Is Intestinal Bacterial Diversity Enhanced by Trans-Species Spread in the Mixed-Species Flock of Hooded Crane (Grus monacha) and Bean Goose (Anser fabalis) Wintering in the Lower and Middle Yangtze River Floodplain?



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Site Selection and Fecal Sample Collection
2.3. Fecal DNA Extraction
2.4. Bird Species Determination
2.5. Illumina MiSeq Sequencing
2.6. Processing of Sequenced Data
2.7. Identification of Potential Pathogenic Bacteria
2.8. Biostatistical Analyses
3. Results
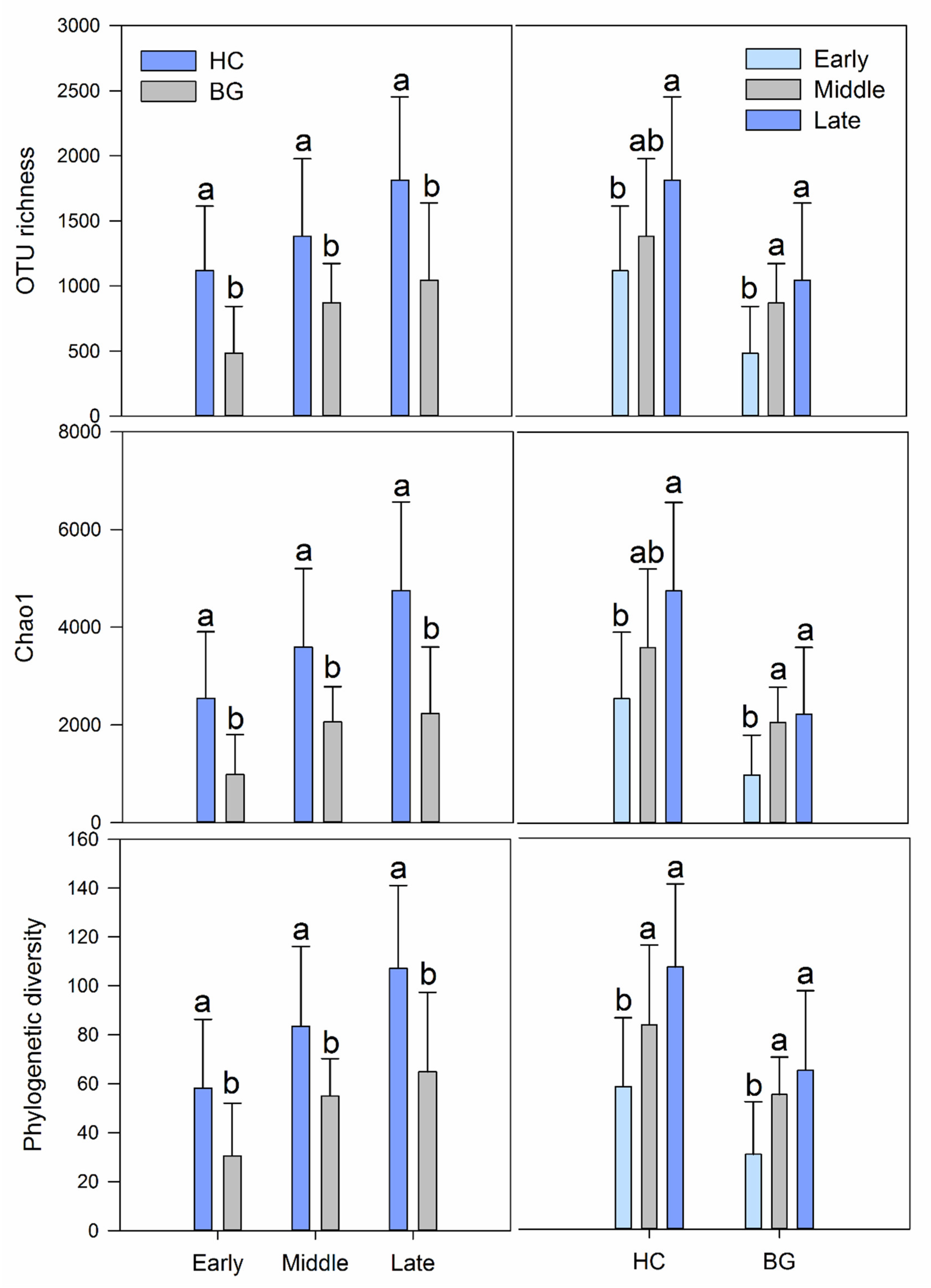
3.1. Differences in Intestinal Bacterial Diversity between Hooded Crane and Bean Goose
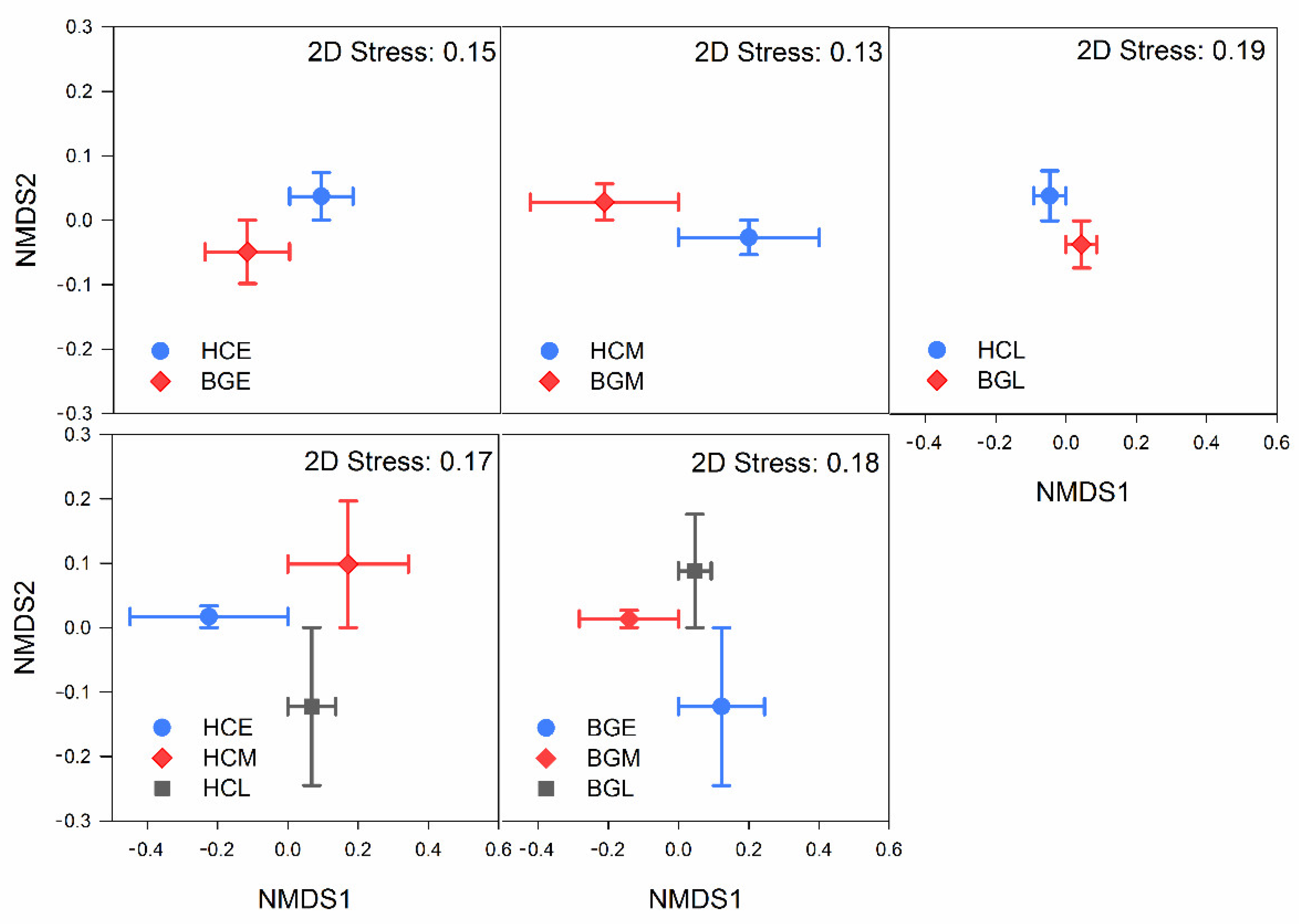
3.2. Intestinal Bacterial Diversity and Its Diversity Patterns in Hooded Cranes and Bean Geese
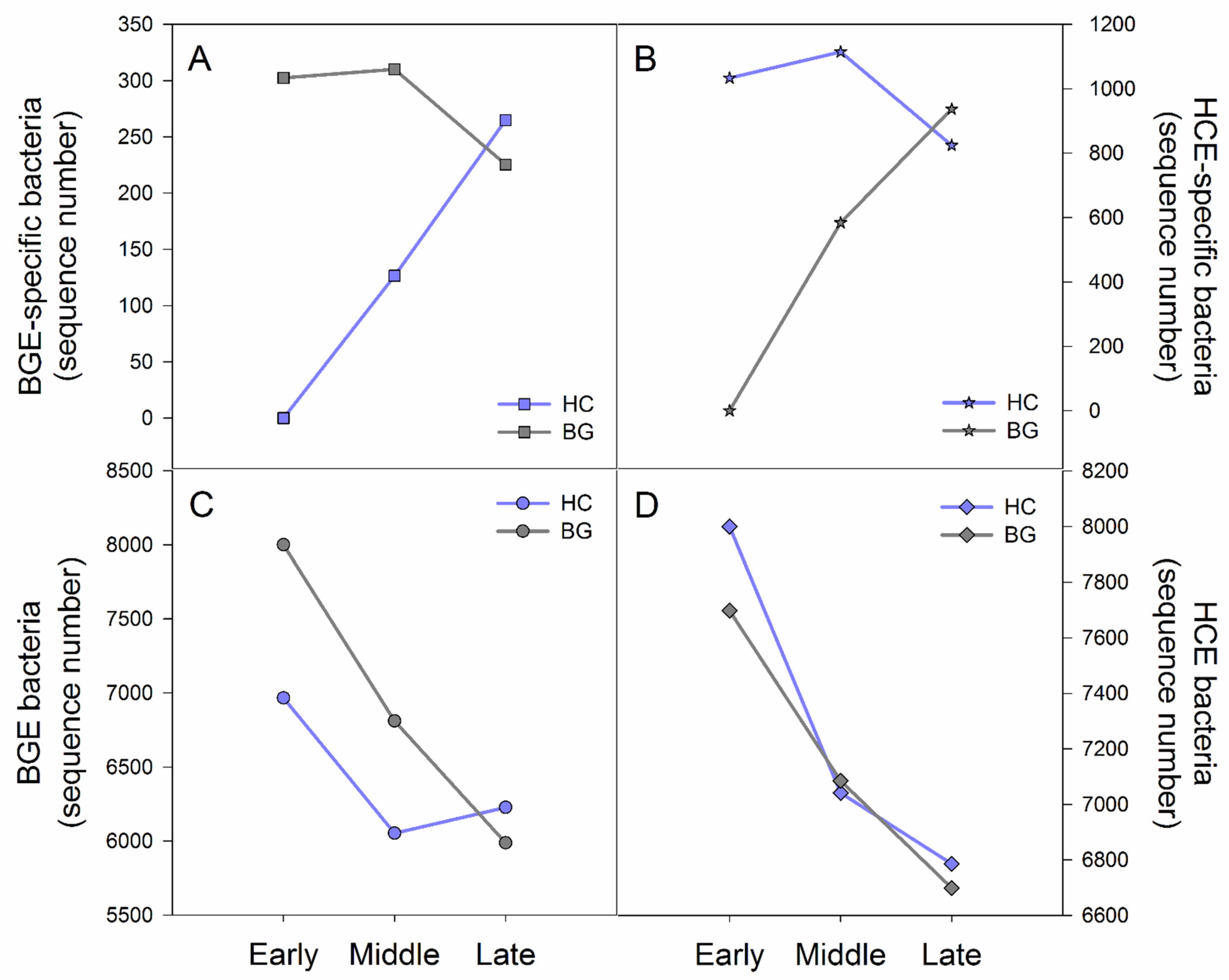
3.3. Variation Characteristics of Early Period Bacteria in Hooded Crane and Bean Goose during the Wintering Period
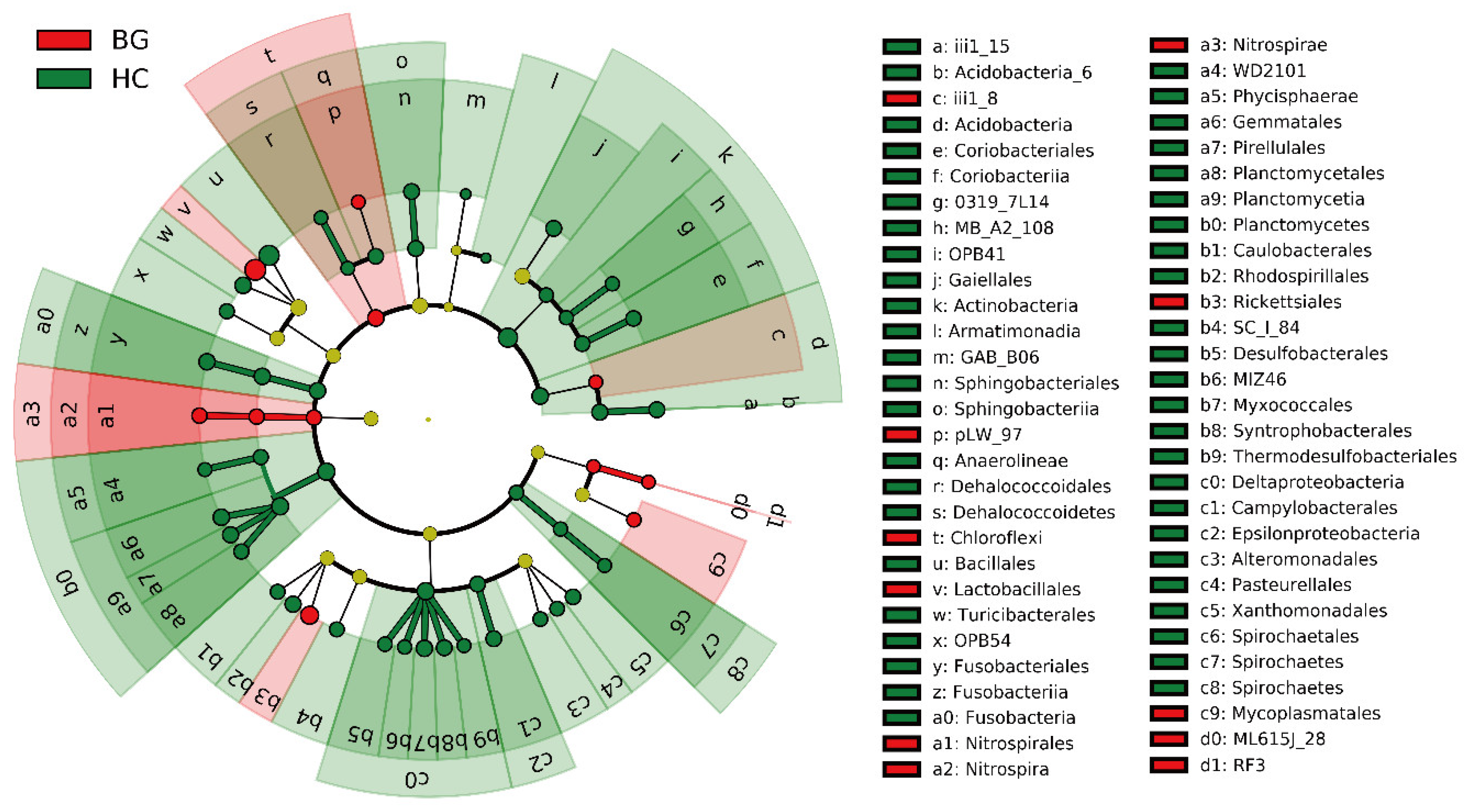
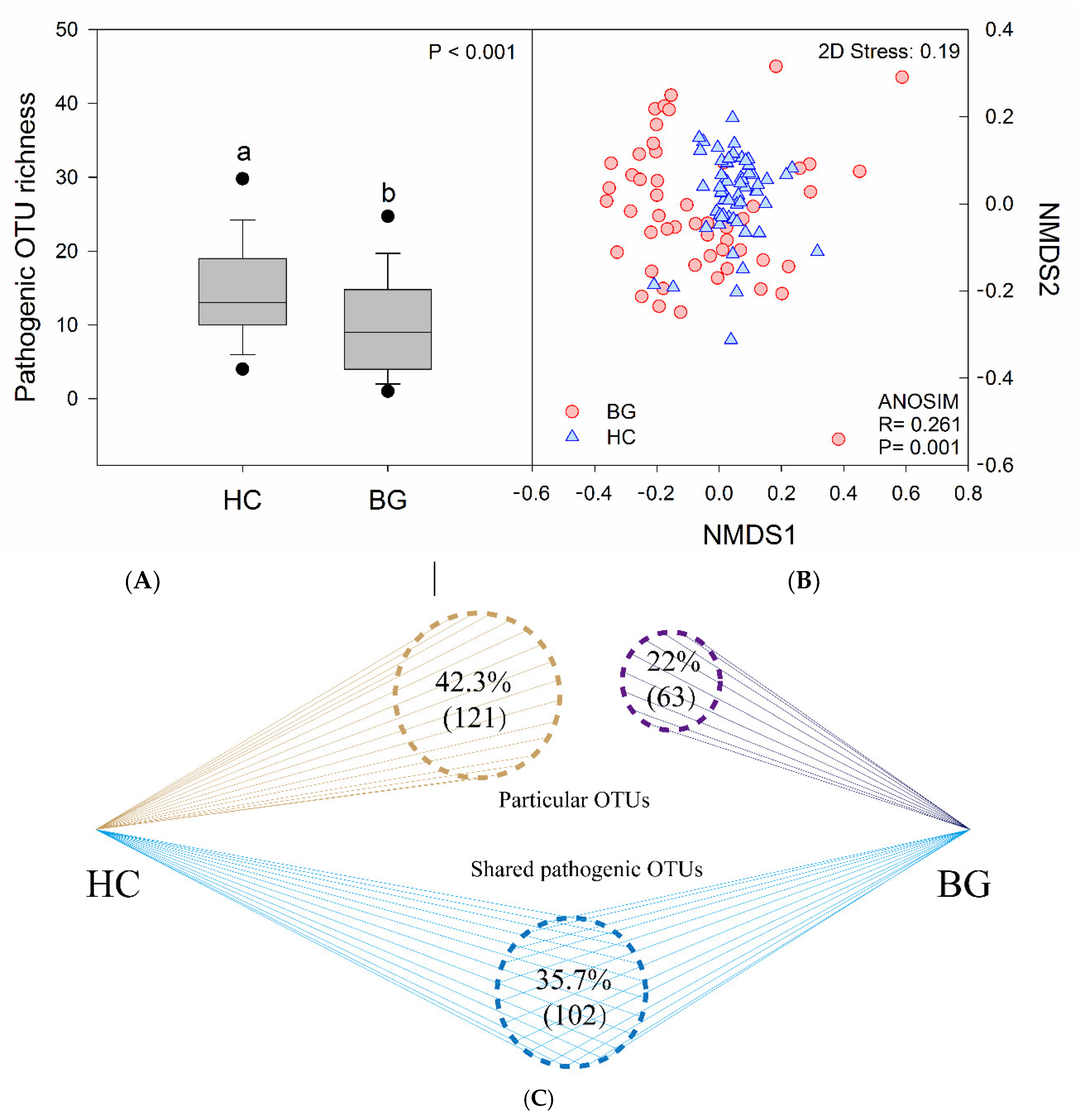
3.4. Intestinal Potential Pathogens in Hooded Cranes and Bean Geese
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lewis, W.B.; Moore, F.R.; Wang, S. Characterization of the gut microbiota of migratory passerines during stopover along the northern coast of the Gulf of Mexico. J. Avian Biol. 2016, 47, 1–10. [Google Scholar] [CrossRef]
- Xiang, X.J.; Zhang, F.L.; Fu, R.; Yan, S.F.; Zhou, L.Z. Significant differences in bacterial and potentially pathogenic communities between sympatric hooded crane and greater white-fronted goose. Front. Microbiol. 2019, 10, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waite, D.W.; Taylor, M.W. Characterizing the avian gut microbiota: Membership, driving influences, and potential function. Front. Microbiol. 2014, 5, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vo, A.T.E. Determinants and Implications of Variation in the Avian Gut Microbiota. Ph.D. Thesis, University of California, Berkeley, CA, USA, 2014. [Google Scholar]
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Treuren, W.V.; Knight, R.; Bell, J.T.; et al. Human genetics shape the gut microbiome. Cell 2014, 159, 789–799. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Rodriguez, M.; Lucas, F.S.; Heeb, P.; Soler, J.J. Differences in intestinal microbiota between avian brood parasites and their hosts. Biol. J. Linn. Soc. 2009, 96, 406–414. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Lozupone, C.A.; Hamady, M.; Knight, R.; Gordon, J.I. Worlds within worlds: Evolution of the vertebrate gut microbiota. Nat. Rev. Microbiol. 2008, 6, 776–788. [Google Scholar] [CrossRef] [Green Version]
- Ley, R.E.; Hamady, M.; Lozupone, C.; Turnbaugh, P.; Ramey, R.R.; Bircher, J.S.; Schlege, M.L.; Tucker, T.A.; Schrenzel, M.D.; Knight, R.; et al. Evolution of mammals and their gut microbes. Science 2008, 320, 1647–1651. [Google Scholar] [CrossRef] [Green Version]
- Bolnick, D.I.; Snowberg, L.K.; Hirsch, P.E.; Lauber, C.L.; Org, E.; Parks, B.; Lusis, A.J.; Knight, R.; Caporaso, J.G.; Svanbäck, R.; et al. Individual diet has sex-dependent effects on vertebrate gut microbiota. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef]
- Inamine, H.; Ellner, S.P.; Newell, P.D.; Luo, Y.; Buchon, N.; Douglas, A.E. Spatiotemporally heterogeneous population dynamics of gut bacteria inferred from fecal time series data. mBio 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.Y.; Chen, P.C.; Weng, F.C.H.; Shaw, G.T.W.; Wang, D. Habitat and indigenous gut microbes contribute to the plasticity of gut microbiome in oriental river prawn during rapid environmental change. PLoS ONE 2017, 12, e0181427. [Google Scholar] [CrossRef]
- Seedorf, H.; Griffin, N.W.; Ridaura, V.K.; Reyes, A.; Cheng, J.; Rey, F.E.; Smith, M.I.; Simon, G.M.; Scheffrahn, R.H.; Woebken, D.; et al. Bacteria from diverse habitats colonize and compete in the mouse gut. Cell 2014, 159, 253–266. [Google Scholar] [CrossRef] [Green Version]
- Groussin, M.; Mazel, F.; Sanders, J.G.; Smillie, C.S.; Lavergne, S.; Thuiller, W.; Alm, E.J. Unraveling the processes shaping mammalian gut microbiomes over evolutionary time. Nat. Commun. 2017, 8, 14319. [Google Scholar] [CrossRef] [Green Version]
- Garud, N.R.; Good, B.H.; Hallatschek, O.; Pollard, K.S. Evolutionary dynamics of bacteria in the gut microbiome within and across hosts. PLoS Biol. 2019, 17, e3000102. [Google Scholar] [CrossRef] [Green Version]
- Liddicoat, C.; Sydnor, H.; Cando-Dumancela, C.; Dresken, R.; Liu, J.; Gellie, N.J.C.; Mills, J.G.; Young, J.M.; Weyrich, L.S.; Hutchinson, M.R.; et al. Naturally-diverse airborne environmental microbial exposures modulate the gut microbiome and may provide anxiolytic benefits in mice. Sci. Total Environ. 2019, 701, 134684. [Google Scholar] [CrossRef]
- Perofsky, A.C.; Lewis, R.J.; Meyers, L.A. Terrestriality and bacterial transfer: A comparative study of gut microbiomes in sympatric Malagasy mammals. ISME J. 2019, 13, 50–63. [Google Scholar] [CrossRef] [Green Version]
- Van Veelen, H.P.J.; Salles, J.F.; Tieleman, B.I. Multi-level comparisons of cloacal, skin, feather and nest-associated microbiota suggest considerable influence of horizontal acquisition on the microbiota assembly of sympatric woodlarks and skylarks. Microbiome 2017, 5, 156. [Google Scholar] [CrossRef] [Green Version]
- Koch, H.; Schmid-Hempel, P. Gut microbiota instead of host genotype drive the specificity in the interaction of a natural host-parasite system. Ecol. Lett. 2012, 15, 1095–1103. [Google Scholar] [CrossRef]
- Grond, K.; Ryu, H.; Baker, A.J.; Santo Domingo, J.W.; Buehler, D.M. Gastro-intestinal microbiota of two migratory shorebird species during spring migration staging in Delaware Bay, USA. J. Ornithol. 2014, 155, 969–977. [Google Scholar] [CrossRef]
- Ryu, H.; Grond, K.; Verheijen, B.; Elk, M.; Buehler, D.M.; Santo Domingoa, J.W. Intestinal microbiota and species diversity of Campylobacter and Helicobacter spp. in migrating shorebirds in delaware bay. Appl. Environ. Microbiol. 2014, 80, 1838–1847. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Zhou, L.Z.; Zhao, N.N. Temporal-spatial patterns of intestinal parasites of the Hooded Crane (Grus monacha) wintering in lakes of the middle and lower Yangtze River floodplain. Avian Res. 2014, 5, 40–48. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.L. The Infection Status Survey of Main Immunosuppressive Disease Pathogen in Northeast Wild Birds and Yeast Two-Hybrid Screening for the Interactional Host Protein with Revpr. Ph.D. Thesis, Northeast Forestry University, Harbin, China, 2014. [Google Scholar]
- Alm, E.W.; Daniels-Witt, Q.R.; Learman, D.R.; Ryu, H.; Jordan, D.W.; Gehring, T.M.; Domingo, J.S. Potential for gulls to transport bacteria from human waste sites to beaches. Sci. Total Environ. 2018, 615, 123–130. [Google Scholar] [CrossRef]
- Yang, L.; Zhou, L.Z.; Song, Y.W. The effects of food abundance and disturbance on foraging flock patterns of the wintering Hooded Crane (Grus monacha). Avian Res. 2015, 6, 178–185. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.J.; Fraser, J.D.; Chen, J.K. Wintering waterbirds in the middle and lower Yangtze River floodplain: Changes in abundance and distribution. Bird Conserv. Int. 2017, 1, 1–20. [Google Scholar] [CrossRef]
- Liu, J. Diet and Activities of Wintering Tundra Bean Geese (Anser fabalis serrirostris) at Shengjin Lake, NNR, Anhui Province. Master’s Thesis, University of Science and Technology of China, Hefei, China, 2011. [Google Scholar]
- Zhao, F.T.; Zhou, L.Z.; Xu, W.B. Habitat utilization and resource partitioning of wintering Hooded Cranes and three goose species at Shengjin Lake. Avian Res. 2013, 4, 281–290. [Google Scholar] [CrossRef]
- Kreisinger, J.; Kropácková, L.; Petrželková, A.; Adámková, M.; Tomášek, O.; Martin, J.F.; Michálková, R.; Albrecht, T. Temporal stability and the effect of transgenerational transfer on fecal microbiota structure in a long distance migratory bird. Front. Microbiol. 2017, 8, 50. [Google Scholar] [CrossRef]
- Cheng, L.J.; Xue, B.; Yao, S.C.; Liu, J.L. Response of Cladocera fauna to environmental change based on sediments from Shengjin Lake, a Yangtze River-connected lake in China. Quat. Int. 2020, 536, 52–59. [Google Scholar] [CrossRef]
- Dong, Y.Q. Study on Variation and its Determinants in Gut Microbial Communities of Hooded Crane (Grus monacha) over Spatial-Temporal Scales. Ph.D. Thesis, Anhui University, Hefei, China, 2019. [Google Scholar] [CrossRef]
- Zhou, B.; Zhou, L.Z.; Chen, J.Y.; Cheng, Y.Q.; Xu, W.B. Diurnal time-activity budgets of wintering Hooded Cranes (Grus monacha) in Shengjin Lake, China. Waterbirds 2010, 33, 110–115. [Google Scholar] [CrossRef]
- Saitoh, T.; Sugita, N.; Someya, S.; Iwami, Y.; Kobayashi, S.; Kamigaich, H.; Higuch, A.; Asai, S.; Yamamoto, Y.; Nishiumi, I. DNA barcoding reveals 24 distinct lineages as cryptic bird species candidates in and around the Japanese Archipelago. Mol. Ecol. Resour. 2014, 15, 177–186. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibert, C.; Escarguel, G. PER-SIMPER—A new tool for inferring community assembly processes from taxon occurrences. Glob. Ecol. Biogeogr. 2019, 28, 374–385. [Google Scholar] [CrossRef]
- Ignatov, A.N.; Khodykina, M.V.; Vinogradova, S.V.; Polityko, V.A.; Kornev, K.P.; Mazurin, E.S. First report of Agrobacterium vitis causing crown galls of wine grape in Russia. Plant. Dis. 2016, 100, 853–854. [Google Scholar] [CrossRef]
- Zhang, F.L.; Xiang, X.J.; Dong, Y.Q.; Yan, S.F.; Song, Y.W.; Zhou, L.Z. Significant differences in the gut bacterial communities of Hooded Crane (Grus monacha) in different seasons at a stopover site on the flyway. Animals 2020, 10, 701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, T.M.; Rogers, T.L.; Carlini, A.R.; Brown, M.V. Diet and phylogeny shape the gut microbiota of Antarctic seals: A comparison of wild and captive animals. Environ. Microbiol. 2013, 15, 1132–1145. [Google Scholar] [CrossRef] [PubMed]
- Hird, S.M.; Sánchez, C.; Carstens, B.C.; Brumfield, R.T. Comparative gut microbiota of 59 neotropical Bird Species. Front. Microbiol. 2015, 6, 1403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banks, J.C.; Cary, S.C.; Hogg, I.D. The phylogeography of Adelie Penguin faecal flora. Environ. Microbiol. 2009, 11, 577–588. [Google Scholar] [CrossRef]
- Fu, R.; Xiang, X.J.; Dong, Y.Q.; Cheng, L.; Zhou, L.Z. Comparing the intestinal bacterial communities of sympatric wintering Hooded Crane (Grus monacha) and Domestic Goose (Anser anser domesticus). Avian Res. 2020, 11, 106–114. [Google Scholar] [CrossRef]
- Zhao, M.J. Wintering Population Ecology of Greater White-Fronted Goose and Bean Goose Along the Middle and Lower Yangtze River Floodplain. Ph.D. Thesis, University of Science and Technology of China, Hefei, China, 2017. [Google Scholar]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [Green Version]
- Wan, W.J.; Zhou, L.Z.; Song, Y.W. Shifts in foraging behavior of wintering Hooded Cranes (Grus monacha) in three different habitats at Shengjin Lake, China. Avian Res. 2016, 7, 167–173. [Google Scholar] [CrossRef]
- Midani, F.S.; Weil, A.A.; Chowdhury, F.; Begum, Y.A.; Khan, A.I.; Debela, M.D.; Durand, H.K.; Reese, A.T.; Nimmagadda, S.N.; Silverman, J.D.; et al. Human gut microbiota predicts susceptibility to Vibrio cholerae infection. J. Infect. Dis. 2018, 218, 645–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, T.; Kawaoka, Y. Host-range barrier of influenza a viruses. Vet. Microbiol. 2000, 74, 71–75. [Google Scholar] [CrossRef]
- Karesh, W.B.; Cook, R.A.; Bennett, E.L.; Newcomb, J. Wildlife trade and global disease emergence. Emerg. Infect. Dis. 2005, 11, 1000–1002. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Phyla | Average Relative Abundance 1 (%) | Significance 2 | |
|---|---|---|---|
| HC | BG | ||
| Firmicutes | 64.93 (22.30) a | 68.67 (24.90) a | 0.410 |
| Proteobacteria | 20.80 (16.02) a | 20.81 (18.27) a | 0.999 |
| Actinobacteria | 8.28 (6.76) a | 5.86 (6.15) b | 0.049 |
| Bacteroidetes | 1.77 (4.86) a | 1.81 (3.54) a | 0.723 |
| Host Variable | Period Variable | ||||
|---|---|---|---|---|---|
| R | p | R | p | ||
| HCE VS. BGE | 0.374 | 0.001 | HCE VS. HCM | 0.748 | 0.001 |
| HCM VS. BGM | 0.720 | 0.001 | HCM VS. HCL | 0.429 | 0.001 |
| HCL VS. BGL | 0.250 | 0.001 | BGE VS. BGM | 0.327 | 0.001 |
| HC VS. BG | 0.259 | 0.001 | BGM VS. BGL | 0.052 | 0.194 |
| Contribution (%) | |||||||
|---|---|---|---|---|---|---|---|
| Taxa | HCE vs. BGE | HCM vs. BGM | HCL vs. BGL | HCE vs. HCM | HCM vs. HCL | BGE vs. BGM | BGM vs. BGL |
| g__Lactobacillus | 12.83 | 37.02 | 14.61 | 18.29 | 26.28 | 20.68 | 19.86 |
| g__Solibacillus | 7.27 | — | 2.88 | 5.08 | 2.59 | 5.96 | — |
| f__Peptostreptococcaceae | 7.19 | 1.89 | — | 5.89 | — | 4.81 | 2.09 |
| g__Exiguobacterium | 3.09 | — | — | — | — | — | — |
| g__Streptococcus | 2.53 | — | — | — | — | 3.42 | 1.48 |
| f__Enterococcaceae | — | 4.43 | — | — | — | 4.88 | 5.22 |
| g__Bacillus | — | 1.49 | 1.03 | — | 2.29 | — | — |
| g__Agrobacterium | — | 1.39 | — | — | 1.56 | — | — |
| g__Clostridium | — | — | 4.28 | — | — | — | 4.60 |
| s__F.prausnitzii | — | — | 1.03 | — | — | — | — |
| f__Nitrosomonadaceae | — | — | — | 2.17 | — | — | — |
| g__Paenibacillus | — | — | — | 1.91 | 1.18 | — | — |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Zhou, L. Is Intestinal Bacterial Diversity Enhanced by Trans-Species Spread in the Mixed-Species Flock of Hooded Crane (Grus monacha) and Bean Goose (Anser fabalis) Wintering in the Lower and Middle Yangtze River Floodplain? Animals 2021, 11, 233. https://doi.org/10.3390/ani11010233
Yang Z, Zhou L. Is Intestinal Bacterial Diversity Enhanced by Trans-Species Spread in the Mixed-Species Flock of Hooded Crane (Grus monacha) and Bean Goose (Anser fabalis) Wintering in the Lower and Middle Yangtze River Floodplain? Animals. 2021; 11(1):233. https://doi.org/10.3390/ani11010233
Chicago/Turabian StyleYang, Zhuqing, and Lizhi Zhou. 2021. "Is Intestinal Bacterial Diversity Enhanced by Trans-Species Spread in the Mixed-Species Flock of Hooded Crane (Grus monacha) and Bean Goose (Anser fabalis) Wintering in the Lower and Middle Yangtze River Floodplain?" Animals 11, no. 1: 233. https://doi.org/10.3390/ani11010233
APA StyleYang, Z., & Zhou, L. (2021). Is Intestinal Bacterial Diversity Enhanced by Trans-Species Spread in the Mixed-Species Flock of Hooded Crane (Grus monacha) and Bean Goose (Anser fabalis) Wintering in the Lower and Middle Yangtze River Floodplain? Animals, 11(1), 233. https://doi.org/10.3390/ani11010233