Entomological Surveillance and Cantharidin Concentrations in Mylabris variabilis and Epicauta rufidorsum Blister Beetles in Slovenia

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Entomological Surveillance
2.2. Cantharidin Detection
2.2.1. Standards and Chemicals
2.2.2. Analytical Procedure
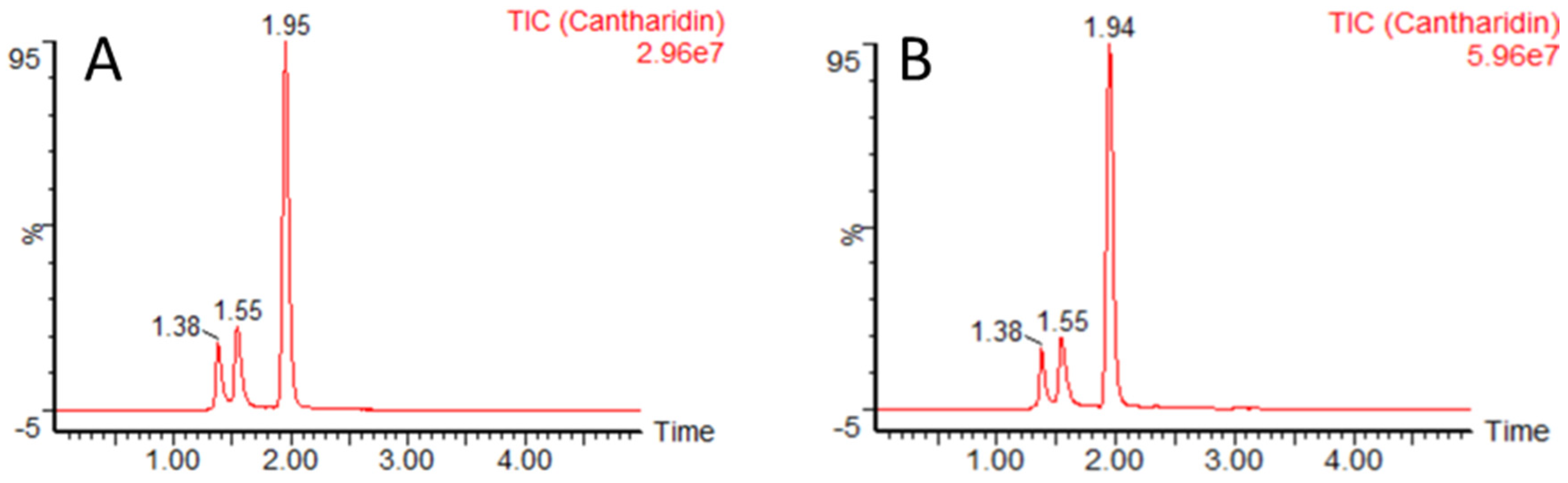
2.2.3. LC-MS/MS Analysis
2.2.4. Statistical Analysis
3. Results
3.1. Entomological Observations
3.2. Total Cantharidin Concentrations
3.3. Cantharidin Concentrations in M. variabilis and E. rufidorsum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pan, Z.; Ren, G.D.; Wang, X.P.; Bologna, M.A. Revision of the genus Pseudabris Fairmaire (Coleoptera, Meloidae), an endemic to the Tibetan Plateau, with biogeographical comments. Syst. Entomol. 2013, 38, 134–150. [Google Scholar] [CrossRef]
- McCormick, J.P.; Carrel, J.E. Cantharidin biosynthesis and function in meloid beetles. In Pheromone Biochemistry; Prestwich, G.D., Bloomquist, H.F., Eds.; Harcourt, Brace, & Jovanovich: Orlando, FL, USA, 1987; pp. 307–350. [Google Scholar]
- Ghoneim, K. Cantharidin toxicosis to animal and human in the world: A review. Standard Res. J. Toxicol. Environ. Health Sci. 2013, 1, 1–16. [Google Scholar]
- Frenzel, M.; Dettner, K. Quantification of cantharidin in canthariphilous Ceratopogonidae (Diptera), Anthomyiidae (Diptera) and cantharidin-producing Oedemeridae (Coleoptera). J. Chem. Ecol. 1994, 20, 1795–1812. [Google Scholar] [CrossRef] [PubMed]
- Day, R.M.; Harbord, M.; Forbes, A.; Segal, A.W. Cantharidin blisters: A technique for investigating leukocyte trafficking and cytokine production at sites of inflammation in humans. J. Immunol. Methods 2001, 257, 213–220. [Google Scholar] [CrossRef]
- Mullen, G.R.; Durden, L.A. Medical and Veterinary Entomology, 2nd ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: Boston, MA, USA, 2009. [Google Scholar]
- Dettner, K. Inter-and Intraspecific Transfer of Toxic Insect Compound Cantharidin. In Vertical Food Web Interactions; Springer: Berlin/Heidelberg, Germany, 1997; pp. 115–145. [Google Scholar]
- Gisondi, S.; Gasperi, T.; Roma, E.; Tomai, P.; Gentili, A.; Vignoli, L.; Bologna, M.A.; Mancini, E. Cantharidin content in two Mediterranean species of blister beetles, Lydus trimaculatus and Mylabris variabilis (Coleoptera: Meloidae). Entomol. Sci. 2019, 22, 258–263. [Google Scholar] [CrossRef]
- Ray, A.C.; Post, L.O.; Hurst, J.M.; Edwards, W.C.; Reagor, J.C. Evaluation of an analytical method for the diagnosis of cantharidin toxicosis due to ingestion of blister beetles (Epicauta lemniscata) by horses and sheep. Am. J. Vet. Res. 1980, 41, 932–933. [Google Scholar]
- Gayle, L.G.; Reagor, J.C.; Ray, A.; Bailey, E.M. Cantharidin poisoning in cattle. J. Am. Vet. Med. Assoc. 1981, 179, 263. [Google Scholar]
- Penrith, M.L.; Naude, T.W. Mortality in chickens associated with blister beetle consumption. J. S. Afr. Vet. Assoc. 1996, 67, 97–99. [Google Scholar]
- Helman, R.G.; Edwards, W.C. Clinical features of blister beetle poisoning in equids: 70 cases (1983–1996). J. Am. Vet. Med. Assoc. 1997, 211, 1018–1021. [Google Scholar]
- Tagwireyi, D.; Ball, D.E.; Loga, P.J.; Moyo, S. Cantharidin poisoning due to “Blister beetle” ingestion. Toxicon 2000, 38, 1865–1869. [Google Scholar] [CrossRef]
- Sánchez-Barbudo, I.S.; Camarero, P.R.; García-Montijano, M.; Mateo, R. Possible cantharidin poisoning of a great bustard (Otis tarda). Toxicon 2012, 59, 100–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, K.M.; Streeter, R.N.; De Souza, P.; Genova, S.G.; Morgan, S.E. Cantharidin Toxicosis in 2 alpacas. Can. Vet. J. 2013, 54, 456. [Google Scholar] [PubMed]
- Schoeb, T.R.; Panciera, R.J. Blister beetle poisoning in horses. J. Am. Vet. Med. Assoc. 1978, 173, 75–77. [Google Scholar] [PubMed]
- Schmitz, D.G. Cantharidin toxicosis in horses. J. Vet. Int. Med. 1989, 3, 208–215. [Google Scholar] [CrossRef]
- Blodgett, S.L.; Carrel, J.E.; Higgins, R.A. Cantharidin contamination of alfalfa hay. J. Med. Entomol. 1992, 29, 700–703. [Google Scholar] [CrossRef]
- Zhu, Y.-C.; Charlton, R.E.; Higgins, R.A. Factors influencing quantity of cantharidin transferred to alfalfa from Epicauta occidentalis Werner (Coleoptera: Meloidae). J. Econ. Entomol. 1997, 90, 1665–1671. [Google Scholar] [CrossRef]
- Fenyvesi, I.; Gounden, V.; Lutchman, N.; Snyman, T. Case report: Identification of possible cantharidin contamination in a South African traditional remedy. Toxicol. Lett. 2010, 196, S294. [Google Scholar] [CrossRef]
- Bonness, K.; Aragon, I.V.; Rutland, B.; Ofori-Acquah, S.; Dean, N.M.; Honkanen, R.E. Cantharidin-induced mitotic arrest is associated with the formation of aberrant mitotic spindles and lagging chromosomes resulting, in part, from the suppression of PP2Aalpha. Mol. Cancer Ther. 2006, 5, 2727–2736. [Google Scholar] [CrossRef] [Green Version]
- Database of Invertebrate Pictures PMSL. Epicauta rufidorsum (Goeze, 1777). 2020. Available online: http://www1.pms-lj.si/animalia/load.php?species=3747 (accessed on 16 November 2020).
- Global Biodiversity Information Facility. Available online: www.gbif.org (accessed on 16 November 2020).
- GBIF. Mylabris variabilis (Pallas, 1781). Sources of the Data: Application Inpn Espèces: Observations Naturalistes, Participatives et Opportunistes, Fondées sur des Photographies; cen_paca_2017_12_18; Données Entomologiques, a. Horellou 1988–2018; Données Naturalistes de Maxime Esnault.; Données onf Faune-Flore-Fonge; Données Photographiques Validées de la Galerie du Forum “le Monde des Insectes”; Explor’nature 2017, Inventaire Biologique de la Commune de Barcelonnette; Geographically Tagged Insdc Sequences; Inaturalist Research-Grade Observations; International Barcode of Life Project (Ibol); jeu de Données Convention Fegve-Spn-2012–2016; Naturalia_2017_12_18; Naturgucker; Observation.org, Nature Data from Around the World; Observations Faune et Flore du Parc National Des écrins; Parc Naturel Régional du Verdon 2017_12_18; Proserpine_2017_12_18; Prospections des Invertébrés Terrestres—La Planète Revisitée Corse 2019; Sorties de Terrain du Forum “le Monde des Insectes”; Ssnatv_2017_12_18; Swiss National Coleoptera Databank. Available online: https://www.gbif.org/species/5751064 (accessed on 16 November 2020).
- GBIF. Epicauta rufidorsum (Goeze, 1777). Sources of the Data: Biologiezentrum Linz, iNaturalist Research-Grade Observations, Frost Entomological Museum, Lund Museum of Zoology—Insect Collections (MZLU), Données Photographiques Validées de la Galerie du Forum “Le Monde des Insectes”, Données Naturalistes de Hervé BOUYON. Available online: https://www.gbif.org/species/4457450 (accessed on 16 November 2020).
- Altman, J. Observational sampling methods for insect behavioral ecology. Fla. Entomol. 1984, 67, 50–66. [Google Scholar] [CrossRef]
- Abtahi, S.M.; Nikbakhtzadeh, M.R.; Vatandoost, H.; Mehdinia, A.; Foroshani, A.R.; Shayeghi, M. Quantitative characterization of cantharidin in the false blister beetle, Oedemera podagrariae ventralis, of the southern slopes of Mount Elborz, Iran. J. Insect Sci. 2012, 12, 152. [Google Scholar] [CrossRef]
- Nikbakhtzadeh, M.R.; Vahedi, M.; Vatandoost, H.; Mehdinia, A. Origin, transfer and distribution of cantharidin-related compounds in the blister beetle Hycleus scabiosae. J. Venom. Anim. Toxins Incl. Trop. Dis. 2012, 18, 88–96. [Google Scholar] [CrossRef] [Green Version]
- GraphPad Prism; Version 6.04 for Windows; GraphPad Software Inc.: San Diego, CA, USA; Available online: www.graphpad.com (accessed on 30 September 2020).
- Wikipedia. Mylabris variabilis. Available online: https://en.wikipedia.org/wiki/Mylabris_variabilis#Distribution (accessed on 16 November 2020).
- Pan, Z.; Bologna, M.A. Taxonomy, bionomics and faunistics of the nominate subgenus of Mylabris Fabricius, 1775, with the description of five new species (Coleoptera: Meloidae: Mylabrini). Zootaxa 2014, 3806, 1–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallas, P.S. Reise Durch Verschiedene Provinzen des Rußischen Reichs Theil 1 (Nr.46 Meloe erythrocephalus); Gedruckt bey der Kayserlichen Academie der Wissenschaften: Saint Petersburg, Russia, 1771; p. 466. [Google Scholar]
- Kaszab, Z. Die paläarktichen und orientalischen Arten der Meloiden-Gattung Epicauta Redtb. Acta Zool. Acad. Sci. Hung. 1952, 3, 573–599. [Google Scholar]
- Özbek, H.; Szaloki, D. A contribution to the knowledge of the Meloidae (Coleoptera) fauna of Turkey along with new records. Tr. J. Zool. 1998, 22, 23–40. [Google Scholar]
- Siberian Zoological Museum of the Institute of Animal Systematics and Ecology. Available online: http://szmn.sbras.ru/Coleop/Meloidae.htm (accessed on 16 November 2020).
- Zandigiacomo, P.; Zlatich, M. Epicauta rufidorsum: Un Meloide di interesse naturalistico dannoso alla patata in provincia di Trieste. Notiziario ERSA 2006, 3–4, 38–39. [Google Scholar]
- Horváth, Z.; Hatvani, A.; Škorić, D. Damage in sunflower fields and in hybrid multiplication caused by species of the Coleoptera (Anthiciadae) family. Helia 2014, 26, 101–108. [Google Scholar] [CrossRef]
- Mebs, D.; Pogoda, W.; Schneider, M.; Kauert, G. Cantharidin and demethylcantharidin (palasonin) content of blister beetles (Coleoptera: Meloidae) from southern Africa. Toxicon 2009, 53, 466–468. [Google Scholar] [CrossRef]
- Nikbakhtzadeh, M.R.; Tirgari, S. Cantharidin component of Iranian blister beetles (Col: Meloidae) and their differences between Iranian and exotic species. Iran. J. Publ. Health 2002, 31, 113–117. [Google Scholar]
- Blodgett, S.L.; James, E.C.; Higgins, R.A. Cantharidin content of blister beetles (Coleoptera: Meloidae) collected from Kansas alfalfa and implications for inducing cantharidiasis. Environ. Entomol. 1991, 20, 776–780. [Google Scholar] [CrossRef]
- Percino-Daniel, N.; Buckley, D.; García-París, M. Pharmacological properties of blister beetles (Coleoptera: Meloidae) promoted their integration into the cultural heritage of native rural Spain as inferred by vernacular names diversity, traditions, and mitochondrial DNA. J. Ethnopharmacol. 2013, 147, 570–583. [Google Scholar] [CrossRef] [Green Version]
- Bravo, C.; Bautista, L.M.; García-París, M.; Blanco, G.; Alonso, J.C. Males of a strongly polygynous species consume more poisonous food than females. PLoS ONE 2014, 9, e111057. [Google Scholar] [CrossRef] [PubMed]
- Whitman, D.W.; Andrés, M.F.; Martínez-Díaz, R.A.; Ibáñez-Escribano, A.; Olmeda, A.S.; González-Coloma, A. Antiparasitic properties of cantharidin and the blister beetle Berberomeloe majalis (Coleoptera: Meloidae). Toxins 2019, 11, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capinera, J.L.; Gardner, D.R.; Stermitz, F.R. Cantharidin levels in blister beetles (Coleoptera: Meloidae) associated with alfalfa in Colorado. J. Econ. Entomol. 1985, 78, 1052–1055. [Google Scholar] [CrossRef]
- Carrel, J.E.; McCairel, M.H.; Slagle, A.J.; Doom, J.P.; Brill, J.; McCormick, J.P. Cantharidin production in a blister beetle. Experientia 1993, 49, 171–174. [Google Scholar] [CrossRef]
- Sierra, J.R.; Woggon, W.D.; Schmid, H. Transfer of cantharidin (1) during copulation from the adult male to the female Lytta vesicatoria (‘Spanish flies’). Experientia 1976, 32, 142–144. [Google Scholar] [CrossRef]
- Nikbakhtzadeh, M.R. Transfer and Distribution of Cantharidin within Selected Members of Blister Beetles (Coleoptera: Meloidae) and Its Probable Importance in Sexual Behaviour. Doctoral Dissertation, The University of Bayreuth, Bayreuth, Germany, 2004. [Google Scholar]
- Whitman, D.W. Allelochemical interactions among plants, herbivores, and their predators. In Novel Aspects of Insect-Plant Interactions; Barbosa, P., Letrourneau, D., Eds.; John Wiley: New York, NY, USA, 1988; pp. 11–64. [Google Scholar]
- Alcock, J.; Hadley, N.F. Tests of alternative hypotheses for assortative mating by size: A comparison of two meloid beetles (Coleoptera: Meloidae). J. Kans. Entomol. Soc. 1987, 41–50. [Google Scholar]
- Spinoni, J.; Naumann, G.; Vogt, J.V.; Barbosa, P. The biggest drought events in Europe from 1950 to 2012. J. Hydrol. Reg. Stud. 2015, 3, 509–524. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
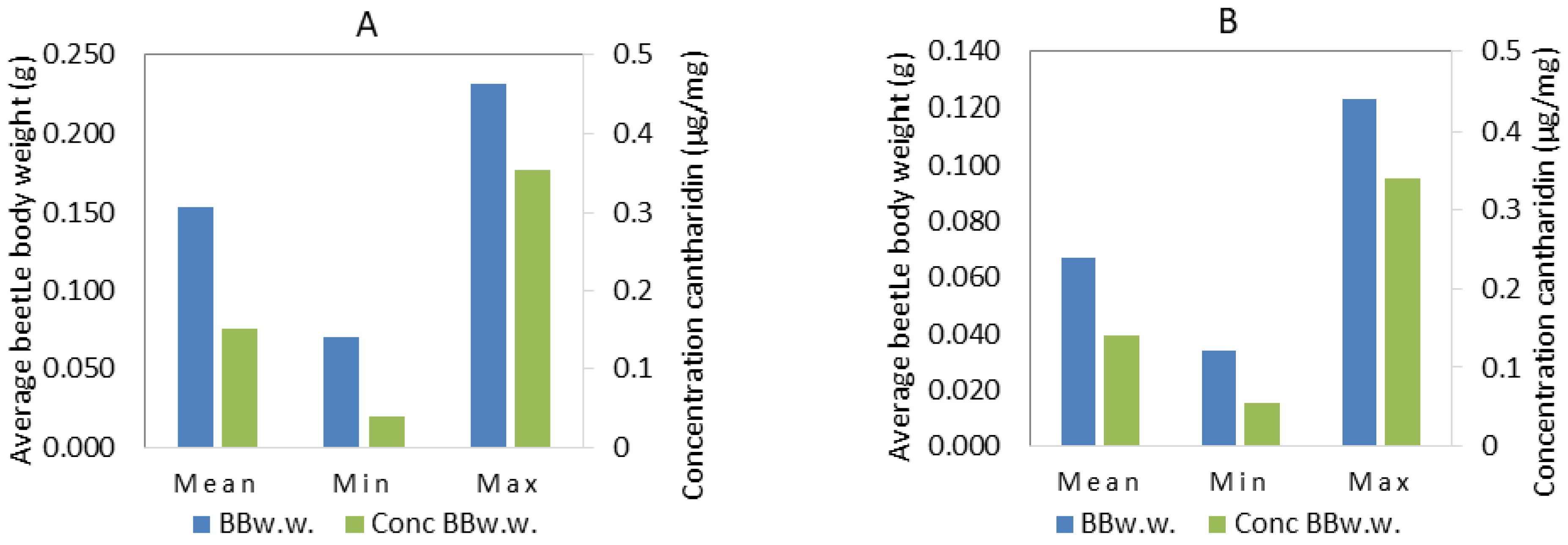
| Descriptive Statistics | mCanth. (µg) | BBd.w. (g) | Conc. d.w. (µg/mg) | BBw.w. (g) | Conc. w.w. (µg/mg) |
|---|---|---|---|---|---|
| Mean | 22.2 | 0.064 | 0.377 | 0.166 | 0.145 |
| Min | 6.5 | 0.026 | 0.100 | 0.070 | 0.038 |
| Max | 51.8 | 0.123 | 1.08 | 0.334 | 0.354 |
| Median | 21.4 | 0.065 | 0.322 | 0.165 | 0.128 |
| Species | Sex | Body w.w. (g) | Body d.w. (g) | Cantharidin per Beetle (mg) | Cantharidin Concentration d.w. (%) |
|---|---|---|---|---|---|
| M. variabilis | M (n = 7) | 0.172 ± 0.042 | 0.067 ± 0.015 | 0.022 ± 0.012 | 0.032 ± 0.014 |
| F (n = 10) | 0.141 ± 0.046 | 0.049 ± 0.016 | 0.022 ± 0.009 | 0.049 ± 0.026 | |
| Total | 0.154 ± 0.046 | 0.056 ± 0.017 | 0.022 ± 0.010 | 0.042 ± 0.023 | |
| 1p value M × F | 0.281 | 0.066 | 0.944 | 0.239 | |
| E. rufidorsum | M (n = 33) | 0.173 ± 0.054 | 0.067 ± 0.019 | 0.022 ± 0.006 | 0.036 ± 0.015 |
| F (n = 3) | 0.164 ± 0.060 | 0.070 ± 0.027 | 0.025 ± 0.009 | 0.037 ± 0.001 | |
| Total | 0.172 ± 0.054 | 0.067 ± 0.020 | 0.022 ± 0.006 | 0.036 ± 0.014 | |
| 1p value M × F | 0.628 | 0.990 | 0.405 | 0.238 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakovac-Strajn, B.; Brozić, D.; Tavčar-Kalcher, G.; Babič, J.; Trilar, T.; Vengust, M. Entomological Surveillance and Cantharidin Concentrations in Mylabris variabilis and Epicauta rufidorsum Blister Beetles in Slovenia. Animals 2021, 11, 220. https://doi.org/10.3390/ani11010220
Jakovac-Strajn B, Brozić D, Tavčar-Kalcher G, Babič J, Trilar T, Vengust M. Entomological Surveillance and Cantharidin Concentrations in Mylabris variabilis and Epicauta rufidorsum Blister Beetles in Slovenia. Animals. 2021; 11(1):220. https://doi.org/10.3390/ani11010220
Chicago/Turabian StyleJakovac-Strajn, Breda, Diana Brozić, Gabrijela Tavčar-Kalcher, Janja Babič, Tomi Trilar, and Modest Vengust. 2021. "Entomological Surveillance and Cantharidin Concentrations in Mylabris variabilis and Epicauta rufidorsum Blister Beetles in Slovenia" Animals 11, no. 1: 220. https://doi.org/10.3390/ani11010220
APA StyleJakovac-Strajn, B., Brozić, D., Tavčar-Kalcher, G., Babič, J., Trilar, T., & Vengust, M. (2021). Entomological Surveillance and Cantharidin Concentrations in Mylabris variabilis and Epicauta rufidorsum Blister Beetles in Slovenia. Animals, 11(1), 220. https://doi.org/10.3390/ani11010220