
Ion Channel and Ubiquitin Differential Expression during Erythromycin-Induced Anhidrosis in Foals



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
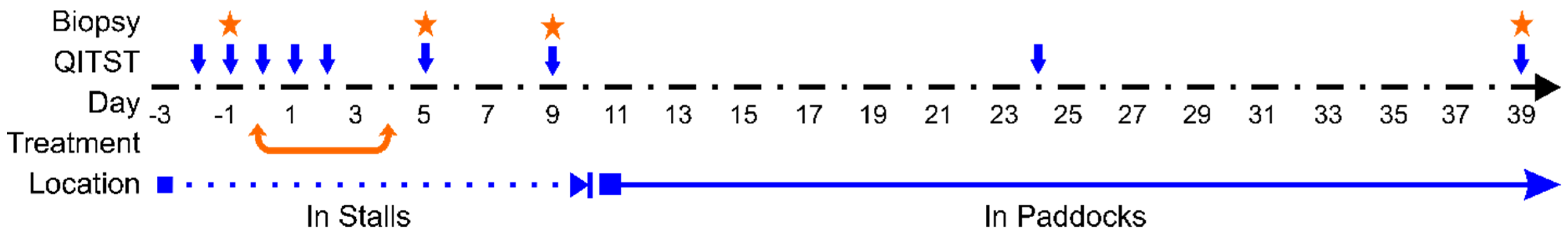
2.1. Experimental Animals and Design
2.2. Erythromycin Treatment
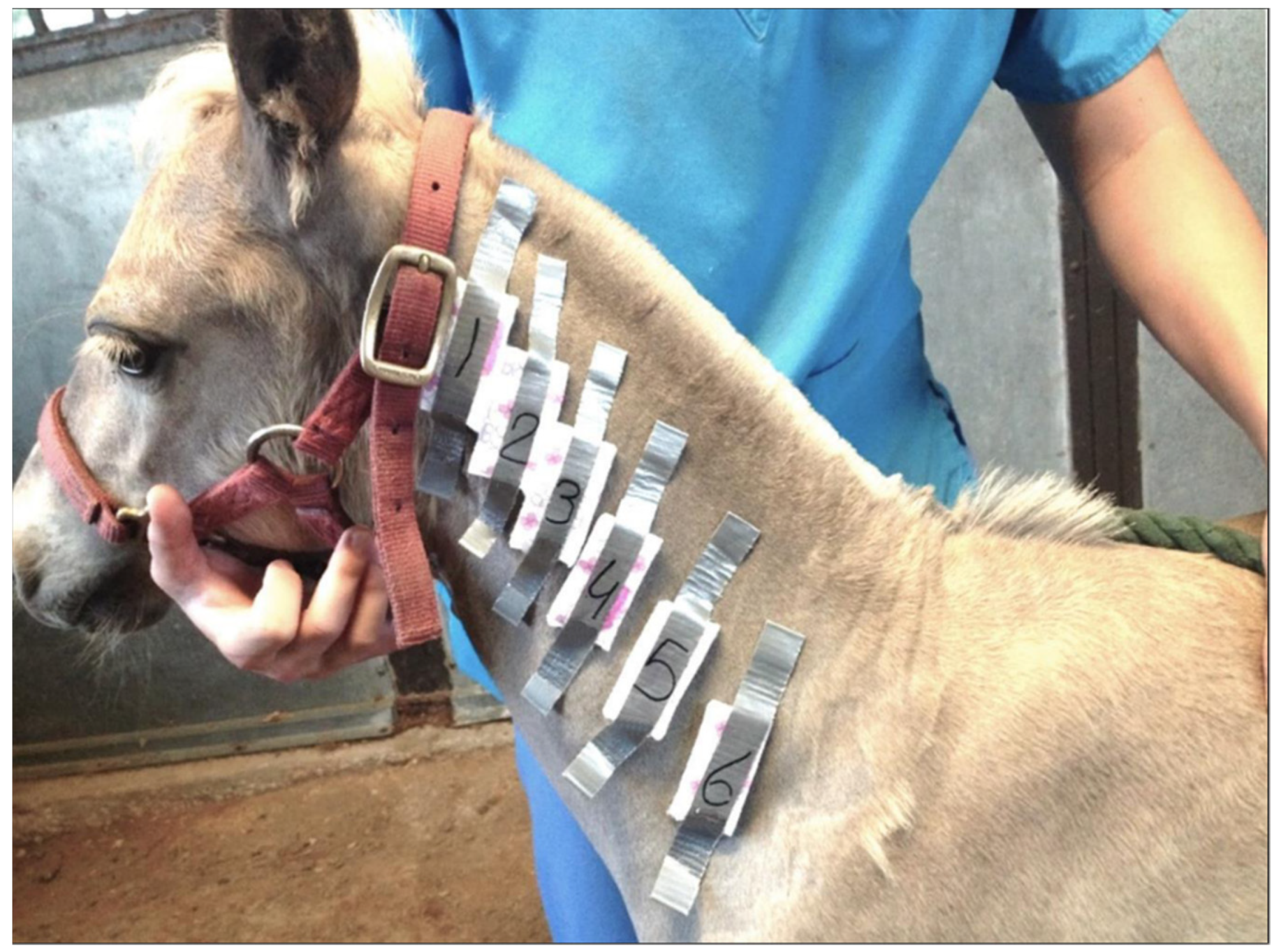
2.3. Quantitative Sweat Tests
2.4. Biopsy Collection
2.5. Illumina Sequencing and Library Construction
2.6. RNA-seq Read Quality Control and Mapping
2.7. Differential Expression Analysis
2.8. Differentially Expressed Gene Ontology and Literature Search
3. Results
3.1. Erythromycin Causes Hyperthermia and Diarrhea
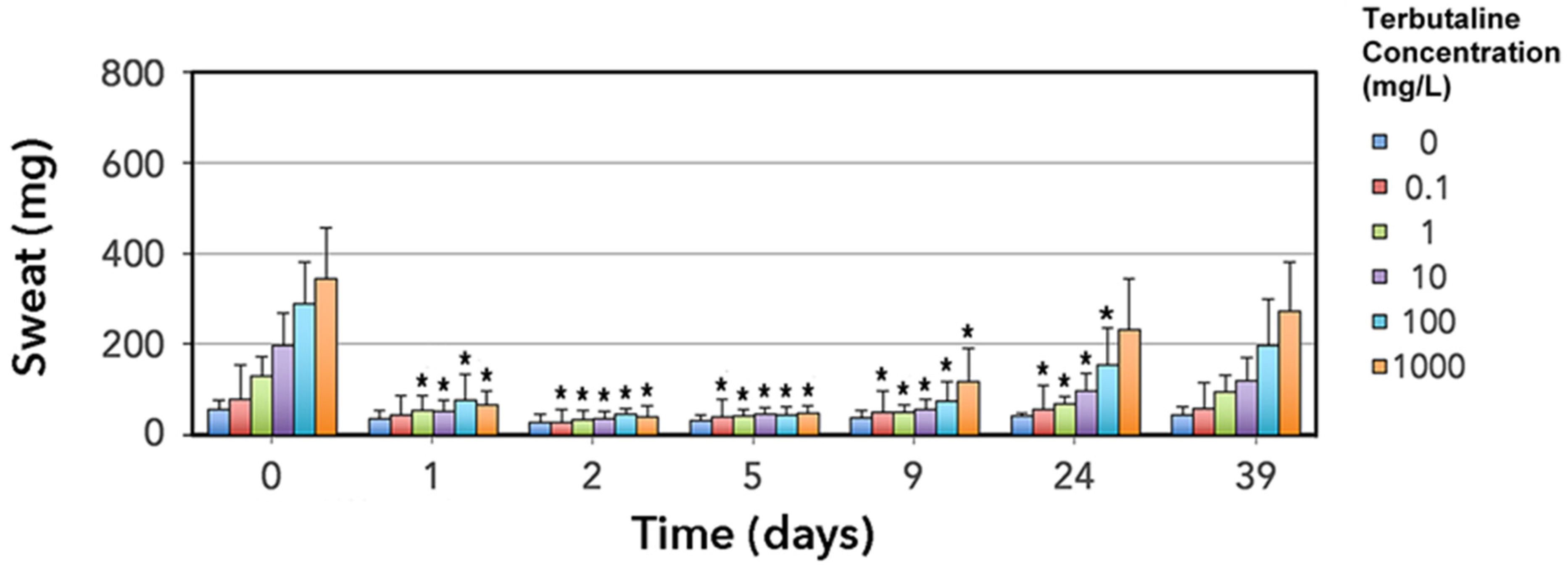
3.2. Thermoregulatory Sweat Response Is Impaired by Erythromycin
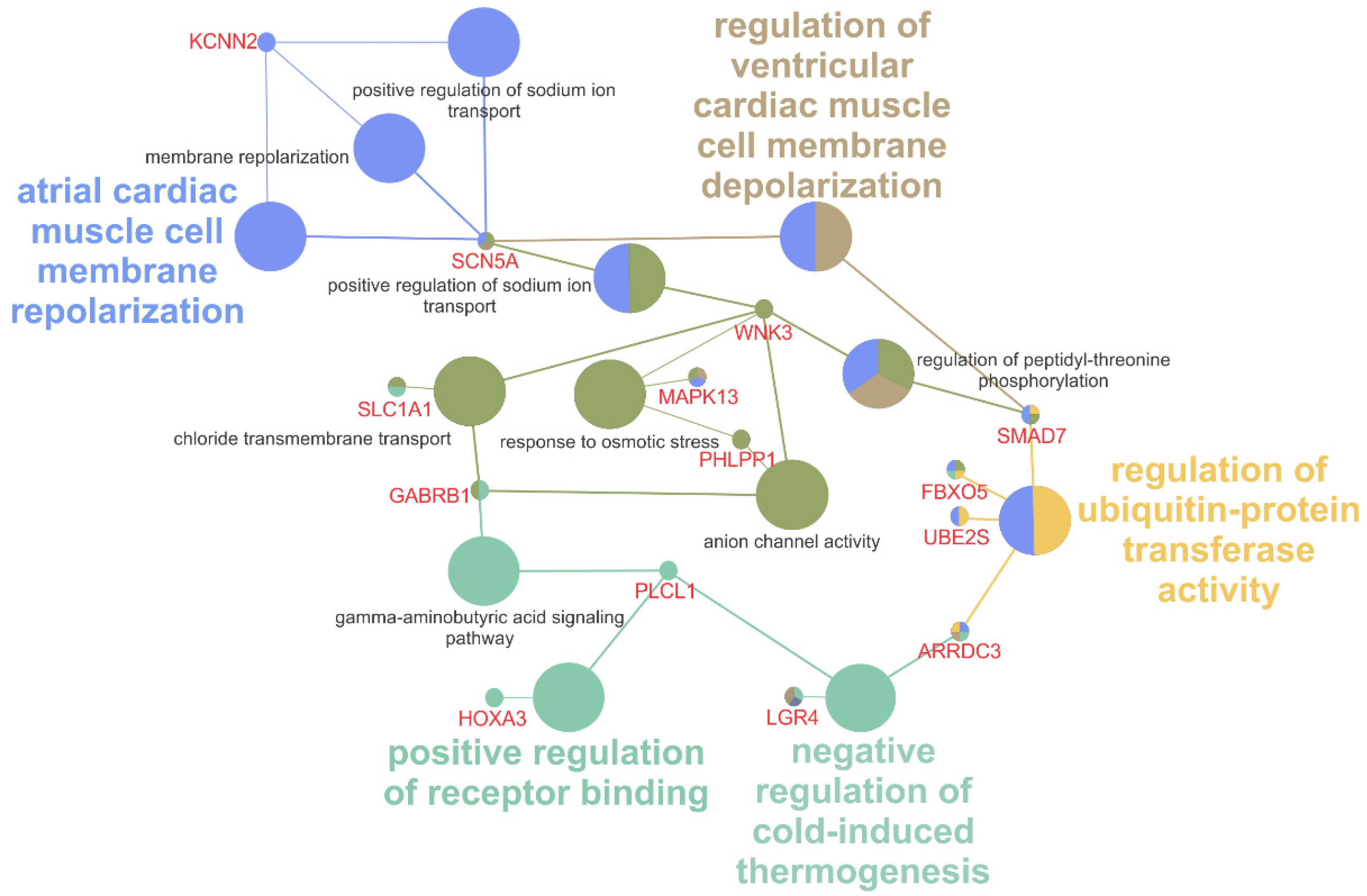
3.3. Differentially Expressed Genes Are Involved in Mitosis, Ion Channel Function, Ubiquitin Activity, and Immune Response
4. Discussion
4.1. Erythromycin Activation of SK2 Channel as a Cause of Diarrhea
4.2. Upregulation of Ion Channels and β2-Adrenergic Receptors by Erythromycin in Thermoregulatory Sweat
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Giguère, S.; Prescott, J.F. Clinical manifestations, diagnosis, treatment, and prevention of Rhodococcus equi infections in foals. Veter. Microbiol. 1997, 56, 313–334. [Google Scholar] [CrossRef]
- Elissalde, G.; Renshaw, H.W.; A Walberg, J. Corynebacterium equi: An interhost review with emphasis on the foal. Comp. Immunol. Microbiol. Infect. Dis. 1980, 3, 433–445. [Google Scholar] [CrossRef]
- Giguère, S.; Cohen, N.D.; Chaffin, M.K.; Slovis, N.M.; Hondalus, M.K.; Hines, S.A.; Prescott, J.F. Diagnosis, Treatment, Control, and Prevention of Infections Caused by R hodococcus equi in Foals. J. Vet. Intern. Med. 2011, 25, 1209–1220. [Google Scholar] [CrossRef] [PubMed]
- Hondalus, M.K. Pathogenesis and virulence of Rhodococcus equi. Veter. Microbiol. 1997, 56, 257–268. [Google Scholar] [CrossRef]
- Jacks, S.S.; Giguère, S.; Nguyen, A. In vitro susceptibilities of Rhodococcus equi and other common equine pathogens to azithromycin, clarithromycin, and 20 other antimicrobials. Antimicrob. Agents Chemother. 2003, 47, 1742–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, C.; Sweeney, R.; Divers, T. Rhodococcus equi pneumonia in 48 foals: Response to antimicrobial therapy. Veter. Microbiol. 1987, 14, 329–336. [Google Scholar] [CrossRef]
- Hillidge, C.J. Use of erythromycin-rifampin combination in treatment of Rhodococcus equi pneumonia. Veter. Microbiol. 1987, 14, 337–342. [Google Scholar] [CrossRef]
- Prokesch, R.C.; Hand, W.L. Antibiotic entry into human polymorphonuclear leukocytes. Antimicrob. Agents Chemother. 1982, 21, 373–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stratton-Phelps, M.; Wilson, W.D.; Gardner, I.A. Risk of adverse effects in pneumonic foals treated with erythromycin versus other antibiotics: 143 cases (1986–1996). J. Am. Veter. Med. Assoc. 2000, 217, 68–73. [Google Scholar] [CrossRef]
- Simon, H.B. Hyperthermia. N. Engl. J. Med. 1993, 329, 483–487. [Google Scholar] [PubMed]
- Traub-Dargatz, J.; Wilson, W.D.; Conboy, H.S. Hyperthermia in Foals Treated with Erythromycin Alone or in Combination with Rifampin for Respiratory Disease during Hot Environmental Conditions. Proc. Am. Assoc. Equine Pract. 1996, 42, 243–244. [Google Scholar]
- Lakritz, J.; Wilson, W.D.; Watson, J.L.; Hyde, D.M.; Mihalyi, J.; Plopper, C.G. Effect of treatment with erythromycin on bronchoalveolar lavage fluid cell populations in foals. Am. J. Veter. Res. 1997, 58, 56–61. [Google Scholar]
- McDonald, R.E.; Fleming, R.I.; Beeley, J.G.; Bovell, D.L.; Lu, J.R.; Zhao, X.; Cooper, A.; Kennedy, M.W. Latherin: A Surfactant Protein of Horse Sweat and Saliva. PLoS ONE 2009, 4, e5726. [Google Scholar] [CrossRef]
- Hinchcliff, K.W.; Kaneps, A.J.; Geor, R.J. Equine Exercise Physiology: The Science of Exercise in the Athletic Horse. In Elsevier Health Sciences; Saunders/Elsevier: Edinburgh, UK, 2008; Print. [Google Scholar]
- Breuhaus, B. Thyroid Function in Anhidrotic Horses. J. Veter. Intern. Med. 2009, 23, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, D.M.; Elder, H.Y.; Bovell, D.L. Equine sweating and anhidrosis Part 1--equine sweating. Veter. Dermatol. 2006, 17, 361–392. [Google Scholar] [CrossRef] [PubMed]
- Currie, A.K.; Seager, S.W.J. Anhidrosis. In Proceedings of the Annual Convention of the American Association of Equine Practitioners; American Association of Equine Practitioners: Lexington, KY, USA, 1976; Volume 22, pp. 249–251. [Google Scholar]
- Geor, R.J.; McCutcheon, L.J. Thermoregulation and clinical disorders associated with exercise and heat stress. Compend. Contin. Educ. Pract. Vet. 1996, 18, 436. [Google Scholar]
- Stewart, A.; Sanchez, L.C.; Mallicote, M.F.; Muniz, A.L.; Westerterp, M.S.; Burrow, J.A.; MacKAY, R.J. Effects of clarithromycin, azithromycin and rifampicin on terbutaline-induced sweating in foals. Equine Veter. J. 2017, 49, 624–628. [Google Scholar] [CrossRef] [PubMed]
- Stieler, A.L.; Mallicote, M.F.; Martabano, B.B.; Sanchez, L.C.; Burrow, J.A.; Mackay, R.J. Macrolide-induced hyperthermia in foals: Role of impaired sweat responses. Equine Veter. J. 2015, 48, 590–594. [Google Scholar] [CrossRef]
- Smith, B.P.; Van Metre, D.C.; Pusterla, N. Large Animal Internal Medicine, 6th ed.; Elsevier Mosby: St. Louis, MI, USA, 2020. [Google Scholar]
- Mackay, R.J. Quantitative intradermal terbutaline sweat test in horses. Equine Veter. J. 2008, 40, 518–520. [Google Scholar] [CrossRef] [PubMed]
- Mueller, O.; Lightfoot, S.; Schroeder, A. RNA integrity number (RIN)–standardization of RNA quality control. Agil. Appl. Note Publ. 2004, 1, 1–8. [Google Scholar]
- Schroeder, A.; Mueller, O.; Stocker, S.; Salowsky, R.; Leiber, M.; Gassmann, M.; Lightfoot, S.; Menzel, W.; Granzow, M.; Ragg, T. The RIN: An RNA integrity number for assigning integrity values to RNA measurements. BMC Mol. Biol. 2006, 7, 3. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; He, X.; A Hoadley, K.; Parker, J.S.; Hayes, D.N.; Perou, C.M. Comparison of RNA-Seq by poly (A) capture, ribosomal RNA depletion, and DNA microarray for expression profiling. BMC Genom. 2014, 15, 419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penland, S.K.; O Keku, T.; Torrice, C.; He, X.; Krishnamurthy, J.; A Hoadley, K.; Woosley, J.T.; E Thomas, N.; Perou, C.M.; Sandler, R.S.; et al. RNA expression analysis of formalin-fixed paraffin-embedded tumors. Lab. Investig. 2007, 87, 383–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. Babraham bioinformatics—FastQC a quality control tool for high throughput sequence data. Soil 2020, 5, 47–81. [Google Scholar]
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, N.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal RNA-Seq quantification with kallisto. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef] [PubMed]
- Zerbino, D.R.; Achuthan, P.; Akanni, W.; Amode, M.R.; Barrell, D.; Bhai, J.; Billis, K.; Cummins, C.; Gall, A.; Girón, C.G. Ensembl 2018. Nucleic Acids Res. 2017, 46, D754–D761. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Erratum: Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pimentel, H.; Bray, N.L.; Puente, S.; Melsted, P.; Pachter, L. Differential analysis of RNA-seq incorporating quantification uncertainty. Nat. Methods 2017, 14, 687–690. [Google Scholar] [CrossRef]
- Chen, Y.; Lun, A.T.L.; Smyth, G.K. Differential Expression Analysis of Complex RNA-seq Experiments Using edgeR. In Stat. Anal. Next Gener. Seq. Data; Springer International Publishing: Cham, Switzerland, 2014; pp. 51–74. [Google Scholar] [CrossRef] [Green Version]
- Lun, A.T.L.; Chen, Y.; Smyth, G.K. It’s DE-licious: A Recipe for Differential Expression Analyses of RNA-seq Experiments Using Quasi-Likelihood Methods in edgeR. In Statistical Genomics: Methods and Protocols; Mathé, E., Davis, S., Eds.; Springer: New York, NY, USA, 2016; pp. 391–416. [Google Scholar]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Cline, M.S.; Smoot, M.; Cerami, E.; Kuchinsky, A.; Landys, N.; Workman, C.; Christmas, R.; Avila-Campilo, I.; Creech, M.; Gross, B.; et al. Integration of biological networks and gene expression data using Cytoscape. Nat. Protoc. 2007, 2, 2366–2382. [Google Scholar] [CrossRef] [Green Version]
- Kanoh, S.; Rubin, B.K. Mechanisms of Action and Clinical Application of Macrolides as Immunomodulatory Medications. Clin. Microbiol. Rev. 2010, 23, 590–615. [Google Scholar] [CrossRef] [Green Version]
- Yu, C.-C.; Chia-Ti, T.; Chen, P.-L.; Wu, C.-K.; Chiu, F.-C.; Chiang, F.-T.; Chen, P.-S.; Chen, C.-L.; Lin, L.-Y.; Juang, J.-M.; et al. KCNN2 polymorphisms and cardiac tachyarrhythmias. Medicine 2016, 95, e4312. [Google Scholar] [CrossRef]
- Gellens, M.E.; George, A.L.; Chen, L.Q.; Chahine, M.; Horn, R.; Barchi, R.L.; Kallen, R.G. Primary structure and functional expression of the human cardiac tetrodotoxin-insensitive voltage-dependent sodium channel. Proc. Natl. Acad. Sci. USA 1992, 89, 554–558. [Google Scholar] [CrossRef] [Green Version]
- Gabelli, S.B.; Boto, A.; Kuhns, V.H.; Bianchet, M.A.; Farinelli, F.; Aripirala, S.; Yoder, J.; Jakoncic, J.; Tomaselli, G.F.; Amzel, L.M. Regulation of the NaV1.5 cytoplasmic domain by calmodulin. Nat. Commun. 2014, 5, 5126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musa, H.; Kline, C.F.; Sturm, A.C.; Murphy, N.; Adelman, S.; Wang, C.; Yan, H.; Johnson, B.L.; Csepe, T.A.; Kilic, A.; et al. SCN5A variant that blocks fibroblast growth factor homologous factor regulation causes human arrhythmia. Proc. Natl. Acad. Sci. USA 2015, 112, 12528–12533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Meng, X.; Yuchi, Z.; Zhao, Z.; Xu, D.; Fedida, D.; Wang, Z.; Huang, C. De Novo Mutation in the SCN5A Gene Associated with Brugada Syndrome. Cell. Physiol. Biochem. 2015, 36, 2250–2262. [Google Scholar] [CrossRef] [Green Version]
- Gokhman, D.; Kelman, G.; Amartely, A.; Gershon, G.; Tsur, S.; Carmel, L. Gene ORGANizer: Linking genes to the organs they affect. Nucleic Acids Res. 2017, 45, W138–W145. [Google Scholar] [CrossRef] [Green Version]
- Stewart, S.E.; Fagerness, J.A.; Platko, J.; Smoller, J.W.; Scharf, J.M.; Illmann, C.; Jenike, E.; Chabane, N.; Leboyer, M.; Delorme, R.; et al. Association of theSLC1A1 glutamate transporter gene and obsessive-compulsive disorder. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2007, 144B, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Karan, K.; Satishchandra, P.; Sinha, S.; Anand, A. Rare SLC1A1 variants in hot water epilepsy. Qual. Life Res. 2017, 136, 693–703. [Google Scholar] [CrossRef]
- Giguère, S.; Lee, E.; Williams, E.; Cohen, N.; Chaffin, M.K.; Halbert, N.; Martens, R.J.; Franklin, R.P.; Clark, C.C.; Slovis, N.M. Determination of the prevalence of antimicrobial resistance to macrolide antimicrobials or rifampin inRhodococcus equiisolates and treatment outcome in foals infected with antimicrobial-resistant isolates ofR equi. J. Am. Veter. Med. Assoc. 2010, 237, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Huber, L.; Giguère, S.; Slovis, N.M.; Carter, C.N.; Barr, B.S.; Cohen, N.D.; Elam, J.; Erol, E.; Locke, S.J.; Phillips, E.D. Emergence of resistance to macrolides and rifampin in clinical isolates of Rhodococcus equi from foals in central Kentucky, 1995 to 2017. Antimicrob. Agents Chemother. 2019, 63, e01714-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popovic, D.; Vucic, D.; Dikic, I. Ubiquitination in disease pathogenesis and treatment. Nat. Med. 2014, 20, 1242–1253. [Google Scholar] [CrossRef]
- He, W.; Li, A.G.; Wang, D.; Han, S.; Zheng, B.; Goumans, M.; Dijke, P.T.; Wang, X. Overexpression of Smad7 results in severe pathological alterations in multiple epithelial tissues. EMBO J. 2002, 21, 2580–2590. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Li, A.G.; Liang, Y.-Y.; Owens, P.; He, W.; Lu, S.; Yoshimatsu, Y.; Wang, D.; Dijke, P.T.; Lin, X.; et al. Smad7-Induced β-Catenin Degradation Alters Epidermal Appendage Development. Dev. Cell 2006, 11, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Li, A.G.; Lu, S.-L.; Han, G.; Hoot, K.E.; Wang, X.-J. Role of TGFβ in skin inflammation and carcinogenesis. Mol. Carcinog. 2006, 45, 389–396. [Google Scholar] [CrossRef]
- Pruitt, K.D.; Brown, G.R.; Hiatt, S.M.; Thibaud-Nissen, F.; Astashyn, A.; Ermolaeva, O.; Farrell, C.M.; Hart, J.; Landrum, M.J.; McGarvey, K.M.; et al. RefSeq: An update on mammalian reference sequences. Nucleic Acids Res. 2014, 42, D756–D763. [Google Scholar] [CrossRef]
- Delamater, E.D.; Hunter, M.E.; Szybalski, W.; Bryson, V. Chemically Induced Aberrations of Mitosis in Bacteria. J. Gen. Microbiol. 1955, 12, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Martinez, A.O.; Norwood, T.H.; Martin, G.M. Growth inhibition and morphologic modulation of human fibroblastlike cells by erythromycin. In Vitro 1981, 17, 979–984. [Google Scholar] [CrossRef]
- Lastikka, L.; Virsu, M.L.; Halkka, O.; Eriksson, K.; Estola, T. Goniomitosis in rats affected by mycoplasma or macrolides. Med. Biol. 1976, 54, 146–149. [Google Scholar] [PubMed]
- Giguère, S.; Jacks, S.; Roberts, G.D.; Hernandez, J.; Long, M.T.; Ellis, C. Retrospective Comparison of Azithromycin, Clarithromycin, and Erythromycin for the Treatment of Foals with Rhodococcus equi Pneumonia. J. Veter. Intern. Med. 2004, 18, 568. [Google Scholar] [CrossRef]
- Båverud, V. Clostridium difficile infections in animals with special reference to the horse. A review. Vet. Q. 2002, 24, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.; Diab, S.; Blanchard, P.; Moore, J.; Anthenill, L.; Shahriar, F.; Garcia, J.; Songer, J. Clostridium perfringens type C and Clostridium difficile co-infection in foals. Veter. Microbiol. 2012, 156, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Periti, P.; Mazzei, T.; Mini, E.; Novelli, A.; Periti, P.; Periti, P.P. Adverse Effects of Macrolide Antibacterials. Drug Saf. 1993, 9, 346–364. [Google Scholar] [CrossRef]
- So, K.W.; Ng, P.C. Erythromycin and gastrointestinal dysmotility in preterm infants. East. J. Med. 2010, 15, 146. [Google Scholar]
- Armstrong, D.N.; Ballantyne, G.H.; Modlin, I.M. Erythromycin stimulates ileal motility by activation of dihydropyridine-sensitive calcium channels. J. Surg. Res. 1992, 52, 140–146. [Google Scholar] [CrossRef]
- Broad, J.; Sanger, G.J. The antibiotic azithromycin is a motilin receptor agonist in human stomach: Comparison with erythromycin. Br. J. Pharmacol. 2013, 168, 1859–1867. [Google Scholar] [CrossRef] [Green Version]
- Koh, S.D.; Ward, S.M.; Sanders, K.M. Ionic conductances regulating the excitability of colonic smooth muscles. Neurogastroenterol. Motil. 2012, 24, 705–718. [Google Scholar] [CrossRef]
- Kerr, M.G.; Snow, D.H. Composition of sweat of the horse during prolonged epinephrine (adrenaline) infusion, heat exposure, and exercise. Am. J. Veter. Res. 1983, 44, 1571–1577. [Google Scholar]
- Marlin, D.J.; Schroter, R.C.; Scott, C.M.; White, S.; Nyrop, K.A.; Maykuth, P.L.; Harris, P.A. Sweating and skin temperature responses of normal and anhidrotic horses to intravenous adrenaline. Equine Veter. J. 2010, 31, 362–369. [Google Scholar] [CrossRef]
- Tamaoki, J.; Takemura, H.; Tagaya, E.; Konno, K. Effect of Clarithromycin on Transepithelial Potential Difference in Rabbit Tracheal Mucosa. J. Infect. Chemother. 1995, 1, 112–115. [Google Scholar] [CrossRef]
- Afshari, P.; Yao, W.-D.; Middleton, F.A. Reduced Slc1a1 expression is associated with neuroinflammation and impaired sensorimotor gating and cognitive performance in mice: Implications for schizophrenia. PLoS ONE 2017, 12, e0183854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolter, J.; Seeney, S.; Bell, S.; Bowler, S.; Masel, P.; McCormack, J. Effect of long term treatment with azithromycin on disease parameters in cystic fibrosis: A randomised trial. Thorax 2002, 57, 212–216. [Google Scholar] [CrossRef] [Green Version]
- Equi, A.; Balfour-Lynn, I.; Bush, A.; Rosenthal, M. Long term azithromycin in children with cystic fibrosis: A randomised, placebo-controlled crossover trial. Lancet 2002, 360, 978–984. [Google Scholar] [CrossRef]
- Cigana, C.; Assael, B.M.; Melotti, P. Azithromycin Selectively Reduces Tumor Necrosis Factor Alpha Levels in Cystic Fibrosis Airway Epithelial Cells. Antimicrob. Agents Chemother. 2007, 51, 975–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelman, A.; Saussereau, E. [Cystic fibrosis and other channelopathies]. Arch Pediatr 2012, 19 (Suppl. 1), S13–S16. [Google Scholar] [CrossRef]
- Quinton, P.M. Chloride impermeability in cystic fibrosis. Nat. Cell Biol. 1983, 301, 421–422. [Google Scholar] [CrossRef]
- Collins, F.; Jurivich, D.; Sistonen, L.; Kroes, R.; Morimoto, R. Cystic fibrosis: Molecular biology and therapeutic implications. Science 1992, 256, 774–779. [Google Scholar] [CrossRef]
- Szczesniak, R.; Heltshe, S.L.; Stanojevic, S.; Mayer-Hamblett, N. Use of FEV1 in cystic fibrosis epidemiologic studies and clinical trials: A statistical perspective for the clinical researcher. J. Cyst. Fibros. 2017, 16, 318–326. [Google Scholar] [CrossRef]
- Tirouvanziam, R.; De Bentzmann, S.; Hubeau, C.; Hinnrasky, J.; Jacquot, J.; Péault, B.; Puchelle, E. Inflammation and Infection in Naive Human Cystic Fibrosis Airway Grafts. Am. J. Respir. Cell Mol. Biol. 2000, 23, 121–127. [Google Scholar] [CrossRef]
- Reemann, P.; Reimann, E.; Ilmjärv, S.; Porosaar, O.; Silm, H.; Jaks, V.; Vasar, E.; Kingo, K.; Kõks, S. Melanocytes in the skin--comparative whole transcriptome analysis of main skin cell types [published correction appears in PLoS One. 2017 Mar 7;12 (3):e0173792]. PLoS ONE 2014, 9, e115717. [Google Scholar] [CrossRef] [Green Version]
- Robertshaw, D. Sweat and heat-exchange in man and other mammals. J. Hum. Evol. 1985, 14, 63–73. [Google Scholar] [CrossRef]
- Evans, C.L.; Ross, K.A.; Smith, D.F.G.; Weilmalherbe, H. A physiological explanation of equine tropical anhidrosis. J. Physiol.-Lond. 1957, 135, P41–P42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkinson, D.M.; Elder, H.Y.; Bovell, D.L. Equine sweating and anhidrosis Part 2: Anhidrosis. Veter. Dermatol. 2007, 18, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Hammond, R.S.; Bond, C.T.; Strassmaier, T.; Ngo-Anh, T.J.; Adelman, J.P.; Maylie, J.; Stackman, R.W. Small-Conductance Ca2+-Activated K+ Channel Type 2 (SK2) Modulates Hippocampal Learning, Memory, and Synaptic Plasticity. J. Neurosci. 2006, 26, 1844–1853. [Google Scholar] [CrossRef] [Green Version]
- Ko, W.H.; O’Dowd, J.J.; Pediani, J.D.; Bovell, D.L.; Elder, H.Y.; Jenkinson, D.M.; Wilson, S.M. Extracellular ATP can activate autonomic signal transduction pathways in cultured equine sweat gland epithelial cells. J. Exp. Biol. 1994, 190, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Rosa, L.P.; Walker, N.; Mallicote, M.; MacKay, R.J.; Brooks, S.A. Genomic Association of Chronic Idiopathic Anhidrosis to a Potassium Channel Subunit in a Large Animal Model. J. Investig. Dermatol. 2021, 141, 2639–2645.e3. [Google Scholar] [CrossRef]
- Ray, W.A.; Murray, K.T.; Meredith, S.; Narasimhulu, S.S.; Hall, K.; Stein, C.M. Oral Erythromycin and the Risk of Sudden Death from Cardiac Causes. N. Engl. J. Med. 2004, 351, 1089–1096. [Google Scholar] [CrossRef]
- Wang, Q.; Li, Z.; Shen, J.; Keating, M.T. Genomic Organization of the HumanSCN5AGene Encoding the Cardiac Sodium Channel. Genomics 1996, 34, 9–16. [Google Scholar] [CrossRef]
- Zhang, T.; Yong, S.L.; Tian, X.-L.; Wang, Q.K. Cardiac-specific overexpression of SCN5A gene leads to shorter P wave duration and PR interval in transgenic mice. Biochem. Biophys. Res. Commun. 2007, 355, 444–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drenth, J.P.; Waxman, S.G. Mutations in sodium-channel gene SCN9A cause a spectrum of human genetic pain disorders. J. Clin. Investig. 2007, 117, 3603–3609. [Google Scholar] [CrossRef] [PubMed]
- Majeed, M.; Ubaidulhaq, M.; Rugnath, A.; Eriator, I. Extreme Ends of Pain Sensitivity in SCN9A Mutation Variants: Case Report and Literature Review. Innov. Clin. Neurosci. 2018, 15, 33–35. [Google Scholar] [PubMed]
- Tian, X.; Irannejad, R.; Bowman, S.L.; Du, Y.; Puthenveedu, M.A.; von Zastrow, M.; Benovic, J.L. The α-Arrestin ARRDC3 Regulates the Endosomal Residence Time and Intracellular Signaling of the β2-Adrenergic Receptor. J. Biol. Chem. 2016, 291, 14510–14525. [Google Scholar] [CrossRef] [Green Version]
- Mersmann, H.J. Overview of the effects of β-adrenergic receptor agonists on animal growth including mechanisms of action. J. Anim. Sci. 1998, 76, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.L.; Smith, D.F.G.; Weil-Malherbe, H. The relation between sweating and the catechol content of the blood in the horse. J. Physiol. 1956, 132, 542–552. [Google Scholar] [CrossRef] [Green Version]
- Evans, C.L.; Smith, D.F.G.; Ross, K.A.; Weil-Malherbe, H. Physiological factors in the condition of “dry coat” in horses. Vet. Rec. 1957, 69, 1–9. [Google Scholar]
- Bijman, J.; Quinton, P.M. Predominantly beta-adrenergic control of equine sweating. Am. J. Physiol. Integr. Comp. Physiol. 1984, 246, R349–R353. [Google Scholar] [CrossRef]
- Hislop, J.N.; von Zastrow, M. Role of ubiquitination in endocytic trafficking of G-protein-coupled receptors. Traffic 2011, 12, 137–148. [Google Scholar] [CrossRef]
- Qi, S.; O’Hayre, M.; Gutkind, J.S.; Hurley, J.H. Insights into β2-adrenergic receptor binding from structures of the N-terminal lobe of ARRDC3. Protein Sci. 2014, 23, 1708–1716. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.C.S.; Corbett, A.D.; Steel, C.; Pannirselvam, R.; Bovell, D.L. A preliminary study of the short circuit current (Isc) responses of sweat gland cells from normal and anhidrotic horses to purinergic and adrenergic agonists. Veter. Dermatol. 2007, 18, 152–160. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | logFC | logCPM | LR | p Value | |
|---|---|---|---|---|---|
| DOWNREGULATED | ENSECAT00000046497 | −7.38989 | 7.370481 | 38.36492 | 7.22 × 10−8 |
| MAP3K1 | −7.90642 | 6.997842 | 40.21128 | 1.69 × 10−7 | |
| ATP5F1E | −8.13404 | 6.513773 | 39.98864 | 1.79 × 10−7 | |
| ENSECAT00000003591 | −6.44825 | 6.857703 | 34.98519 | 2.01 × 10−7 | |
| CXXC1 | −6.81169 | 6.772696 | 34.79323 | 2.19 × 10−7 | |
| CTNNBL1 | −6.88469 | 6.306215 | 37.67177 | 5.04 × 10−7 | |
| ENSECAT00000004781 | −5.20358 | 5.421982 | 37.01026 | 5.81 × 10−7 | |
| BTAF1 | −6.27921 | 7.804846 | 31.50879 | 6.34 × 10−7 | |
| DRG1 | −6.94174 | 6.382133 | 32.17161 | 7.78 × 10−7 | |
| MEST | −7.69547 | 7.025976 | 34.08881 | 8.49 × 10−7 | |
| UPREGULATED | HAUS1 | 7.426906 | 5.73278 | 153.6852 | 1.68 × 10−15 |
| SMPD4 | 6.856443 | 6.812866 | 110.1973 | 2.86 × 10−14 | |
| ARRDC3 | 6.410874 | 6.280866 | 87.45059 | 9.47 × 10−12 | |
| PLCL1 | 6.732944 | 5.847365 | 78.57107 | 4.21 × 10−11 | |
| FBXO5 | 4.928078 | 6.33902 | 72.20562 | 1.32 × 10−10 | |
| SMAD7 | 5.714715 | 5.256898 | 79.97352 | 2.28 × 10−10 | |
| TRAK2 | 5.459412 | 5.220843 | 79.77119 | 2.35 × 10−10 | |
| KLF5 | 6.298051 | 6.073846 | 60.85152 | 4.25 × 10−10 | |
| REV1 | 4.162814 | 4.400905 | 64.81132 | 5.39 × 10−10 | |
| GABRB1 | 5.821451 | 5.342432 | 68.36093 | 1.41 × 10−9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patterson Rosa, L.; Mallicote, M.F.; MacKay, R.J.; Brooks, S.A. Ion Channel and Ubiquitin Differential Expression during Erythromycin-Induced Anhidrosis in Foals. Animals 2021, 11, 3379. https://doi.org/10.3390/ani11123379
Patterson Rosa L, Mallicote MF, MacKay RJ, Brooks SA. Ion Channel and Ubiquitin Differential Expression during Erythromycin-Induced Anhidrosis in Foals. Animals. 2021; 11(12):3379. https://doi.org/10.3390/ani11123379
Chicago/Turabian StylePatterson Rosa, Laura, Martha F. Mallicote, Robert J. MacKay, and Samantha A. Brooks. 2021. "Ion Channel and Ubiquitin Differential Expression during Erythromycin-Induced Anhidrosis in Foals" Animals 11, no. 12: 3379. https://doi.org/10.3390/ani11123379
APA StylePatterson Rosa, L., Mallicote, M. F., MacKay, R. J., & Brooks, S. A. (2021). Ion Channel and Ubiquitin Differential Expression during Erythromycin-Induced Anhidrosis in Foals. Animals, 11(12), 3379. https://doi.org/10.3390/ani11123379