
Prevalence Rate and Molecular Characteristics of Oestrus ovis L. (Diptera, Oestridae) in Sheep and Goats from Riyadh, Saudi Arabia

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Larvae Collection and Morphological Identification
2.2. DNA Extraction and Polymerase Chain Reaction (PCR)
2.3. Nucleotide Sequences of the mtCOI Gene
2.4. Statistical Analysis
3. Results
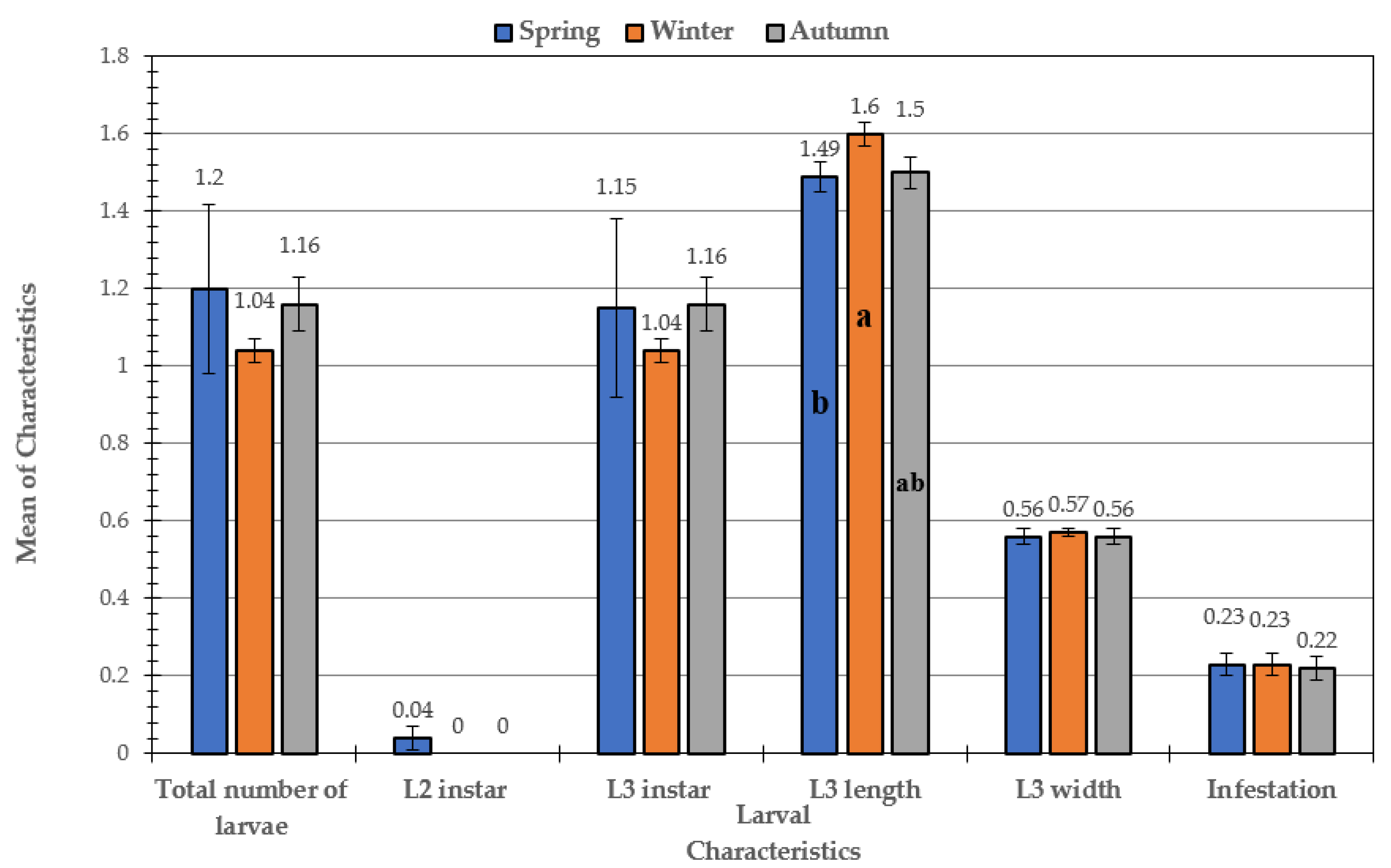
3.1. Prevalence of Infestation over the Course of the Study in Sheep and Goats
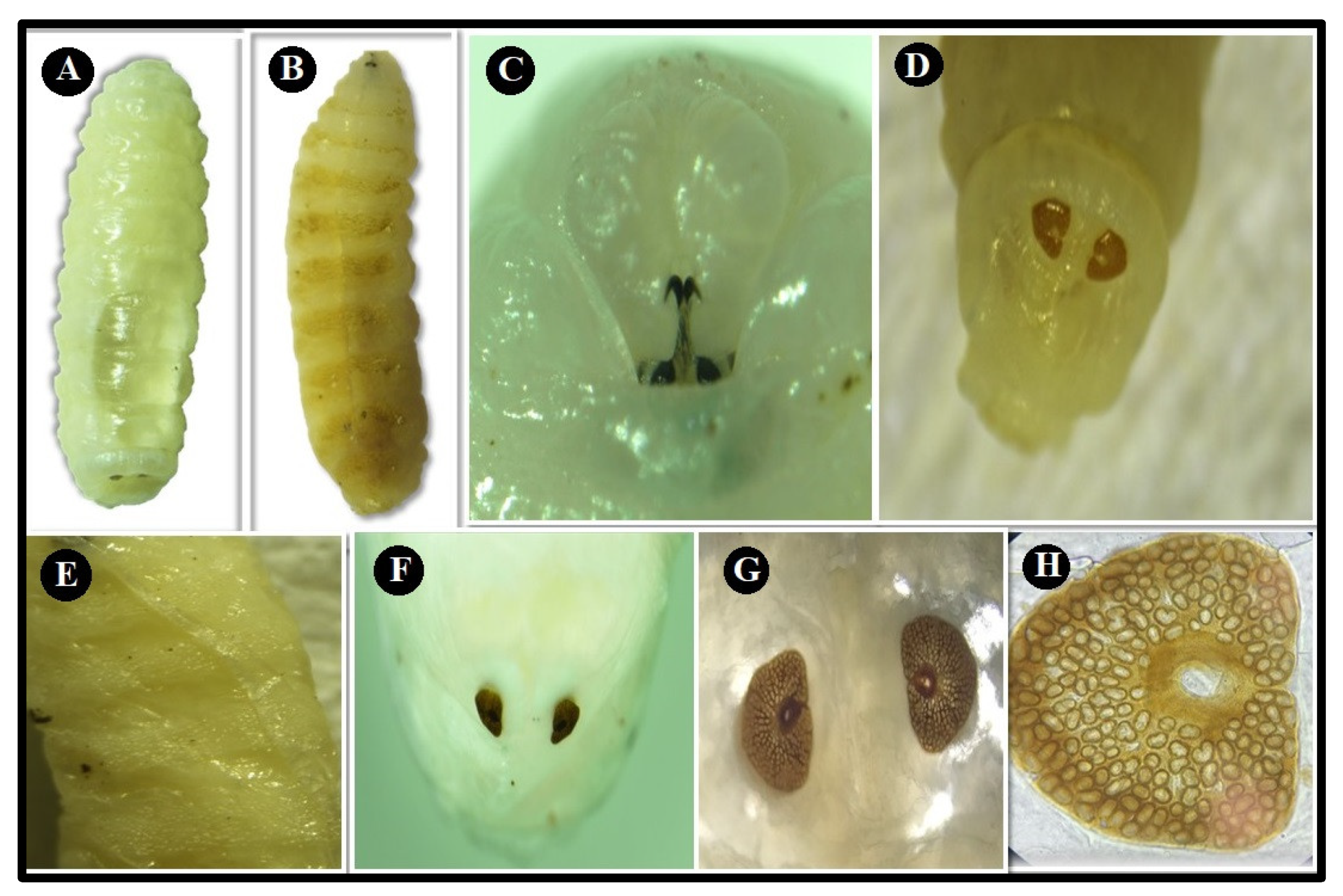
3.2. Morphological Examination
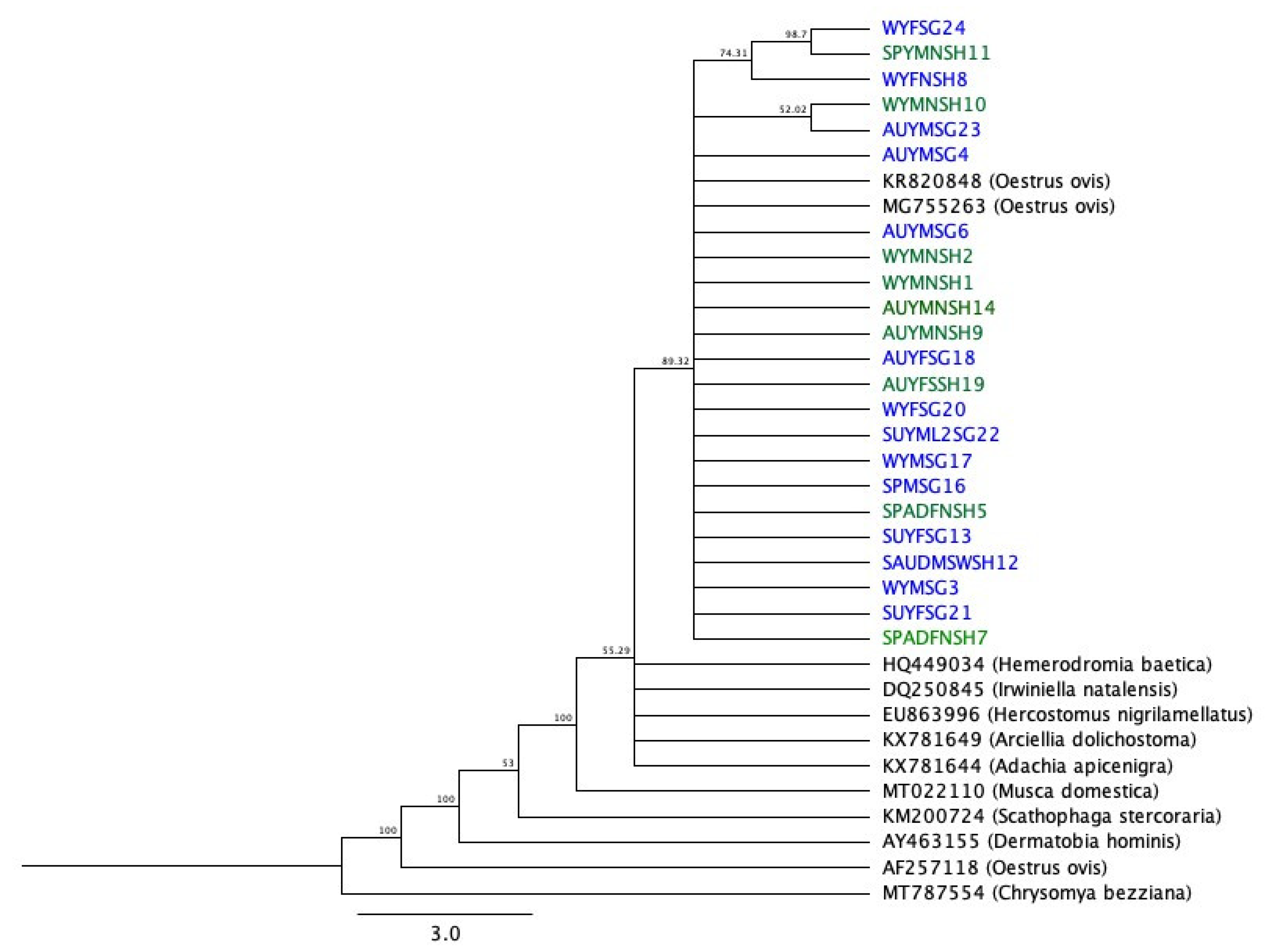
3.3. PCR Amplification and Nucleotide Sequence Analysis of the Partial mtCOI Gene
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hanan, B.A. Seasonal prevalence of Oestrus ovis L. (Diptera: Oestridae) larvae in infested sheep in Jazan Region. Saudi Arabia. J. Parasitol. Vector Biol. 2013, 5, 66–71. [Google Scholar]
- Traversa, D.; Otranto, D. A new approach for the diagnosis of myiasis of animals: The example of horse nasal myiasis. Vet. Parasitol. 2006, 141, 186–190. [Google Scholar] [CrossRef]
- Hidalgo, A.; Palma, H.; Oberg, C.; Fonseca-Salamanca, F. Oestrus ovis infestation of grazing sheep during summer in southern Chile. Pesqui. Veterinária Bras. 2015, 35, 497–500. [Google Scholar] [CrossRef] [Green Version]
- Ogo, N.I.; Onovoh, E.; Okubanjo, O.O.; Galindo, R.C.; De La Lastra, J.-M.P.; De La Fuente, J.; De La Lastra, J.M.P. Molecular identification of Cordylobia anthropophaga Blanchard (Diptera: Calliphoridae) larvae collected from dogs (Canis familiaris) in Jos South, Plateau State, Nigeria. Onderstepoort J. Vet. Res. 2012, 79, 4. [Google Scholar] [CrossRef] [Green Version]
- Otranto, D.; Traversa, D. Molecular evidence indicating that Przhevalskiana silenus, P. aegagri and P. crossii (Diptera, Oestridae) are one species. Acta Parasitol. 2004, 49, 173–176. [Google Scholar]
- Hall, M.; Adams, Z.; Wyatt, N.; Testa, J.; Edge, W.; Nikolausz, M.; Farkas, R.; Ready, P. Morphological and mitochondrial DNA characters for identification and phylogenetic analysis of the myiasis causing flesh fly Wohlfahrtia magnifica and its relatives, with a description of Wohlfahrtia monegrosensis sp. n. Wyatt & Hall. Med. Vet. Entomol. 2009, 23, 59–71. [Google Scholar]
- Ames, C.; Turner, B.; Daniel, B. The use of mitochondrial cytochrome oxidase I gene (COI) to differentiate two UK blowfly species—Calliphora vicina and Calliphora vomitoria. Forensic Sci. Int. 2006, 164, 179–182. [Google Scholar] [CrossRef]
- Otranto, D.; Stevens, J.R. Molecular approaches to the study of myiasis-causing larvae. Int. J. Parasitol. 2002, 32, 1345–1360. [Google Scholar] [CrossRef]
- Otranto, D.; Puccini, V. Cytochrome oxidase I (COI) gene of some obligate myiasis causing larvae (Diptera: Oestridae): Which perspective for phylogenetic studies? Preliminary data. Proc. Cost Action 2000, 833, 195–201. [Google Scholar]
- Otranto, D.; Colwell, D.D.; Milillo, P.; Di Marco, V.; Paradies, P.; Napoli, C.; Giannetto, S. Report in Europe of nasal myiasis by Rhinoestrus spp. in horses and donkeys: Seasonal patterns and taxonomical considerations. Vet. Parasitol. 2004, 122, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Otranto, D.; Colwell, D.D.; Traversa, D.; Stevens, J.R. Species identification of Hypoderma affecting domestic and wild ruminants by morphological and molecular characterization. Med. Vet. Entomol. 2003, 17, 316–325. [Google Scholar] [CrossRef]
- Otranto, D.; Traversa, D.; Guida, B.; Tarsitano, E.; Fiorente, P.; Stevens, J.R. Molecular characterization of the mitochondrial cytochrome oxidase I gene of Oestridae species causing obligate myiasis. Med. Vet. Entomol. 2003, 17, 307–315. [Google Scholar] [CrossRef]
- Otranto, D.; Milillo, P.; Traversa, D.; Colwell, D.D. Morphological variability and genetic identity in Rhinoestrus spp. causing horse nasal myiasis. Med. Vet. Entomol. 2005, 19, 96–100. [Google Scholar] [CrossRef]
- Li, X.; Cai, J.F.; Guo, Y.D.; Wu, K.L.; Wang, J.F.; Liu, Q.L.; Wang, X.H.; Chang, Y.F.; Yang, L.; Lan, L.M.; et al. The availability of 16S rRNA for the identification of forensically important flies (Diptera: Muscidae) in China. Trop. Biomed. 2010, 27, 155–166. [Google Scholar] [PubMed]
- Xinghua, W.; Jufeng, C.; Yadong, G.; Yufeng, C.; Kunlu, W.; Qinlai, L.; Jifeng, C.; Yunfeng, C.; Jiangfeng, W.; Yang, L.; et al. The availability of 16SrDNA gene for identifying forensically important blowflies in China. Rom. J. Leg. Med. 2010, 18, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Hendawy, S.H.; Allam, N.A.; Kandil, O.M.; Zayed, A.A.; Desouky AR, A.; Mahmoud, A. Partial COI and 16S rRNA genes sequences of Cephalopina titillator mitochondrial dna: Evidence for variation in evolutionary rates within myiasis causing species. Indian J. Anim. Res. 2012, 52, 105–110. [Google Scholar]
- Moreno, V.; Romero-Fernández, I.; Marchal, J.A.; Beltran, M.; Granados, J.E.; Habela, M.A.; Tamadon, A.; Rakhshandehroo, E.; Sarasa, M.; Pérez, J.M.; et al. Molecular characterization of bot flies, Oestrus spp., (Diptera, Oestridae), from domestic and wild Bovidae hosts. Vet. Parasitol. 2015, 212, 473–477. [Google Scholar] [CrossRef]
- Ferrar, P. A Guide to the Breeding Habits and Immature Stages of Diptera Cyclorrhapha; EJ Brill/Scandinavian Science Press: Copenhagen, Denmark, 1987; Volume 8, pp. 1–2. [Google Scholar]
- Smith, K.G. An Introduction to the Immature Stages of British Flies: Diptera Larvae, with Notes on Eggs, Puparia and Pupae; Royal Entomological Society: London, UK, 1989; Volume 10, p. 14. [Google Scholar]
- Zumpt, F. Myiasis in man and animals in the Old World. A textbook for physicians, veterinarians and zoologists. In Myiasis in Man and Animals in the Old World: A Textbook for Physicians, Veterinarians and Zoologists; Butterworth & Co. (Publishers) Ltd.: London, UK, 1965. [Google Scholar]
- Bosly, A.H. Molecular identification studies on Oestrus ovis L. (Diptera: Oestridae) larvae infested sheep in jazan region, Saudi Arabia. Indian J. Anim. Res. 2018, 52, 105–110. [Google Scholar]
- Drummond, A.J.; Ashton, B.; Buxton, S.; Cheung, M.; Cooper, A.; Duran, C.; Field, M.; Heled, J.; Kearse, M.; Markowitz, S.; et al. Geneious v5.3. Biomatters Ltd: Auckland, New Zealand. 2010. Available online: http://www.geneious.com (accessed on 28 January 2021).
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Alahmed, A.M. Seasonal infestation of Oestrus ovis larvae in sheep heads in central region of Saudi Arabia. J. Egypt. Soc. Parasitol. 2000, 30, 895–901. [Google Scholar]
- Alikhan, M.; Al Ghamdi, K.; Al Zahrani, F.S.; Khater, E.I.; Allam, A.M. Prevalence and Salient Morphological Features of Myiasis Causing Dipteran Flies in Jeddah, Saudi Arabia. Biosci. Biotechnol. Res. Asia 2018, 15, 101–109. [Google Scholar] [CrossRef]
- Papadopoulos, E.; Chaligiannis, I.; Morgan, E.R. Epidemiology of Oestrus ovis L. (Diptera: Oestridae) larvae in sheep and goats in Greece. Small Rumin. Res. 2010, 89, 51–56. [Google Scholar] [CrossRef]
- Karatepe, B.; Karatepe, M.; Güler, S. Epidemiology of Oestrus ovis L. infestation in sheep in Nigde province, Turkey. Rev. Médecine Vétérinaire 2014, 165, 225–230. [Google Scholar]
- Mohsen, M.N.; Raham, S.E.; Gasim, M.H. Oestrus ovis larval infestation among sheep and goats of green moun-tain areas in Libya. J. Adv. Vet. Anim. Res. 2015, 2, 382–387. [Google Scholar]
- Amin, A.R.; Morsy, T.A.; Shoukry, A.; Mazyad, S.A. Oestrid head maggots in slaughtered sheep in Cairo abattoir. J. Egypt. Soc. Parasitol. 1997, 27, 855–861. [Google Scholar] [PubMed]
- Alcaide, M.; Reina, D.; Sánchez, J.; Frontera, E.; Navarrete, I. Seasonal variations in the larval burden distribution of Oestrus ovis in sheep in the southwest of Spain. Vet. Parasitol. 2003, 118, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Caracappa, S.; Rilli, S.; Zanghi, P.; Di Marco, V.; Dorchies, P. Epidemiology of ovine oestrosis (Oestrus ovis Linné 1761, Diptera: Oestridæ) in Sicily. Vet. Parasitol. 2000, 92, 233–237. [Google Scholar] [CrossRef]
- Rogers, C.E.; Knapp, F.W. Bionomics of the Sheep Bot Fly, Oestrus Ovis. Environ. Entomol. 1973, 2, 11–23. [Google Scholar] [CrossRef]
- Arslan, M.O.; Kara, M.; Gicik, Y. Epidemiology of Oestrus ovis infestations in sheep in Kars province of north-eastern Turkey. Trop. Anim. Health Prod. 2009, 41, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, M.Y.; Khater, H.F.; Omer, S.F.; Rahman, A.A. Epidemiology of Oestrus ovis infesting Egyptian sheep. In Proceedings of the XX International Congress of Mediterranean Federation of Health and Production of Ruminants, Assiut, Egypt, 19–22 February 2013; pp. 19–22. [Google Scholar]
- Mohammed, R.G.; Josef, S.S.; Abed, K.J. Prevalence of Oestrus ovis Larvae in Slaughtered Sheep of Misan City, Iraq. Syst. Rev. Pharm. 2020, 11, 652–655. [Google Scholar]
- Yilma, J.; Dorchies, P. Epidemiology of Oestrus ovis in southwest France. Vet. Parasitol. 1991, 40, 315–323. [Google Scholar] [CrossRef]
- Bart, A.G.; Minár, J. Probability description of regulation on the level of population and individual in the host-parasite system using Oestrus ovis (Diptera: Oestridae) as an example. Folia Parasitol. 1992, 39, 75–83. [Google Scholar]
- Marinho, M.A.T.; Wolff, M.; Ramos-Pastrana, Y.; de Azeredo-Espin, A.M.L.; Amorim, D.D.S. The first phylo-genetic study of Mesembrinellidae (Diptera: Oestroidea) based on molecular data: Clades and congruence with morpholog-ical characters. Cladistics 2017, 33, 134–152. [Google Scholar] [CrossRef]
- Karademir, G.K.; Usluğ, S.; Okur, M.; Inci, A.; Yıldırım, A. Molecular Characterization and Phylogenetic Analyses of Oestrus ovis Larvae Causing Human Naso-pharyngeal Myiasis Based on CO1 Barcode Sequences. Turk. J. Parasitol. 2020, 44, 43–47. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Sequences | References |
|---|---|---|---|
| mtCOI | FFCOI | 5′-GGAGCATTAATYGGRGAYG-3′ | [21] |
| RHCO | 5′-TAAACTTCAGGGTGACCAAAAATCA-3′ |
| Species | No. of Head Examined | Infestation Rate (%) | Mean of Infestation | No. of Larvae (%) | Total Larvae (%) | ||
|---|---|---|---|---|---|---|---|
| L1 | L2 | L3 | |||||
| Sheep | 600 | 134 (22.3) | 1.78 ± 0.02 a | 0.0 (0.0) | 2.0 (1.5) | 149.0 (111.2) | 151.0 (112.7) |
| Goat | 800 | 356 (44.5) | 1.56 ± 0.02 b | 0.0 (0.0) | 6.0 (1.7) | 482.0 (135.4) | 486 (136.5) |
| Species | Sex | No. of Examined Animals | No. of Infested (%) |
|---|---|---|---|
| Sheep | Male | 300 | 98 (32.66) |
| Female | 300 | 36 (12) | |
| Goat | Male | 400 | 262 (65.5) |
| Female | 400 | 94 (32.5) | |
| Total | 1400 | 490 (35.00) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Metwally, D.M.; Albasyouni, S.A.; Barakat, I.A.H.; Al-Turaiki, I.M.; Almuhanna, A.M.; Bashir, M.A.; Baghdadi, H.B.; El-Khadragy, M.F.; Alajmi, R.A. Prevalence Rate and Molecular Characteristics of Oestrus ovis L. (Diptera, Oestridae) in Sheep and Goats from Riyadh, Saudi Arabia. Animals 2021, 11, 689. https://doi.org/10.3390/ani11030689
Metwally DM, Albasyouni SA, Barakat IAH, Al-Turaiki IM, Almuhanna AM, Bashir MA, Baghdadi HB, El-Khadragy MF, Alajmi RA. Prevalence Rate and Molecular Characteristics of Oestrus ovis L. (Diptera, Oestridae) in Sheep and Goats from Riyadh, Saudi Arabia. Animals. 2021; 11(3):689. https://doi.org/10.3390/ani11030689
Chicago/Turabian StyleMetwally, Dina M., Shurug A. Albasyouni, Ibrahim A.H. Barakat, Isra M. Al-Turaiki, Amal M. Almuhanna, Muhammad A. Bashir, Hanadi B. Baghdadi, Manal F. El-Khadragy, and Reem A. Alajmi. 2021. "Prevalence Rate and Molecular Characteristics of Oestrus ovis L. (Diptera, Oestridae) in Sheep and Goats from Riyadh, Saudi Arabia" Animals 11, no. 3: 689. https://doi.org/10.3390/ani11030689
APA StyleMetwally, D. M., Albasyouni, S. A., Barakat, I. A. H., Al-Turaiki, I. M., Almuhanna, A. M., Bashir, M. A., Baghdadi, H. B., El-Khadragy, M. F., & Alajmi, R. A. (2021). Prevalence Rate and Molecular Characteristics of Oestrus ovis L. (Diptera, Oestridae) in Sheep and Goats from Riyadh, Saudi Arabia. Animals, 11(3), 689. https://doi.org/10.3390/ani11030689