoto, Y.O.; Xing, H.C.; Northey, D.R.; Hughson, R.L. Heart rate variability and fractal dimension during orthostatic challenges. J. Appl. Physiol. 1993, 75, 2602–2612. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.O.; Hughson, R.L. On the fractal nature of heart rate variability in humans: Effects of data length and beta-adrenergic blockade. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1994, 266, R40–R49. [Google Scholar] [CrossRef]
- Pielou, E.C. The measurement of diversity in different types of biological collections. J. Biol. 1966, 13, 131–144. [Google Scholar] [CrossRef]
- McIntosh, R.P. An index of diversity and the relation of certain concepts to diversity. Ecology 1967, 48, 392–404. [Google Scholar] [CrossRef]
- Peet, R.K. The measurement of species diversity. Annu. Rev. Ecol. Syst. 1974, 5, 285–307. [Google Scholar] [CrossRef]
- DeJong, T.M. A comparison of three diversity indices based on their components of richness and evenness. Oikos 1975, 26, 222–227. [Google Scholar] [CrossRef]
- Cronin, K.A.; Ross, S.R. Technical contribution: A cautionary note on the use of behavioural diversity (H-Index) in animal welfare science. Anim. Welf. 2019, 28, 157–164. [Google Scholar] [CrossRef]
- Collins, C.K.; Quirke, T.; Overy, L.; Flannery, K.; O’Riordan, R. The effect of the zoo setting on the behavioural diversity of captive gentoo penguins and the implications for their educational potential. J. Zoo Aquar. Res. 2016, 3, 85–90. [Google Scholar]
- Stanton, L.A.; Sullivan, M.S.; Fazio, J.M. A standardized ethogram for the felidae: A tool for behavioral researchers. Appl. Anim. Behav. Sci. 2015, 173, 3–16. [Google Scholar] [CrossRef]
- Patil, G.P.; Taillie, C. Diversity as a concept and its measurement. J. Am. Stat. Assoc. 1982, 77, 548–567. [Google Scholar] [CrossRef]
- MacArthur, R.H. Patterns of species diversity. Biol. Rev. 1965, 40, 510–533. [Google Scholar] [CrossRef]
- Hill, M.O. Diversity and evenness: A unifying notation and its consequences. Ecology 1973, 54, 427–432. [Google Scholar] [CrossRef]
- Luebke, J.F.; Watters, J.V.; Packer, J.; Miller, L.J.; Powell, D.M. Zoo visitors’ affective responses to observing animal behaviors. Visit Stud. 2016, 19, 60–76. [Google Scholar] [CrossRef]
- Miller, L.J.; Luebke, J.F.; Matiasek, J. Viewing African and Asian elephants at accredited zoological institutions: Conservation intent and perceptions of animal welfare. Zoo Biol. 2018, 37, 466–477. [Google Scholar] [CrossRef]


{kind=link}
| Calculation Method | Number of Publications |
|---|---|
| Shannon’s Diversity Index | 28 |
| Activity Budgets | 9 |
| Behavioral Richness | 7 |
| Shannon’s Equitability | 4 |
| Behavioral Variety Index | 2 |
| Power Spectral Density | 2 |
| Behavioral Evenness | 1 |
| Gini-Simpson Index | 1 |
| Multifactorial Correspondence Analysis | 1 |
| Zipf-Mandelbrot Law | 1 |
| Species | Shannon’s Diversity Index Range | Reference |
|---|---|---|
| Ghost Bats | 3.22–3.69 | [53] |
| African Elephants | 1.43–2.11 | [59] |
| Domestic Dogs | 0.92–1.20 | [65] |
| Asiatic Lion | 0.73–1.26 | [22] |
| Black and Sun Bears | 0.69–2.09 | [25] |
| Tamandua | 0.53–2.89 | [36] |
| Leopard Cats | 0.46–0.55 | [26] |
| Leopard Gecko | 0.41–0.75 | [38] |
| Fishing Cats | 0.31–0.55 | [26] |
| Gentoo Penguins | 0.28–1.74 | [90] |
| Carnivores (34 Species) | 0.20–2.01 | [43] |
| Hognose Snake | 0.10–0.43 | [56] |
| Cheetah | 0.03–0.20 | [7] |
| Step 1. Identify species-specific behaviors | Play | Drink | Scent Mark | Hunt | |||
| Step 2. Identify contexts or components | - | - | - | Small-Sized Prey | Medium-Sized Prey | Large-Sized Prey | Birds |
| Step 3. List the natural history adaptations | - | - | - | Quick reaction time, quick response to unpredictable spatial or temporal appearance. Uses multiple sensory cues. High risk of failure. | Relies on sensory cues. Must use claws, legs, teeth, eyes, ears, balance, muscles, speed, and strength. Risk of failure or injury. | Relies on sensory cues, seasonal cues, and must use claws, legs, teeth, eyes, ears, balance, muscles, speed, and strength. Risk of failure or injury. | Stalking, noise reduction, vertical leaping ability, long leaping ability, and paw eye coordination. Uses multiple sensory cues. |
| Step 4. Determine behavioral outcomes | - | - | - | Limited window of opportunity, pouncing, improved accuracy, opportunistic, and fast responses. High energy output for small reward. | Use correct responses to cues over a long period of time. Problem solving, stalking, physically demanding, and unpredictable. High value and high motivation. | Long buildup of responses to sensory and seasonal cues, stalking, jumping, and long pre-process time (attack/kill), stashing, dragging heavy objects, high motivation, high reward, and high energy output. | Must jump and swipe, unpredictable, must be fast and have quick reaction time, and vertical pursuits. High output for small reward. |
| Step 5. Identify inputs to achieve behavioral goals | - | - | - | Offer smaller, whole prey items in hard to reach places. Only available for a designated period of time. Attached to zip line that retreats into an inaccessible area. Rabbit in polyvinyl chloride (PVC) pipe in the ground (with or without bungee). Preceded by automatic scent dispenser, environmental changes (holes dug, rabbit sounds, bushes shaking, and small pieces of pelt) for days prior. | Following scent, visual, and auditory cues for a period of days, food is located out of reach in novel locations and/or attached to bungee or other device. Offer medium-sized whole prey items following environmental events (rain, cloud cover, heat). Increase time allowed with food item, placed in a way that only some parts can be stashed if not removed by the cat properly. | Very rare following long period of visual cues (vultures), then scent cues, and finally auditory cues (other predators/scavengers, antler rubs, mating calls). Large and rare whole prey item. Very difficult to move, fixed at several points in a very difficult area to access. Only provided on occasion following a specific seasonal event (dry to wet). | Novel food item with timed delivery and quick dispersal. Small whole prey item like a bone on a vertical pulley, swipe tube, or cat operated zip line. Preceded with auditory cues days prior, and feathers closer to provision. Seasonally directed with high concentration time points after natural event (cold snap, rain event, or season change). |
| Behavior | Description |
|---|---|
| Pacing * | Walking in fixed pattern with little or no variation. Must complete 3 repetitions to qualify |
| Sit | Hind legs and rump in contact with surface |
| Stand | Upright position, all four paws on surface |
| Rest Alert * | Lying down, head up, alert, and responsive to stimulus |
| Rest Non-Alert * | Lying down, head down, and eyes mostly closed, generally immobile or sleeping |
| Flehman/Sniff | Flehman-Lips pulled back, mouth open, tongue partially extended to allow air flow. Sniff-inhaling air through nose in short repetitive manner, directed at the air, object or substrate |
| Scent Mark | Deposits odor on object(s) with intent. Includes scratch, rub, scrape, spray, and roll |
| Vocalize | Makes any vocalization |
| Self-Maintenance | Licking body or paw and passing the paw over the head, may include chewing on fur, squatting with elimination of feces or urine |
| Eat | Actively ingesting food item or water |
| Food Acquisition | Any obvious effort required to obtain food reward. Includes stalk, pounce, swat, claw |
| Play | Physically or visually engaged with an object, self (chasing paws or own tail) or conspecific in a non-serious manner |
| Social | Actively engaged in behavior with others. Same space—grooming, play, aggression, breeding, attack, pounce, stalk, and physical contact. Adjacent Space—Any obvious interaction/interest within one body length of another cat in an adjacent space. Allogrooming—grooming or being groomed by a conspecific |
| Locomotion | Forward motion, non-stereotypic |
| Out of View * | Out of view of the observer |
| Other * | Any other behavior not described above |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miller, L.J.; Vicino, G.A.; Sheftel, J.; Lauderdale, L.K. Behavioral Diversity as a Potential Indicator of Positive Animal Welfare. Animals 2020, 10, 1211. https://doi.org/10.3390/ani10071211
Miller LJ, Vicino GA, Sheftel J, Lauderdale LK. Behavioral Diversity as a Potential Indicator of Positive Animal Welfare. Animals. 2020; 10(7):1211. https://doi.org/10.3390/ani10071211
Chicago/Turabian StyleMiller, Lance J., Greg A. Vicino, Jessica Sheftel, and Lisa K. Lauderdale. 2020. "Behavioral Diversity as a Potential Indicator of Positive Animal Welfare" Animals 10, no. 7: 1211. https://doi.org/10.3390/ani10071211
APA StyleMiller, L. J., Vicino, G. A., Sheftel, J., & Lauderdale, L. K. (2020). Behavioral Diversity as a Potential Indicator of Positive Animal Welfare. Animals, 10(7), 1211. https://doi.org/10.3390/ani10071211