Urinary Reference Values and First Insight into the Urinary Proteome of Captive Giraffes

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Urine Sampling
2.2. Urinalysis
2.3. One-D-Electrophoresis
2.4. Protein Identification by Mass Spectrometry
2.5. Statistical Analysis
3. Results
3.1. Selection of Reference Giraffes and Outlier Identification
3.2. Urinalysis
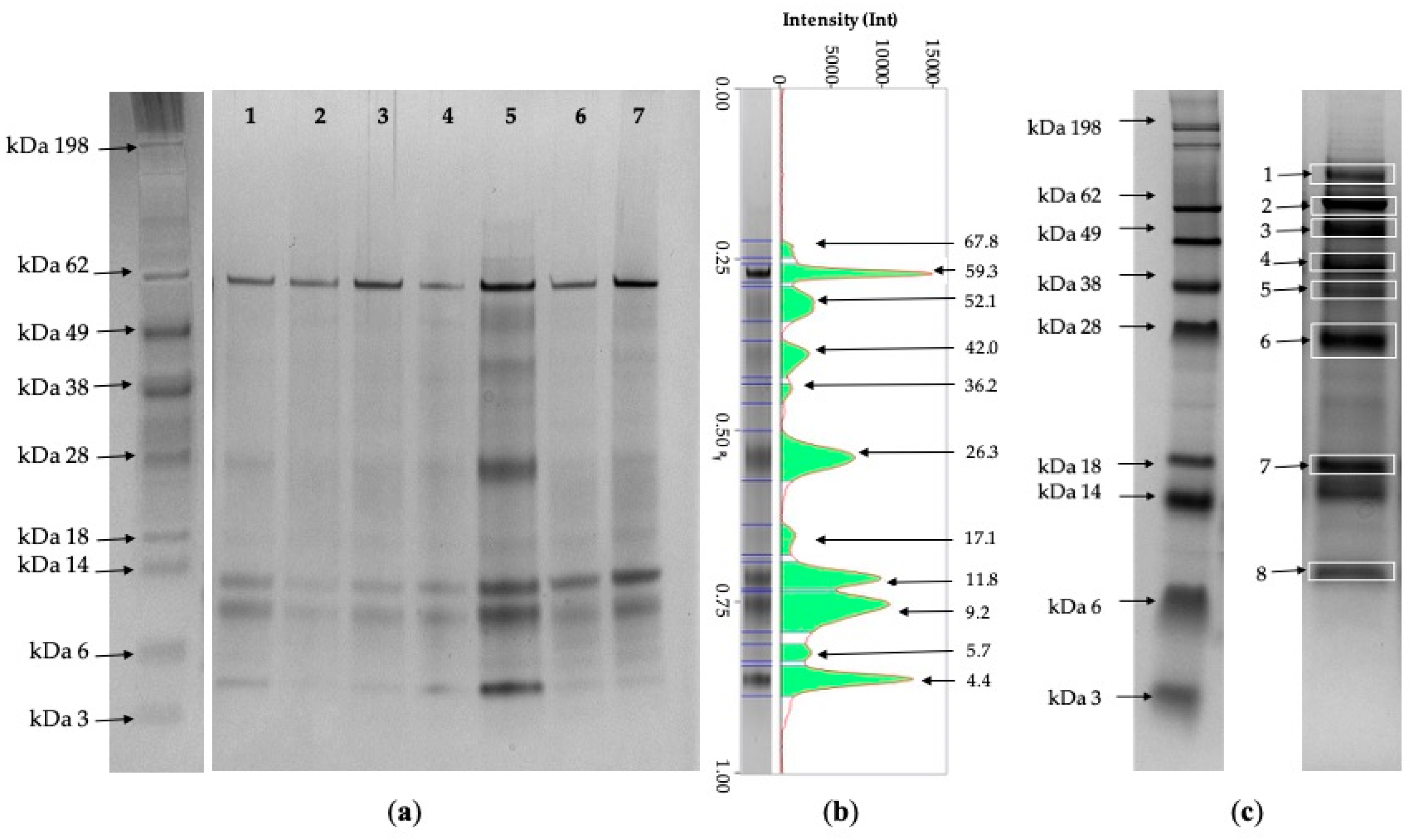
3.3. One-D-Electrophoresis
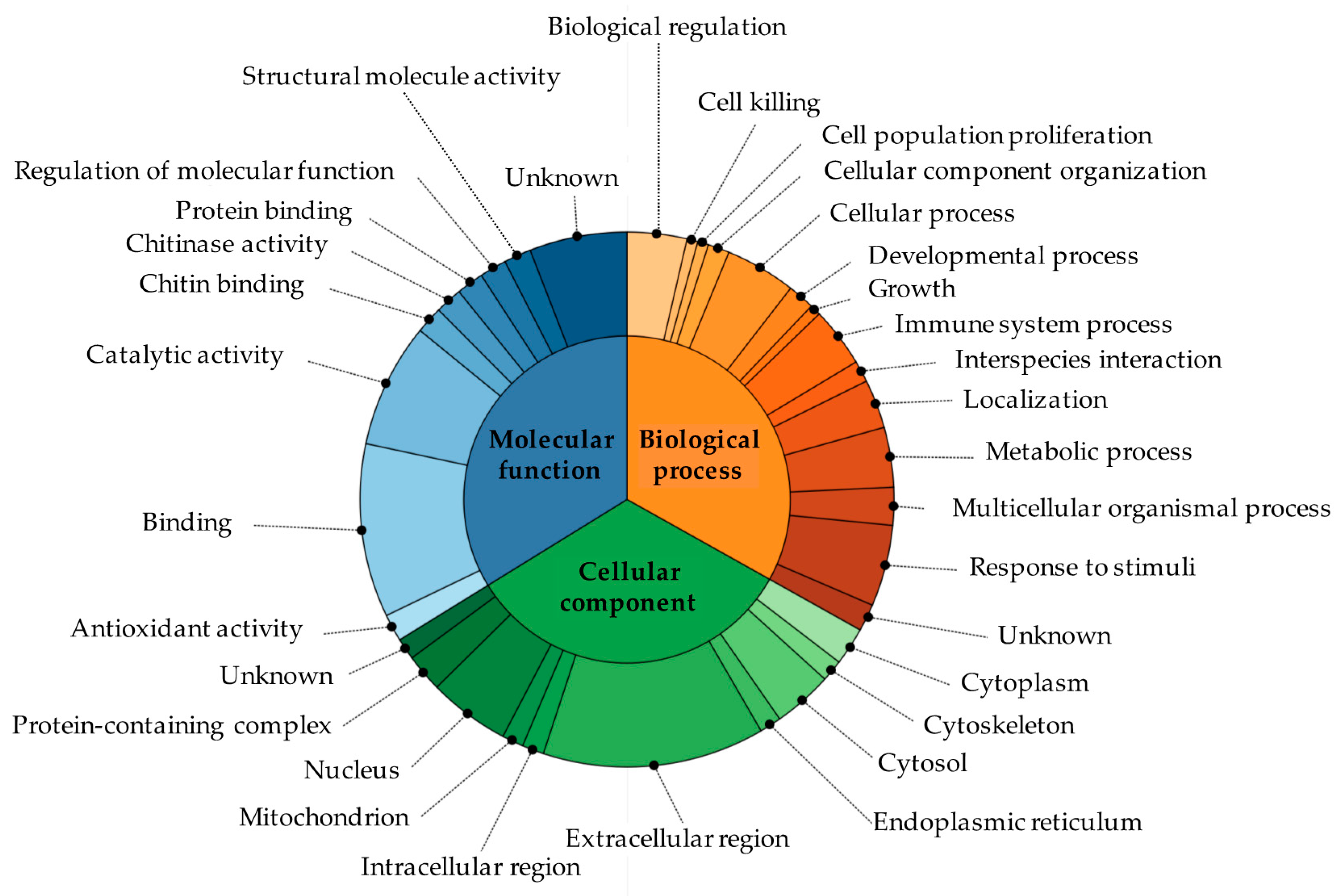
3.4. Protein Identification Using Mass Spectrometry
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Piech, T.L.; Wycislo, K.L. Importance of Urinalysis. Vet. Clin. Small. Anim. 2019, 49, 233–245. [Google Scholar] [CrossRef]
- Callens, A.J.; Bartges, J.W. Urinalysis. Vet. Clin. Small. Anim. 2015, 45, 621–637. [Google Scholar] [CrossRef]
- Friedrich, K.R.; Harr, K.E.; Freeman, K.P.; Szladovits, B.; Walton, R.M.; Barnhart, K.F.; Blanco-Chavez, J. ASVCP reference interval guidelines: Determination of de novo reference intervals in veterinary species and other related topics. Vet. Clin. Pathol. 2012, 41, 441–453. [Google Scholar] [CrossRef]
- Zhu, C.; Fasoli, S.; Isani, G.; Laghi, L. First insights into the urinary metabolome of captive giraffes by Proton Nuclear Magnetic Resonance Spectroscopy. Metabolites 2020, 10, 157. [Google Scholar] [CrossRef]
- Sullivan, K.; Freeman, S.; van Heugten, E.; Ange-van Heugten, K.; Wolfe, B.; Poore, M.H. Impact of two types of complete pelleted, wild ungulate feeds and two pelleted feed to hay ratios on the development of urolithogenic compounds in meat goats as a model for giraffes. J. Anim. Physiol. Anim. Nutr. 2013, 97, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, K.; van Heugten, E.; Ange-van Heugten, K.; Poore, M.H.; Dierenfeld, E.S.; Wolfe, B. Analysis of Nutrient Concentrations in the Diet, Serum, and Urine of Giraffe from Surveyed North American Zoological Institutions. Zoo Biol. 2010, 29, 457–469. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, B.A.; Sladky, K.K.; Loomis, M.R. Obstructive urolithiasis in a reticulated giraffe (Giraffa camelopardalis reticulata). Vet. Rec. 2000, 146, 260–261. [Google Scholar] [CrossRef] [PubMed]
- Fasoli, S.; Ferlizza, E.; Andreani, G.; Sandri, C.; Dondi, F.; Isani, G. Non-invasive sampling method for urinalysis and urine protein profile in captive giraffes. J. Vet. Diagn. Investig. 2020. accepted. [Google Scholar]
- Hokamp, J.A.; Leidy, S.A.; Gaynanova, I.; Cianciolo, R.E.; Nabity, M.B. Correlation of electrophoretic urine protein banding patterns with severity of renal damage in dogs with proteinuric chronic kidney disease. Vet. Clin. Pathol. 2018, 47, 425–434. [Google Scholar] [CrossRef]
- De Loor, J.; Daminet, S.; Smets, P.; Maddens, B.; Meyer, E. Urinary biomarkers for Acute Kidney Injury in dogs. J. Vet. Intern. Med. 2013, 27, 998–1010. [Google Scholar] [CrossRef]
- Ferlizza, E.; Isani, G.; Dondi, F.; Andreani, G.; Vasylyeva, K.; Bellei, E.; Almeida, A.M.; Matzapetakis, M. Urinary proteome and metabolome in dogs (Canis lupus familiaris): The effect of chronic kidney disease. J. Proteom. 2020, 222, 103795. [Google Scholar] [CrossRef] [PubMed]
- Ferlizza, E.; Campos, A.; Neagu, A.; Cuoghi, A.; Bellei, E.; Monari, E.; Dondi, F.; Almeida, A.M.; Isani, G. The effect of chronic kidney disease on the urine proteome in the domestic cat (Felis catus). Vet. J. 2015, 204, 73–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, Y.M.; Zhang, S.; Zhao, J.Y.; Liu, C.Y. Adipokine zinc-alpha-2- glycoprotein as a novel urinary biomarker presents earlier than microalbuminuria in diabetic nephropathy. J. Int. Med. Res. 2016, 44, 278–286. [Google Scholar] [CrossRef] [Green Version]
- Vaidya, V.S.; Ferguson, M.A.; Bonventre, J.V. Biomarkers of Acute Kidney Injury. Annu. Rev. Pharmacol. Toxicol. 2008, 48, 463–493. [Google Scholar] [CrossRef] [Green Version]
- Smets, P.M.Y.; Meyer, E.; Maddens, B.E.J.; Duchateau, L.; Daminet, S. Urinary markers in healthy young and aged dogs and dogs with chronic kidney disease. J. Vet. Intern. Med. 2010, 24, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Ferlizza, E.; Dondi, F.; Andreani, G.; Bucci, D.; Archer, J.; Isani, G. Validation of an electrophoretic method to detect albuminuria in cats. J. Feline Med. Surg. 2017, 19, 860–868. [Google Scholar] [CrossRef]
- Ferlizza, E.; Fasoli, S.; Cavallini, D.; Bolcato, M.; Andreani, G.; Isani, G. Preliminary study on urine chemistry and protein profile in cows and heifers. Pak. Vet. J. 2020. [Google Scholar] [CrossRef]
- Isani, G.; Ferlizza, E.; Nally, J.E. Proteomic research in urine and other fluids. In Proteomics in Domestic Animals: From Farm to Systems Biology; Almeida, A.M., Eckersall, D., Miller, I., Eds.; Springer: Cham, Switzerland, 2018; pp. 121–147. [Google Scholar]
- McLean, L.; Hurst, J.L.; Gaskell, C.J.; Lewis, J.C.; Beynon, R.J. Characterization of cauxin in the urine of domestic and big cats. J. Chem. Ecol. 2007, 33, 1997–2009. [Google Scholar] [CrossRef]
- Alhaider, A.A.; Bayoumy, N.; Argo, E.; Gader, A.G.M.A.; Stead, D.A. Survey of the camel urinary proteome by shotgun proteomics using a multiple database search strategy. Proteomics 2012, 12, 3403–3406. [Google Scholar] [CrossRef]
- Neely, B.A.; Prager, K.C.; Bland, A.M.; Fontaine, C.; Gulland, F.M.; Janech, M.G. Proteomic Analysis of Urine from California Sea Lions (Zalophus californianus): A Resource for Urinary Biomarker Discovery. J. Proteome Res. 2018, 17, 3281–3291. [Google Scholar] [CrossRef]
- Gage, L.J. Giraffe Husbandry and Welfare. In Fowler’s Zoo and Wild Animal Medicine Current Therapy, 1st ed.; Miller, E.R., Lamberski, N., Calle, P., Eds.; Elsevier: St. Louis, MO, USA, 2019; Volume 9, pp. 619–622. [Google Scholar]
- Morgan, K.N.; Tromborg, C.T. Sources of stress in captivity. Appl. Anim. Behav. Sci. 2007, 102, 262–302. [Google Scholar] [CrossRef]
- Isani, G.; Ferlizza, E.; Cuoghi, A.; Bellei, E.; Monari, E.; Bianchin Butina, B.; Castagnetti, C. Identification of the most abundant proteins in equine amniotic fluid by a proteomic approach. Anim. Reprod. Sci. 2016, 174, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Bellei, E.; Rustichelli, C.; Bergamini, S.; Monari, E.; Baraldi, C.; Lo Castro, F.; Tomasi, A.; Ferrari, A. Proteomic serum profile in menstrual-related and post menopause migraine. J. Pharm. Biomed. Anal. 2020, 184, 113165. [Google Scholar] [CrossRef] [PubMed]
- Muller, Z. Population structure of giraffes is affected by management in the Great Rift Valley, Kenya. PLoS ONE 2018, 13, e0189678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrie, A.; Watson, P. Additional techniques. In Statistics for Veterinary and Animal Science, 3rd ed.; Petrie, A., Watson, P., Eds.; John Wiley & Sons: Chichester, UK, 2013; pp. 200–229. [Google Scholar]
- Geffré, A.; Concordet, D.; Braun, J.P.; Trumel, C. Reference Value Advisor: A new freeware set of macroinstructions to calculate reference intervals with Microsoft Excel. Vet. Clin. Pathol. 2011, 40, 107–112. [Google Scholar] [CrossRef]
- Olea-Popelka, F.; Rosen, L.E. A Practical Guide for Statistics in Wildlife Studies. In Fowler’s Zoo and Wild Animal Medicine Current Therapy, 1st ed.; Miller, E.R., Lamberski, N., Calle, P., Eds.; Elsevier: St. Louis, MO, USA, 2019; Volume 9, pp. 21–27. [Google Scholar]
- Herman, N.; Bourgès-Abella, N.; Braun, J.P.; Ancel, C.; Schelcher, F.; Trumel, C. Urinalysis and determination of the urine protein-to-creatinine ratio reference interval in healthy cows. J. Vet. Intern. Med. 2019, 33, 999–1008. [Google Scholar] [CrossRef]
- Jones, M.; Miesner, M.D.; Baird, A.N.; Pugh, D.G. Diseases of the urinary system. In Sheep and Goats Medicine, 2nd ed.; Pugh, D.G., Baird, A.N., Eds.; Elsevier Saunders: Maryland Heights, MO, USA, 2012; pp. 325–360. [Google Scholar]
- Grauer, G.F. Proteinuria: Measurement and Interpretation. Top. Companion. Anim. Med. 2011, 26, 121–127. [Google Scholar] [CrossRef]
- Sink, C.A.; Weinstein, N.M. Specimen procurement. In Practical Veterinary Urinalysis; Sink, C.A., Weinstein, N.M., Eds.; John Wiley & Sons: Chichester, UK, 2012; pp. 9–18. [Google Scholar]
- Wiedner, E.; Alleman, A.R.; Isaza, R. Urinalysis in Asian elephants (Elephas maximus). J. Zoo Wildl. Med. 2009, 40, 659–666. [Google Scholar] [CrossRef]
- Wolfe, B.A. Bovidae (Except Sheep and Goats) and Antilocapridae. In Fowler’s Zoo and Wild Animal Medicine; Miller, R.E., Fowler, M.E., Eds.; Elsevier: St. Louis, MO, USA, 2015; Volume 8, pp. 626–645. [Google Scholar]
- Haffey, M.B.; Pairan, R.D.; Reinhart, P.R.; Stoops, M.A. Urinalysis in three species of captive rhinoceros (Rhinoceros unicornis, Dicerorhinus sumatrensis, and Diceros bicornis). J. Zoo Wildl. Med. 2008, 39, 349–357. [Google Scholar] [CrossRef]
- Chew, D.J.; DiBartola, S.P.; Schneck, P. Clinical Evaluation of the Urinary Tract. In Canine and Feline Nephrology and Urology, 2nd ed.; Chew, D.J., DiBartola, S.P., Schneck, P., Eds.; Elsevier Saunders: Saint Louis, MO, USA, 2011; pp. 32–62. [Google Scholar]
- Braun, J.P.; Lefebvre, H.P. Kidney Function and Damage. In Clinical Biochemistry of Domestic Animals; Kaneko, J.J., Harvey, J.W., Bruss, M.L., Eds.; Academic Press: San Diego, CA, USA, 2008; pp. 485–528. [Google Scholar]
- Gautier, J.C.; Gury, T.; Guffroy, M.; Khan-Malek, R.; Hoffman, D.; Pettit, S.; Harpur, E. Normal Ranges and Variability of Novel Urinary Renal Biomarkers in Sprague-Dawley Rats: Comparison of Constitutive Values between Males and Females and across Assay Platforms. Toxicol. Pathol. 2014, 42, 1092–1104. [Google Scholar] [CrossRef]
- Thomas, N.J.; Shields, B.M.; Besser, R.E.J.; Jones, A.G.; Rawlingson, A.; Goodchild, E.; Leighton, C.; Bowman, P.; Shepherd, M.; Knight, B.A.; et al. The impact of gender on urine C-peptide creatinine ratio interpretation. Ann. Clin. Biochem. 2012, 49, 363–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, S.; Sugiura, M.; Tsutsumi, S.; Yamada, H. Sex differences in the excretion levels of traditional and novel urinary biomarkers of nephrotoxicity in rats. J. Toxicol. Sci. 2017, 42, 615–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dagg, A.I. Anatomy. In Giraffe: Biology, Behaviour and Conservation; Dagg, A.I., Ed.; Cambridge University Press: New York, NY, USA, 2014; pp. 100–116. [Google Scholar]
- Phillips, J.K.; McBride, C.A.; Hale, S.A.; Solomon, R.J.; Badger, G.J.; Bernstein, I.M. Examination of pre-pregnancy and pregnancy urinary protein levels in healthy nulliparous women. Reprod. Sci. 2017, 24, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Molina, L.; Salvetat, N.; Ameur, R.B.; Peres, S.; Sommerer, N.; Jarraya, F.; Ayadi, H.; Molina, F.; Granier, C. Analysis of the variability of human normal urine by 2D-GE reveals a “public” and a “private” proteome. J. Proteom. 2011, 75, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Bathla, S.; Rawat, P.; Baithalu, R.; Yadav, M.L.; Naru, J.; Tiwari, A.; Kumar, S.; Balhara, A.K.; Singh, S.; Chaudhary, S.; et al. Profiling of urinary proteins in Karan Fries cows reveals more than 1550 proteins. J. Proteom. 2015, 127, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Brandt, L.E.; Ehrhart, E.J.; Scherman, H.; Olver, C.S.; Bohn, A.A.; Prenni, J.E. Characterization of the canine urinary proteome. Vet. Clin. Pathol. 2014, 43, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Miller, I.; Preßlmayer-Hartler, A.; Wait, R.; Hummel, K.; Sensi, C.; Eberini, I.; Razzazi-Fazeli, E.; Gianazza, E. In between—Proteomics of dog biological fluids. J. Proteom. 2014, 106, 30–45. [Google Scholar] [CrossRef]
- Devuyst, O.; Olinger, E.; Rampoldi, L. Uromodulin: From physiology to rare and complex kidney disorders. Nat. Rev. Nephrol. 2017, 13, 525–544. [Google Scholar] [CrossRef]
- Jones, M.E.B.; Gasper, D.J.; Mitchell (née Lane), E. Bovidae, Antilocapridae, Giraffidae, Tragulidae, Hippopotamidae. In Pathology of Wildlife and Zoo Animals; Terio, K.A., McAloose, D., Leger, J.S., Eds.; Academic Press: London, UK, 2018; pp. 117–147. [Google Scholar]
- Tabata, E.; Kashimura, A.; Kikuchi, A.; Masuda, H.; Miyahara, R.; Hiruma, Y.; Wakita, S.; Ohno, M.; Sakaguchi, M.; Sugahara, Y.; et al. Chitin digestibility is dependent on feeding behaviors, which determine acidic chitinase mRNA levels in mammalian and poultry stomachs. Sci. Rep. 2018, 8, 1461. [Google Scholar] [CrossRef]
- Mack, I.; Hector, A.; Ballbach, M.; Kohlhäufl, J.; Fuchs, K.J.; Weber, A.; Mall, M.A.; Hartl, D. The role of chitin, chitinases, and chitinase-like proteins in pediatric lung diseases. Mol. Cell. Pediatr. 2015, 2. [Google Scholar] [CrossRef] [Green Version]
- Ohno, M.; Kimura, M.; Miyazaki, H.; Okawa, K.; Onuki, R.; Nemoto, C.; Tabata, E.; Wakita, S.; Kashimura, A.; Sakaguchi, M.; et al. Acidic Mammalian Chitinase Is a Proteases-Resistant Glycosidase in Mouse Digestive System. Sci. Rep. 2016, 6, 37756. [Google Scholar] [CrossRef] [PubMed]
- Vannella, K.M.; Ramalingam, T.R.; Hart, K.M.; de Queiroz Prado, R.; Sciurba, J.; Barron, L.; Borthwick, L.A.; Smith, A.D.; Mentink-Kane, M.; White, S.; et al. Acidic chitinase primes the protective immune response to gastrointestinal nematodes. Nat. Immunol. 2016, 17, 538–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddens, B.; Ghesquière, B.; Vanholder, R.; Demon, D.; Vanmassenhove, J.; Gevaert, K.; Meyer, E. Chitinase-like proteins are candidate biomarkers for sepsis-induced acute kidney injury. Mol. Cell. Proteom. 2012, 11, M111.013094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maack, T. Renal handling of low molecular weight proteins. Am. J. Med. 1975, 58, 57–64. [Google Scholar] [CrossRef]
- Van Harten, R.M.; van Woudenbergh, E.; van Dijk, A.; Haagsman, H.P. Cathelicidins: Immunomodulatory Antimicrobials. Vaccines 2018, 6, 63. [Google Scholar] [CrossRef] [Green Version]
- Shannan, B.; Seifert, M.; Leskov, K.; Willis, J.; Boothman, D.; Tilgen, W.; Reichrath, J. Challenge and promise: Roles for clusterin in pathogenesis, progression and therapy of cancer. Cell Death Differ. 2006, 13, 12–19. [Google Scholar] [CrossRef]
- Lepanto, M.S.; Rosa, L.; Paesano, R.; Valenti, P.; Cutone, A. Lactoferrin in Aseptic and Septic Inflammation. Molecules 2019, 24, 1323. [Google Scholar] [CrossRef] [Green Version]
- Hassan, I.M.; Waheed, A.; Yadav, S.; Singh, T.P.; Ahmad, F. Zinc A2-Glycoprotein: A Multidisciplinary Protein. Mol. Cancer. Res. 2008, 6, 892–906. [Google Scholar] [CrossRef] [Green Version]
- Clerc, F.; Reiding, K.R.; Jansen, B.C.; Kammeijer, G.S.M.; Bondt, A.; Wuhrer, M. Human plasma protein N-glycosylation. Glycoconj. J. 2016, 33, 309–343. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Schwesinger, C. The ubiquitin-proteasome system in kidney physiology and disease. Nat. Rev. Nephrol. 2019, 15, 393–411. [Google Scholar] [CrossRef]
- Ronzaud, C.; Staub, O. Ubiquitylation and Control of Renal Na Balance and Blood Pressure. Physiology 2014, 29, 16–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, Y.; Horikawa, S.; Katayama, Y.; Sasaki, S. Water channel aquaporin-2 directly binds to actin. Biochem. Biophys. Res. Commun. 2004, 322, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, S.; Noda, Y. Aquaporin-2 protein dynamics within the cell. Curr. Opin. Nephrol. Hypertens. 2007, 16, 348–352. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Moeller, H.B.; Stevens, D.A.; Sanchez-Hodge, R.; Childers, G.; Kortenoeven, M.L.A.; Cheng, L.; Rosenbaek, L.L.; Rubel, C.; Patterson, C.; et al. CHIP Regulates Aquaporin-2 Quality Control and Body Water Homeostasis. J. Am. Soc. Nephrol. 2018, 29, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Warda, M.; Prince, A.; Kim, H.K.; Khafaga, N.; Scholkamy, T.; Linhardt, R.J.; Jin, H. Proteomics of old world camelid (Camelus dromedarius): Better understanding the interplay between homeostasis and desert environment. J. Adv. Res. 2014, 5, 219–242. [Google Scholar] [CrossRef] [Green Version]
- Dagg, A.I. Individual behaviours. In Giraffe Biology, Behaviour and Conservation; Dagg, A.I., Ed.; Cambridge University Press: New York, NY, USA, 2014; pp. 60–86. [Google Scholar]
- Damkjær, M.; Wang, T.; Brøndum, E.; Østergaard, K.H.; Baandrup, U.; Hørlyck, A.; Hasenkam, J.M.; Smerup, M.; Funder, J.; Marcussen, N.; et al. The giraffe kidney tolerates high arterial blood pressure by high renal interstitial pressure and low glomerular filtration rate. Acta Physiol. 2015, 214, 497–510. [Google Scholar] [CrossRef]
- Nawata, M.C.; Pannabecker, T.L. Mammalian urine concentration: A review of renal medullary architecture and membrane transporters. J. Comp. Physiol. B 2018, 188, 899–918. [Google Scholar] [CrossRef]
- Dugina, V.; Zwaenepoel, I.; Gabbiani, G.; Clément, S.; Chaponnier, C. β- and γ-cytoplasmic actins display distinct distribution and functional diversity. J. Cell Sci. 2009, 122, 2980–2988. [Google Scholar] [CrossRef] [Green Version]
- Parker, F.; Baboolal, T.G.; Peckham, M. Actin Mutations and Their Role in Disease. Int. J. Mol. Sci. 2020, 21, 3371. [Google Scholar] [CrossRef]
- Saeidi, B.; Koralkar, R.; Griffin, R.L.; Halloran, B.; Ambalavanan, N.; Askenazi, D. Impact of gestational age, sex, and postnatal age on urine biomarkers in premature neonates. Pediatr. Nephrol. 2015, 30, 2037–2044. [Google Scholar] [CrossRef]
- Guo, Z.; Zhang, Y.; Zou, L.; Wang, D.; Shao, C.; Wang, Y.; Sun, W.; Zhang, L. A Proteomic Analysis of Individual and Gender Variations in Normal Human Urine and Cerebrospinal Fluid Using iTRAQ Quantification. PLoS ONE 2015, 10, e0133270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muniyan, S.; Chaturvedi, N.K.; Dwyer, J.G.; LaGrange, C.A.; Chaney, W.G.; Lin, M.-F. Human Prostatic Acid Phosphatase: Structure, Function and Regulation. Int. J. Mol. Sci. 2013, 14, 10438–10464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyo, J.; Hwang, S.I.; Oh, J.; Lee, S.J.; Kang, S.C.; Kim, J.S.; Lim, J. Characterization of a bovine pregnancy-associated protein using two-dimensional gel electrophoresis, N-terminal sequencing and mass spectrometry. Proteomics 2003, 3, 2420–2427. [Google Scholar] [CrossRef] [PubMed]
- Rawat, P.; Bathla, S.; Baithalu, R.; Yadav, M.L.; Kumar, S.; Ali, S.A.; Tiwari, A.; Lotfan, M.; Naru, J.; Jena, M.; et al. Identification of potential protein biomarkers for early detection of pregnancy in cow urine using 2D DIGE and label free quantitation. Clin. Proteom. 2016, 13, 15. [Google Scholar] [CrossRef] [Green Version]
- Balhara, A.K.; Gupta, M.; Singh, S.; Mohanty, A.K.; Singh, I. Early Pregnancy Diagnosis in Bovines: Current Status and Future Directions. Sci. World J. 2013, 10. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Variable | N | Mean | Median | SD | LL (90% CI Range) | UL (90% CI Range) |
|---|---|---|---|---|---|---|
| Bil | 34 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Ery (RBC/µL) | 34 | 3 | 0 | 9 | 0 | 50 |
| Glu (mmol/L) | 34 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Ket (mmol/L) | 34 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Leu (WBC/µL) | 34 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| Nit | 34 | Neg | Neg | - | Neg | Trace |
| pH | 34 | 9.0 | 9.0 | 0.5 | 8.0 | 9.0 |
| Pro (mg/dL) | 34 | 30.0 | 30.0 | 32.0 | 0.0 | 100.0 |
| UBG (µmol/L) | 34 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| uCr (mg/dL) | 41 | 145.23 | 154.62 | 93.56 | 39.59 (26.31–61.56) | 357.95 (302.62–425.80) |
| UPC | 40 | 0.11 | 0.11 | 0.03 | 0.07 (0.07–0.08) | 0.16 (0.15–0.17) |
| USG | 34 | 1.028 | 1.030 | 0.012 | 1.006 (1001–1013) | 1049 (1044–1053) |
| uTP (mg/dL) | 40 | 15.78 | 17.58 | 8.78 | 4.54 (3.03–7.09) | 35.31 (30.23–40.18) |
| N | Database | Accession | Full Protein Name | Species | Score | MM | Mass | Peptides | Pep (sig) | Sequence | Seq (sig) | SC (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | SwissProt | UROM_BOVIN | Uromodulin | Bos taurus | 602 | 77 | 72 | 55 | 35 | 18 | 14 | 20% |
| SwissProt | TRFL_BOVIN | Lactotransferrin | Bos taurus | 16 | 77 | 80 | 8 | 2 | 8 | 2 | 9% | |
| 2 | SwissProt | ALBU_BOVIN | Serum albumin | Bos taurus | 1529 | 64 | 71 | 180 | 104 | 46 | 30 | 50% |
| SwissProt | ALBU_SHEEP | Serum albumin | Ovis aries | 1450 | 64 | 71 | 154 | 92 | 42 | 27 | 43% | |
| 3 | SwissProt | CHIA_BOVIN | Acidic mammalian chitinase | Bos taurus | 196 | 50 | 52 | 16 | 8 | 10 | 5 | 15% |
| SwissProt | A1BG_BOVIN | Alpha-1B-glycoprotein | Bos taurus | 63 | 50 | 54 | 11 | 6 | 5 | 5 | 6% | |
| 4 | SwissProt | CLUS_BOVIN | Clusterin | Bos taurus | 389 | 42 | 51 | 32 | 21 | 15 | 9 | 21% |
| SwissProt | ZA2G_BOVIN | Zinc-alpha-2-glycoprotein | Bos taurus | 231 | 42 | 34 | 19 | 12 | 9 | 8 | 21% | |
| SwissProt | PEPA_BOVIN | Pepsin A | Bos taurus | 166 | 42 | 40 | 31 | 17 | 8 | 6 | 9% | |
| SwissProt | ACTB_BOVIN | Actin, cytoplasmic 1 | Bos taurus | 50 | 42 | 42 | 11 | 3 | 10 | 3 | 29% | |
| 5 | SwissProt | HPT_CAPIB | Haptoglobin | Capra ibex | 92 | 34 | 45 | 12 | 6 | 10 | 5 | 18% |
| SwissProt | DNAS1_PIG | Deoxyribonuclease-1 | Sus scrofa | 85 | 34 | 31 | 11 | 4 | 5 | 3 | 19% | |
| SwissProt | APOD_BOVIN | Apolipoprotein D | Bos taurus | 64 | 34 | 21 | 12 | 5 | 5 | 3 | 20% | |
| 6 | SwissProt | CTHL1_SHEEP | Cathelicidin-1 | Ovis aries | 48 | 25 | 18 | 5 | 2 | 4 | 2 | 23% |
| 7 | SwissProt | LYSC2_BOVIN | Lysozyme C-2 | Bos taurus | 375 | 18 | 16 | 26 | 16 | 7 | 4 | 51% |
| SwissProt | CTHL1_SHEEP | Cathelicidin-1 | Ovis aries | 58 | 18 | 18 | 9 | 5 | 7 | 5 | 38% | |
| 8 | SwissProt | UBIQ_CAMDR | Ubiquitin | Camelus dromedarius | 52 | 10 | 8.5 | 9 | 3 | 7 | 2 | 80% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fasoli, S.; Andreani, G.; Dondi, F.; Ferlizza, E.; Bellei, E.; Isani, G. Urinary Reference Values and First Insight into the Urinary Proteome of Captive Giraffes. Animals 2020, 10, 1696. https://doi.org/10.3390/ani10091696
Fasoli S, Andreani G, Dondi F, Ferlizza E, Bellei E, Isani G. Urinary Reference Values and First Insight into the Urinary Proteome of Captive Giraffes. Animals. 2020; 10(9):1696. https://doi.org/10.3390/ani10091696
Chicago/Turabian StyleFasoli, Sabrina, Giulia Andreani, Francesco Dondi, Enea Ferlizza, Elisa Bellei, and Gloria Isani. 2020. "Urinary Reference Values and First Insight into the Urinary Proteome of Captive Giraffes" Animals 10, no. 9: 1696. https://doi.org/10.3390/ani10091696
APA StyleFasoli, S., Andreani, G., Dondi, F., Ferlizza, E., Bellei, E., & Isani, G. (2020). Urinary Reference Values and First Insight into the Urinary Proteome of Captive Giraffes. Animals, 10(9), 1696. https://doi.org/10.3390/ani10091696