
miR-152 Regulates Bovine Myoblast Proliferation by Targeting KLF6

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Samples Collection
2.2. Cell Culture and Transfection
2.3. RNA Extraction and Quantitative Real-Time PCR
2.4. Western Blotting
2.5. EdU Assay
2.6. Cell Cycle Assay
2.7. Dual-Luciferase Reporter Assay
2.8. Statistical Analyses
3. Results
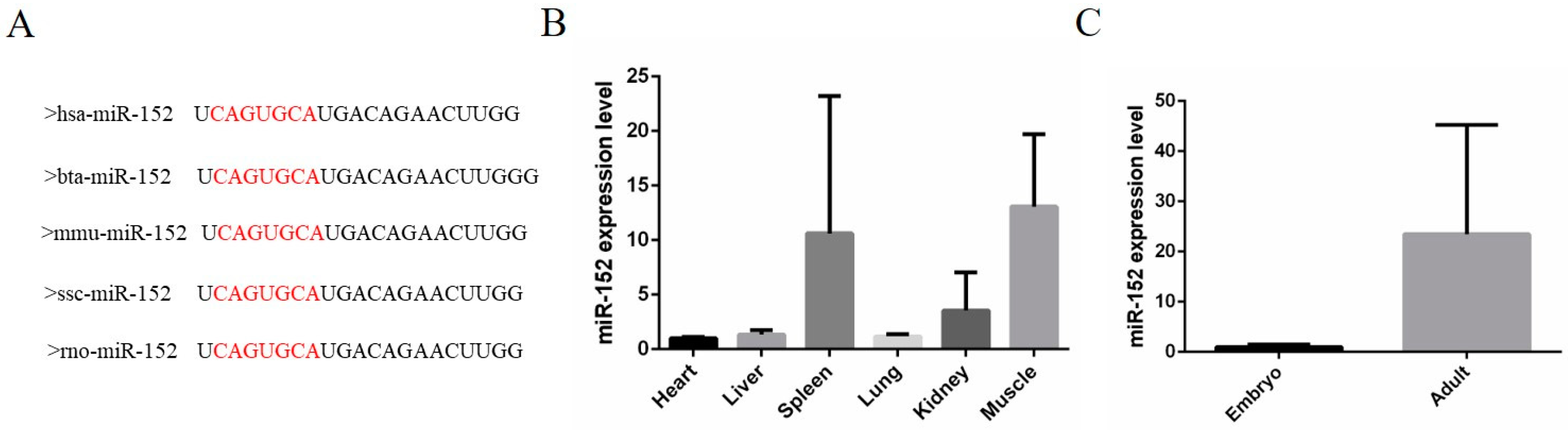
3.1. The Expression Characteristics of miR-152 in Cattle Tissues
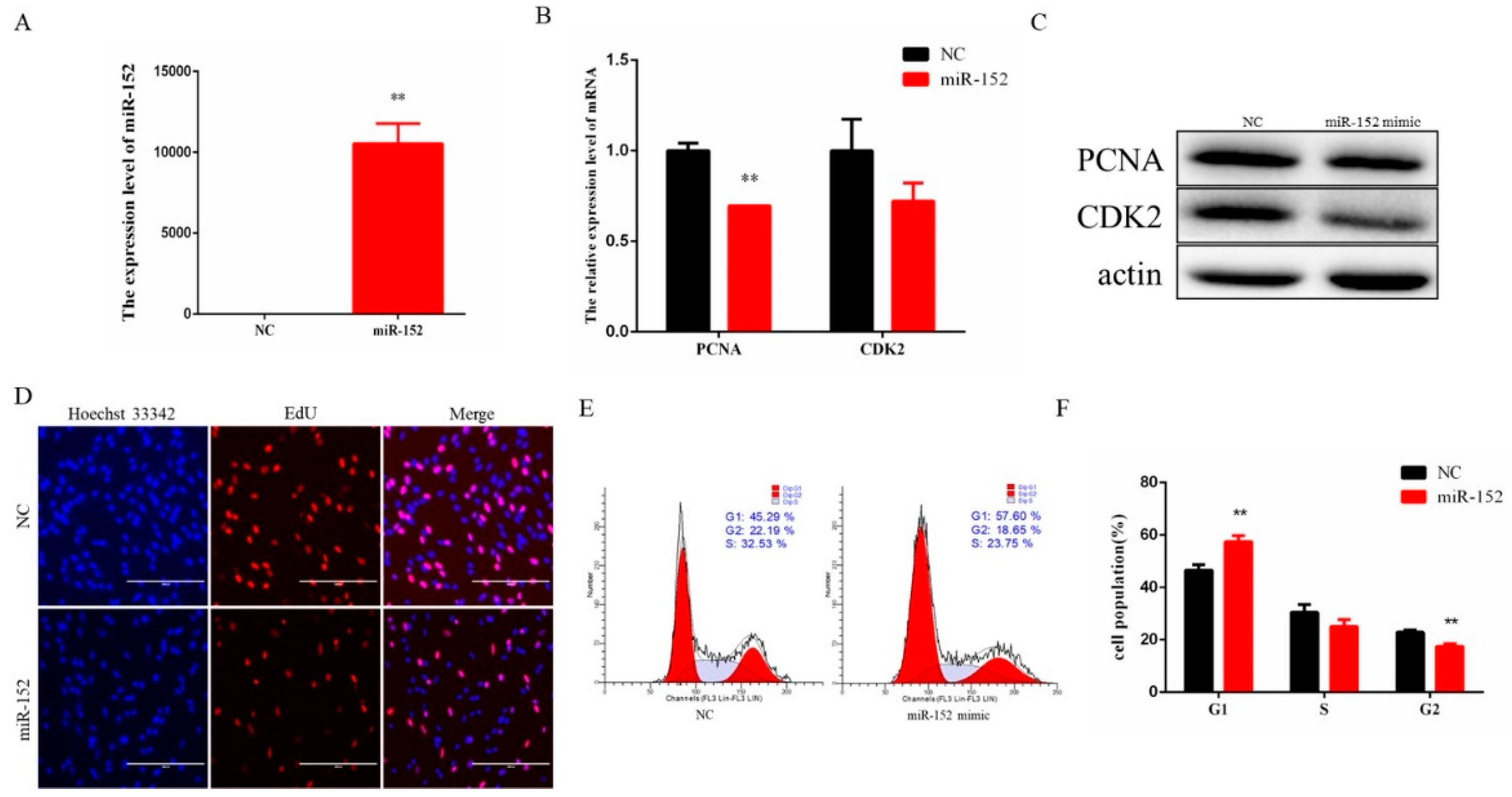
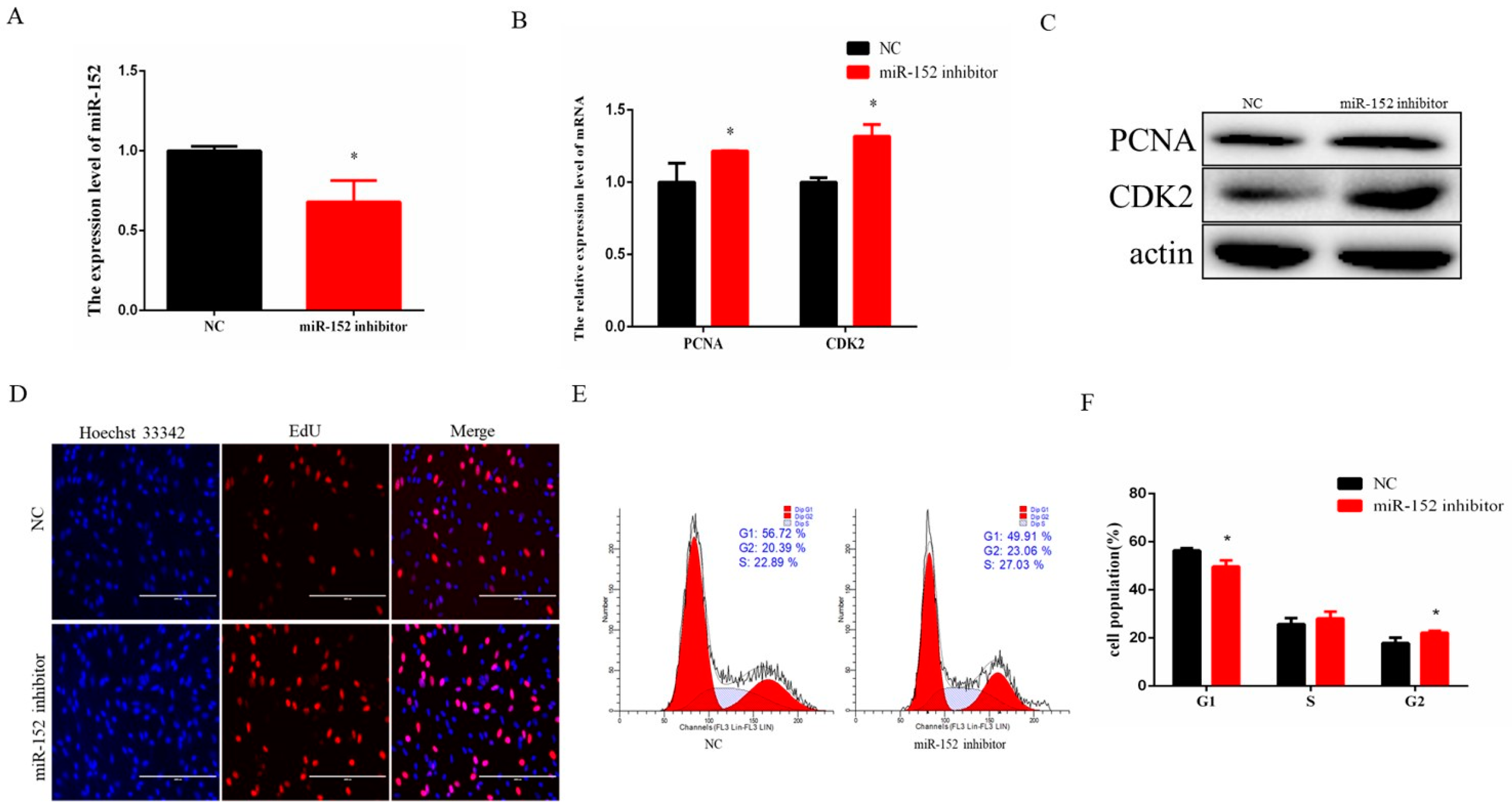
3.2. miR-152 Regulates Bovine Myoblast Proliferation
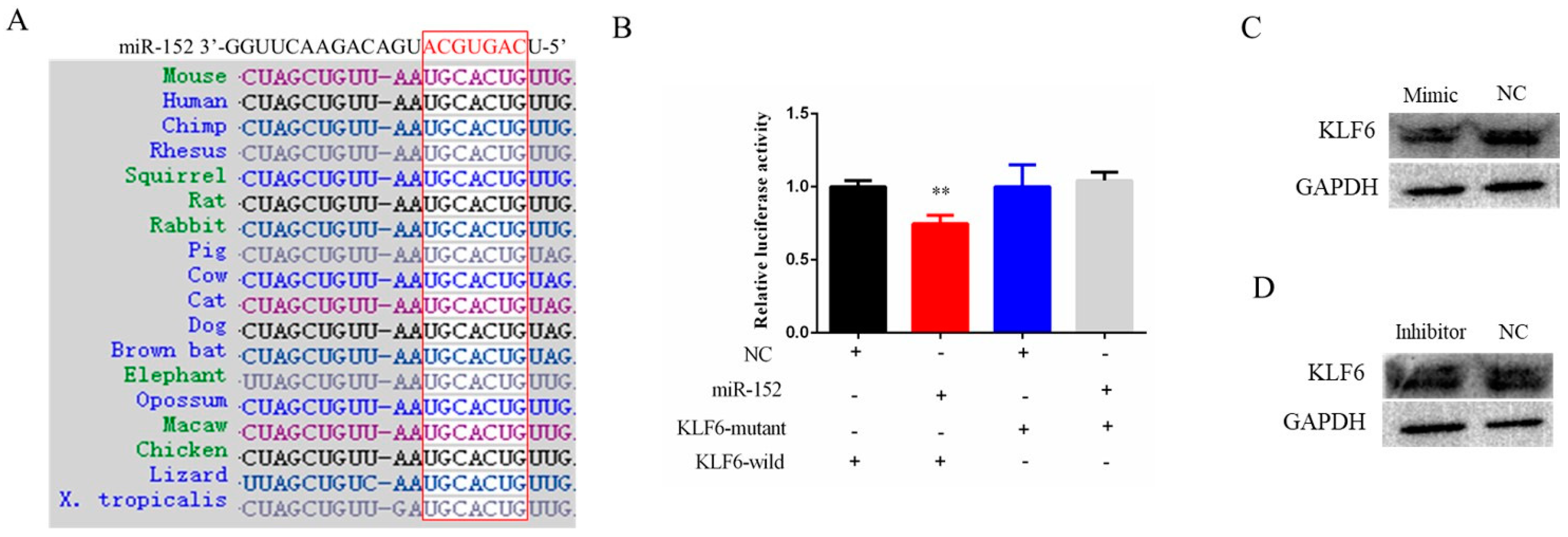
3.3. KLF6 Is a Target Gene of miR-152
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sabourin, L.A.; Rudnicki, M.A. The molecular regulation of myogenesis. Clin. Genet. 2010, 57, 16. [Google Scholar] [CrossRef] [PubMed]
- Bentzinger, C.F.; Wang, Y.X.; Rudnicki, M.A. Building muscle: Molecular regulation of myogenesis. Cold Spring Harb. Perspect. Perspect. Perspect. Biol. 2012, 4, a008342. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Yan, X.; Tong, J.F.; Zhao, J.; Zhu, M.J. Maternal obesity, inflammation, and fetal skeletal muscle development. Biol. Reprod. 2010, 82, 4–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaney, K.; Kasprzycka, P.; Ciemerych, M.A.; Zimowska, M. The role of TGF-β1 during skeletal muscle regeneration. Cell Biol. Int. 2017, 41, 706–715. [Google Scholar] [CrossRef] [Green Version]
- Anderson, K.J.; Russell, A.P.; Foletta, V.C. NDRG2 promotes myoblast proliferation and caspase 3/7 activities during differentiation, and attenuates hydrogen peroxide—But not palmitate-induced toxicity. FEBS Open Bio 2015, 5, 668–681. [Google Scholar] [CrossRef] [Green Version]
- Alteri, A.; De Vito, F.; Messina, G.; Pompili, M.; Calconi, A.; Visca, P.; Mottolese, M.; Presutti, C.; Grossi, M. Cyclin D1 is a major target of miR-206 in cell differentiation and transformation. Cell Cycle 2013, 12, 3781–3790. [Google Scholar] [CrossRef] [Green Version]
- Hao, D.; Wang, X.; Wang, X.; Thomsen, B.; Yang, Y.; Lan, X.; Huang, Y.; Chen, H. MicroRNA bta-miR-365-3p inhibits proliferation but promotes differentiation of primary bovine myoblasts by targeting the activin A receptor type I. J. Anim. Sci. Biotechnol. 2021, 12, 16. [Google Scholar] [CrossRef]
- Li, X.; Qiu, J.; Liu, H.; Deng, Y.; Hu, S.; Hu, J.; Wang, Y.; Wang, J. MicroRNA-33a negatively regulates myoblast proliferation by targeting IGF1, follistatin and cyclin D1. Biosci. Rep. 2020, 40, BSR20191327. [Google Scholar] [CrossRef]
- Filipowicz, W.; Bhattacharyya, S.N.; Sonenberg, N. Mechanisms of post-transcriptional regulation by microRNAs: Are the answers in sight? Nat. Rev. Genet. 2008, 9, 102–114. [Google Scholar] [CrossRef]
- Chen, J.F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; Callis, T.E.; Hammond, S.M.; Conlon, F.L.; Wang, D.Z. The role of microRNA-1 and microRNA-133 in skeletal muscle proliferation and differentiation. Nat. Genet. 2006, 38, 228–233. [Google Scholar] [CrossRef]
- Motohashi, N.; Alexander, M.S.; Shimizu-Motohashi, Y.; Myers, J.A.; Kawahara, G.; Kunkel, L.M. Regulation of IRS1/Akt insulin signaling by microRNA-128a during myogenesis. J. Cell Sci. 2013, 126, 2678–2691. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Yue, B.; Lan, X.; Wang, Y.; Fang, X.; Ma, Y.; Bai, Y.; Qi, X.; Zhang, C.; Chen, H. MiR-499 regulates myoblast proliferation and differentiation by targeting transforming growth factor beta receptor 1. J. Cell. Physiol. 2019, 234, 2523–2536. [Google Scholar] [CrossRef]
- Qiu, H.; Liu, N.; Luo, L.; Zhong, J.; Tang, Z.; Kang, K.; Qu, J.; Peng, W.; Liu, L.; Li, L.; et al. MicroRNA-17-92 regulates myoblast proliferation and differentiation by targeting the ENH1/Id1 signaling axis. Cell Death Differ. 2016, 23, 1658–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Chen, X.; Yu, B.; He, J.; Chen, D. MicroRNA-27a promotes myoblast proliferation by targeting myostatin. Biochem. Biophys. Res. Commun. 2012, 423, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Nie, M.; Deng, Z.L.; Liu, J.; Wang, D.Z. Noncoding RNAs, Emerging Regulators of Skeletal Muscle Development and Diseases. BioMed Res. Int. 2015, 2015, 676575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Sonstegard, T.S.; Li, C.; Huang, Y.; Li, Z.; Lan, X.; Zhang, C.; Lei, C.; Zhao, X.; Chen, H. Altered microRNA expression in bovine skeletal muscle with age. Anim. Genet. 2015, 46, 227–238. [Google Scholar] [CrossRef]
- Song, C.; Yang, Z.; Jiang, R.; Cheng, J.; Yue, B.; Wang, J.; Sun, X.; Huang, Y.; Lan, X.; Lei, C.; et al. lncRNA IGF2 AS Regulates Bovine Myogenesis through Different Pathways. Mol. Ther. Nucleic Acids 2020, 21, 874–884. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, J.; Qin, F.; Dai, S. miR-152 as a tumor suppressor microRNA: Target recognition and regulation in cancer. Oncol. Lett. 2016, 11, 3911–3916. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Zhang, Y.; Qi, Y.; Yu, D.; Shao, Q.; Liang, J. MicroRNA-152 inhibits tumor cell growth by directly targeting RTKN in hepatocellular carcinoma. Oncol. Rep. 2017, 37, 1227–1234. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Li, J.; Ding, Q.; Cheng, G.; Zhou, H.; Tao, L.; Cai, H.; Li, P.; Cao, Q.; Ju, X.; et al. miR-152 controls migration and invasive potential by targeting TGFalpha in prostate cancer cell lines. Prostate 2013, 73, 1082–1089. [Google Scholar] [CrossRef]
- Tsuruta, T.; Kozaki, K.; Uesugi, A.; Furuta, M.; Hirasawa, A.; Imoto, I.; Susumu, N.; Aoki, D.; Inazawa, J. miR-152 is a tumor suppressor microRNA that is silenced by DNA hypermethylation in endometrial cancer. Cancer Res. 2011, 71, 6450–6462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, Y.; Ma, N.; Wang, D.; Zhang, Y.; Zhou, J.; Wu, G.; Zhao, R.; Huang, H.; Wang, X.; Qiao, Y.; et al. MiR-152 and miR-185 co-contribute to ovarian cancer cells cisplatin sensitivity by targeting DNMT1 directly: A novel epigenetic therapy independent of decitabine. Oncogene 2014, 33, 378–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.; Yao, Y.; Wang, P.; Liu, Y.; Zhao, L.; Li, Z.; Li, Z.; Xue, Y. MiR-152 functions as a tumor suppressor in glioblastoma stem cells by targeting Kruppel-like factor 4. Cancer Lett. 2014, 355, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.S.; Li, B.J.; Lu, H.W.; Chen, Y.; Lu, C.; Zhu, R.X.; Liu, S.H.; Yi, Q.T.; Li, J.; Song, C.H. Serum miR-152, miR-148a, miR-148b, and miR-21 as novel biomarkers in non-small cell lung cancer screening. Tumour Biol. 2015, 36, 3035–3042. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Fang, X.; Yang, R.; Yu, H.; Jiang, P.; Sun, B.; Zhao, Z. MiR-152 Regulates Apoptosis and Triglyceride Production in MECs via Targeting ACAA2 and HSD17B12 Genes. Sci. Rep. 2018, 8, 417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, M.; Hansen, J.H.; Hedegaard, J.; Nielsen, R.O.; Panitz, F.; Bendixen, C.; Thomsen, B. MicroRNA identity and abundance in porcine skeletal muscles determined by deep sequencing. Anim. Genet. 2010, 41, 159–168. [Google Scholar] [CrossRef]
- Gan, M.; Du, J.; Shen, L.; Yang, D.; Jiang, A.; Li, Q.; Jiang, Y.; Tang, G.; Li, M.; Wang, J.; et al. miR-152 regulates the proliferation and differentiation of C2C12 myoblasts by targeting E2F3. In Vitro Cell. Dev. Biol. Anim. 2018, 54, 304–310. [Google Scholar] [CrossRef]
- Narla, G.; Heath, K.E.; Reeves, H.L.; Li, D.; Giono, L.E.; Kimmelman, A.C.; Glucksman, M.J.; Narla, J.; Eng, F.J.; Chan, A.M.; et al. KLF6, a candidate tumor suppressor gene mutated in prostate cancer. Science 2001, 294, 2563–2566. [Google Scholar] [CrossRef]
- Cho, Y.G.; Kim, C.J.; Park, C.H.; Yang, Y.M.; Kim, S.Y.; Nam, S.W.; Lee, S.H.; Yoo, N.J.; Lee, J.Y.; Park, W.S. Genetic alterations of the KLF6 gene in gastric cancer. Oncogene 2005, 24, 4588–4590. [Google Scholar] [CrossRef] [Green Version]
- Kang, T.; Xing, W.; Xi, Y.; Chen, K.; Zhan, M.; Tang, X.; Wang, Y.; Zhang, R.; Lei, M. MiR-543 regulates myoblast proliferation and differentiation of C2C12 cells by targeting KLF6. J. Cell. Biochem. 2020, 12, 4827–4837. [Google Scholar] [CrossRef]
- Song, C.; Yang, J.; Jiang, R.; Yang, Z.; Li, H.; Huang, Y.; Lan, X.; Lei, C.; Ma, Y.; Qi, X.; et al. miR-148a-3p regulates proliferation and apoptosis of bovine muscle cells by targeting KLF6. J. Cell. Physiol. 2019, 234, 15742–15750. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, C.; Fang, X.; Yang, Z.; Wang, Q.; Meng, F.; Chen, Y.; Chen, J.; Zhao, B.; Wang, Y.; Fang, X.; et al. miR-152 Regulates Bovine Myoblast Proliferation by Targeting KLF6. Animals 2021, 11, 3001. https://doi.org/10.3390/ani11103001
Song C, Fang X, Yang Z, Wang Q, Meng F, Chen Y, Chen J, Zhao B, Wang Y, Fang X, et al. miR-152 Regulates Bovine Myoblast Proliferation by Targeting KLF6. Animals. 2021; 11(10):3001. https://doi.org/10.3390/ani11103001
Chicago/Turabian StyleSong, Chengchuang, Xue Fang, Zhaoxin Yang, Qi Wang, Fantong Meng, Yaqi Chen, Junhao Chen, Bei Zhao, Yanhong Wang, Xingtang Fang, and et al. 2021. "miR-152 Regulates Bovine Myoblast Proliferation by Targeting KLF6" Animals 11, no. 10: 3001. https://doi.org/10.3390/ani11103001
APA StyleSong, C., Fang, X., Yang, Z., Wang, Q., Meng, F., Chen, Y., Chen, J., Zhao, B., Wang, Y., Fang, X., Gu, L., & Zhang, C. (2021). miR-152 Regulates Bovine Myoblast Proliferation by Targeting KLF6. Animals, 11(10), 3001. https://doi.org/10.3390/ani11103001