
Chromosome Abnormalities and Fertility in Domestic Bovids: A Review




Abstract
:Simple Summary
Abstract
1. Introduction
2. Numerical Chromosome Abnormalities
2.1. Autosomes
2.2. Sex Chromosomes
2.2.1. X Trisomy
2.2.2. X Monosomy
2.2.3. XXY Syndrome
2.3. Sex Reversal Syndrome
2.3.1. XY Sex Reversal
2.3.2. XX Sex Reversal
2.4. XX/XY Mosaicism (Free-Martinism)
2.5. Diploid-Triploid XX/XXY Mosaicism (Mixoploidy)
3. Structural Chromosome Abnormalities
3.1. Reciprocal Translocations
3.2. Robertsonian Translocations (rob)
3.3. Simple Translocation
3.4. Pericentric Inversion
3.5. Tandem Fusion (TAN)
3.6. Cytogenetically Detectable Deletions and Duplications
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BTA | Bos taurus chromosome, 2n = 60 |
| BBU | Bubalus bubalis chromosome, 2n = 50 |
| OAR | Ovis aries chromosome, 2n = 54 |
| CHI | Capra hircus chromosome, 2n = 60 |
| BIN | Bos indicus chromosome, 2n = 60 |
| FISH | fluorescence in situ hybridization |
| Fiber-FISH | extended chromatin fiber-FISH |
| CGH-array | comparative genomic hybridization array |
| DSD | disorder sex development |
References
- Gustavsson, I.; Rockborn, G. Chromosome abnormality in three cases of lymphatic leukemia in cattle. Nature 1964, 203, 990. [Google Scholar] [CrossRef]
- Gustavsson, I. Cytogenetics, distribution and phenotypic effects of a translocation in swedish cattle. Hereditas 2009, 63, 68–169. [Google Scholar] [CrossRef]
- Gustavsson, I. Distribution of the 1/29 translocation in the A.I. Bull population of Swedish Red and White cattle. Hereditas 1971, 69, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Dyrendahl, I.; Gustavsson, I. Sexual functions, semen characteristics and fertility of bulls carrying the 1/29 chromosome translocation. Hereditas 1979, 90, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Kanagawa, H.; Kawata, K.; Ishikawa, T.; Odajima, T.; Inoue, T. Chromosome studies on heterosexual twins in cattle. 3. Sex-chromosome chimerism (XX/XY) in bone marrow specimens. Jpn. J. Vet. Res. 1966, 14, 123–126. [Google Scholar]
- Mcfeely, R.; Hare, W.; Biggers, J.; Diggers, J. Chromosome Studies in 14 Cases of Intersex in Domestic Mammals. Cytogenet. Genome Res. 1967, 6, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Mcfeely, R.A. Chromosome abnormalities in early embryos of the pig. J. Reprod. Fertil. 1967, 13, 579–581. [Google Scholar] [CrossRef] [PubMed]
- Basrur, P.; Kosaka, S.; Kanagawa, H. Blood Cell Chimerism and Freemartinism in Heterosexual Bovine Quadruplets. J. Hered. 1970, 61, 15–18. [Google Scholar] [CrossRef]
- Fechheimer, N.S. A cytogenetic survey of young bulls in the U.S.A. Vet. Rec. 1973, 93, 535–536. [Google Scholar] [CrossRef] [PubMed]
- Sysa, P.; Bernacki, Z.; Kunska, A. Intersexuality in cattle—A case of male pseudohermaphroditismus with a 60.XY karotype. Vet. Rec. 1974, 94, 30–31. [Google Scholar] [CrossRef]
- Dunn, H.O.; Lein, D.H.; McEntee, K. Testicular hypoplasia in a Hereford bull with 61,XXY karyotype: The bovine counterpart of human Klinefelter’s syndrome. Cornell Vet. 1980, 70, 137–146. [Google Scholar] [PubMed]
- King, W. Chromosome variation in the embryos of domestic animals. Cytogenet. Genome Res. 2008, 120, 81–90. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Di Berardino, D. Tools of the trade: Diagnostics and research in domestic animal cytogenetics. J. Appl. Genet. 2008, 49, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Hayes, H. Chromosome painting with human chromosome-specific DNA libraries reveals the extent and distribution of conserved segments in bovine chromosomes. Cytogenet. Cell Genet. 1995, 71, 168–174. [Google Scholar] [CrossRef]
- Iannuzzi, A.; Pereira, J.; Iannuzzi, C.; Fu, B.; Ferguson-Smith, M. Pooling strategy and chromosome painting characterize a living zebroid for the first time. PLoS ONE 2017, 12, e0180158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lorenzi, L.; Rossi, E.; Gimelli, S.; Parma, P. De novo Reciprocal Translocation t(5;6)(q13;q34) in Cattle: Cytogenetic and Molecular Characterization. Cytogenet. Genome Res. 2013, 142, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Cribiu, E.P.; Di Berardino, D.; Di Meo, G.P.; Gallagher, D.S.; Hayes, H.; Iannuzzi, L.; Popescu, C.P.; Rubes, J.; Schmutz, S.; Stranzinger, G.; et al. International System for Chromosome Nomenclature of Domestic Bovids (ISCNDB 2000). Cytogenet. Genome Res. 2001, 95, 283–299. [Google Scholar] [CrossRef]
- Agerholm, J.S.; Christensen, K. Trisomy 22 in a calf. J. Vet. Med. Ser. A 1993, 40, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Christensen, K.; Juul, L. A Case of Trisomy 22 in a Live Hereford Calf. Acta Vet. Scand. 1999, 40, 85–88. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Meo, G.P.; Leifsson, P.S.; Eggen, A.; Christensen, K. A Case of Trisomy 28 in Cattle Revealed by both Banding and FISH-Mapping Techniques. Hereditas 2001, 134, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Dunn, H.O.; Johnson, R.H., Jr. A 61,XY Cell Line in a Calf with Extreme Brachygnathia. J. Dairy Sci. 1972, 55, 524–526. [Google Scholar] [CrossRef]
- Herzog, A.; Höhn, H.; Rieck, G. Survey of recent situation of chromosome pathology in different breeds of german cattle. Ann. Genet. Sel. Anim. 1977, 9, 471–491. [Google Scholar] [CrossRef] [PubMed]
- Herzog, A.; Hoehn, H. Uber zwei weitere Fälle von autosomaler Trisomie, 61,XY, + 12 und 61,XX, + 12, beim Rind [Two ad-ditional cases of autosomal trisomy, 61,XY, + 12 and 61,XX, + 12, in cattle]. Cytogenet. Cell Genet. 1991, 57, 211–213. [Google Scholar] [CrossRef] [PubMed]
- Kovács, A.; Foote, R.H.; Lein, D.H. 1;16 tandem translocation with trisomy 16 in a Brown Swiss bull. Vet. Rec. 1990, 127, 205. [Google Scholar] [PubMed]
- Tschudi, P.; Ueltschi, G.; Martig, J.; Küpfer, U. Autosomale Trisomie als Ursache eines hohen Ventrikelseptumdefekts bei einem Kalb der Simmentalerrasse [Autosomal trisomy as the cause of a high ventricular septal defect in a calf of the Simmental breed]. Schweiz. Arch. Tierheilkd. 1975, 117, 335–340. [Google Scholar] [PubMed]
- Kulikova, S.G.; Petukhov, V.L.; Grafodatskiĭ, A.S. Novyĭ sluchaĭ trisomii u krupnogo rogatogo skota [A new case of trisomy in cattle]. Tsitol. Genet. 1991, 25, 28–31. [Google Scholar] [PubMed]
- Murakami, R.; Miyake, Y.; Kaneda, Y. Cases of XY female, single-birth freemartin and trisomy (61, XX, +20) observed in cy-togenetical studies on 18 sterile heifers. Nihon Juigaku Zasshi 1989, 51, 941–945. [Google Scholar] [CrossRef]
- Lioi, M.B.; Scarfi, M.R.; Di Berardino, D. An autosomal trisomy in cattle. Genet. Sel. Evol. 1995, 27, 473–476. [Google Scholar] [CrossRef]
- Gallagher, D.S., Jr.; Lewis, B.C.; De Donato, M.; Davis, S.K.; Taylor, J.F.; Edwards, J.F. Autosomal trisomy 20 (61,XX, + 20) in a malformed bovine fetus. Vet. Pathol. 1999, 36, 448–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Häfliger, I.M.; Agerholm, J.S.; Drögemüller, C. Constitutional trisomy 20 in an aborted Holstein fetus with pulmonary hy-poplasia and anasarca syndrome. Anim. Genet. 2020, 51, 988–989. [Google Scholar] [CrossRef] [PubMed]
- Long, S.E. Autosomal trisomy in a calf. Vet. Rec. 1984, 115, 16–17. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, S.M.; Moker, J.S.; Clark, E.G.; Orr, J.P. Chromosomal Aneuploidy Associated with Spontaneous Abortions and Neonatal Losses in Cattle. J. Vet. Diagn. Investig. 1996, 8, 91–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayr, B.; Krutzler, H.; Auer, H.; Schleger, W.; Sasshofer, K.; Glawischnig, E. A viable calf with trisomy 22. Cytogenet. Genome Res. 1985, 39, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Mayr, B.; Schellander, K.; Auer, H.; Tesarik, E.; Schleger, W.; Sasshofer, K.; Glawischnig, E. Offspring of a trisomic cow. Cytogenet. Genome Res. 1987, 44, 229–230. [Google Scholar] [CrossRef] [PubMed]
- Coates, J.; Rousseaux, C.; Schmutz, S. Multiple defects in an aborted bovine foetus associated with chromosomal trisomy. N. Z. Vet. J. 1987, 35, 173–174. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, S.M.; Coates, J.W.; Rousseaux, C.G. Chromosomal Trisomy in an Anormalous Bovine Fetus. Can. Vet. J. 1987, 28, 61–62. [Google Scholar] [PubMed]
- Mäkinen, A.; Alitalo, I.; Alanko, M. Autosomal Trisomy in a Heifer. Acta Vet. Scand. 1987, 28, 1–8. [Google Scholar] [CrossRef]
- Ducos, A.; Seguela, A.; Pinton, A.; Berland, H.; Brun-Baronnat, C.; Darre, R.; Manesse, M.; Darre, A. Trisomy 26 mosaicism in a sterile Holstein-Friesian heifer. Vet. Rec. 2000, 146, 163–164. [Google Scholar] [CrossRef]
- Iannuzzi, A.; Genualdo, V.; Perucatti, A.; Pauciullo, A.; Varricchio, G.; Incarnato, D.; Matassino, D.; Iannuzzi, L. Fatal Outcome in a Newborn Calf Associated with Partial Trisomy 25q and Partial Monosomy 11q, 60,XX,der(11)t(11;25)(q11;q14∼21). Cytogenet. Genome Res. 2015, 146, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Häfliger, I.M.; Seefried, F.; Drögemüller, C. Trisomy 29 in a stillborn Swiss Original Braunvieh calf. Anim. Genet. 2020, 51, 483–484. [Google Scholar] [CrossRef] [PubMed]
- Perucatti, A.; Genualdo, V.; Iannuzzi, A.; De Lorenzi, L.; Matassino, D.; Parma, P.; Di Berardino, D.; Di Meo, G. A New and Unusual Reciprocal Translocation in Cattle: Rcp(11;25)(q11;q14–21). Cytogenet. Genome Res. 2011, 134, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, A.; Braun, M.; Genualdo, V.; Perucatti, A.; Reinartz, S.; Proios, I.; Heppelmann, M.; Rehage, J.; Hülskötter, K.; Beineke, A.; et al. Clinical, cytogenetic and molecular genetic characterization of a tandem fusion translocation in a male Holstein cattle with congenital hypospadias and a ventricular septal defect. PLoS ONE 2020, 15, e0227117. [Google Scholar] [CrossRef] [PubMed]
- Hyttel, P.; Viuff, D.; Laurincik, J.; Schmidt, M.; Thomsen, P.; Avery, B.; Callesen, H.; Rath, D.; Niemann, H.; Rosenkranz, H.; et al. Risks of in-vitro production of cattle and swine embryos: Aberrations in chromosome numbers, ribosomal RNA gene activation and perinatal physiology. Hum. Reprod. 2000, 15, 87–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyon, M.F. Gene Action in the X-chromosome of the Mouse (Mus musculus L.). Nat. Cell Biol. 1961, 190, 372–373. [Google Scholar] [CrossRef]
- Burgoyne, P.S.; Ojarikre, O.A.; Turner, J.M.A. Evidence that postnatal growth retardation in XO mice is due to haploinsufficiency for a non-PAR X gene. Cytogenet. Genome Res. 2002, 99, 252–256. [Google Scholar] [CrossRef]
- Iannuzzi, L.; King, W.; Di Berardino, D. Chromosome Evolution in Domestic Bovids as Revealed by Chromosome Banding and FISH-Mapping Techniques. Cytogenet. Genome Res. 2009, 126, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Rieck, G.W.; Höhn, H.; Herzog, A. X-Trisomie beim Rind mit Anzeichen familiärer Disposition für Meiosestörungen [X-trisomy in cattle with signs of familial disposition for meiotic disturbances]. Cytogenetics 1970, 9, 401–409. (In German) [Google Scholar] [CrossRef] [PubMed]
- Norberg, H.S.; Refsdal, A.O.; Garm, O.N.; Nes, N. A case report on X-trisomy in cattle. Hereditas 2009, 82, 69–72. [Google Scholar] [CrossRef]
- Buoen, L.C.; Seguin, B.E.; Weber, A.F.; Shoffner, R.N. X-trisomy karyotype and associated infertility in a Holstein heifer. J. Am. Vet. Med. Assoc. 1981, 179, 808–811. [Google Scholar]
- Moreno-Millan, M.; Bermejo, J.V.D.; Garcia, F.A. X-trisomy in Friesian cow with continuous oestrus. Vet. Rec. 1987, 121, 167–168. [Google Scholar] [CrossRef]
- Pinheiro, L.; Almeida, I.; Garcia, J.; Basrur, P. Trisomy X and translocation in infertile heifers. Theriogenology 1987, 28, 891–898. [Google Scholar] [CrossRef]
- Citek, J.; Rubeš, J.; Hájková, J. Short communication: Robertsonian translocations, chimerism, and aneuploidy in cattle. J. Dairy Sci. 2009, 92, 3481–3483. [Google Scholar] [CrossRef] [PubMed]
- Yadav, B.R.; Balakrishnan, C.R. Trisomy of the X chromosome in a Murrah buffalo. Vet. Rec. 1982, 111, 184–185. [Google Scholar] [CrossRef] [PubMed]
- Prakash, B.; Balain, D.S.; Lathwal, S.S.; Malik, R.K. Trisomy-X in a sterile river buffalo. Vet. Rec. 1994, 134, 241–242. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Incarnato, D.; Palo, R.D.; Zicarelli, L. Reproductive disturbances and sex chromosome abnormalities in two female river buffaloes. Vet. Rec. 2004, 154, 823–824. [Google Scholar] [CrossRef]
- Powell, C. Sex chromosomes and sex chromosome abnormalities. In The Principles of Clinical Cyto-Genetics; Gersen, S., Keagle, M., Eds.; Humana Press: Totowa, NJ, USA, 1999; pp. 229–258. [Google Scholar]
- Prakash, B.; Balain, D.S.; Lathwal, S.S.; Malik, R.K. Infertility associated with monosomy-X in a crossbred cattle heifer. Vet. Rec. 1995, 137, 436–437. [Google Scholar] [CrossRef] [PubMed]
- Romano, J.E.; Raussdepp, T.; Mulon, P.Y.; Villadóniga, G.B. Non-mosaic monosomy 59,X in cattle: A case report. Anim. Reprod. Sci. 2015, 156, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.P.; Wolfe, A.; O’Donovan, J.; Byrne, N.; Sayers, R.G.; Dodds, K.G.; McEwan, J.C.; O’Connor, R.E.; McClure, M.; Purfield, D.C. Characterization of an X-chromosomal non-mosaic monosomy (59, X0) dairy heifer detected using routinely available single nucleotide polymorphism genotype data. J. Anim. Sci. 2017, 95, 1042–1049. [Google Scholar]
- Pinheiro, L.E.L.; Mikich, A.B.; Bechara, G.H.; Almeida, I.L.; Basrur, P.K. Isochromosome Y in an infertile heifer. Genome 1990, 33, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Yadav, B.R.; Kumar, P.; Tomer, O.S.; Kumar, S.; Balain, D.S. Monosomy X and gonadal dysgenesis in a buffalo heifer (Bubalus bubalis). Theriogenology 1990, 34, 99–105. [Google Scholar] [CrossRef]
- Prakash, B.; Balain, D.S.; Lathwal, S.S. A 49, XO sterile murrah buffalo (Bubalus bubalis). Vet. Rec. 1992, 130, 559–560. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Zicarelli, L. Sex chromosome monosomy (2n = 49,X) in a river buffalo (Bubalus bu-balis). Vet. Rec. 2000, 147, 690–691. [Google Scholar]
- Zartman, D.L.; Hinesley, L.L.; Gnatkowski, M.W. A 53, X female sheep (Ovis aries). Cytogenet. Genome Res. 1981, 30, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Baylis, M.; Wayte, D.; Owen, J. An XO/XX mosaic sheep with associated gonadal dysgenesis. Res. Vet. Sci. 1984, 36, 125–126. [Google Scholar] [CrossRef]
- Berry, D.P.; O’Brien, A.; O’Donovan, J.; McHugh, N.; Wall, E.; Randles, S.; McDermott, K.; O’Connor, R.E.; Patil, M.A.; Ho, J.; et al. Aneuploidy in dizygotic twin sheep detected using genome-wide single nucleotide polymorphism data from two commonly used commercial vendors. Animal 2018, 12, 2462–2469. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, S.; Shanker, V. A case report on XO/XX/XXX mixoploidy in a goat. Vet. Rec. 1990, 126, 312–313. [Google Scholar] [PubMed]
- Switonski, M.; Szczerbal, I. Chromosome Abnormalities in Domestic Animals as Causes of Disorders of Sex Development or Impaired Fertility. In Insights from Animal Reproduction; Intech Open Science: London, UK, 2016. [Google Scholar]
- Rieck, G.W.; Höhn, H.; Herzog, A. Hypogonadismus, intermittierender Kryptorchismus und segmentäre Aplasie der Ductus Wolffii bei einem männlichen Rind mit XXY--Gonosomen-Konstellation bzw. XXY--XX--XY-Gonosomen-Mosaik [Hy-pogonadism, intermittent cryptorchism and segmentary aplasia of the Wolffian duct in a bull with XXY gonosome constel-lation or XXY--XX--XY gonosome mosaic]. Dtsch. Tierarztl. Wochenschr. 1969, 76, 133–138. [Google Scholar]
- Rieck, G.W.; Höhn, H.; Schmidt, I. Vulvaaplasie und Urethra masculina: Maskulinisierungseffekte des Sinus urogenitalis bei genetisch weiblichen Rindern durch Chimärismen mit dem XXY-Gonosomen-Komplement [Aplastic vulva and masculine urethra: Masculinization effects of the urogenital sinus in genetically female cattle due to chimerism with the XXY gonosomal complement]. Berl. Munch. Tierarztl. Wochenschr. 1982, 95, 181–185. [Google Scholar]
- Dain, A.R.; Bridge, P.S. A chimaeric calf with XY/XXY mosaicism and intersexuality. J. Reprod. Fertil. 1978, 54, 197–201. [Google Scholar] [CrossRef] [Green Version]
- Logue, D.N.; Harvey, M.J.; Munro, C.D.; Lennox, B. Hormonal and histological studies in a 61XXY bull. Vet. Rec. 1979, 104, 500–503. [Google Scholar] [CrossRef]
- Miyake, Y.; Ishikawa, T.; Kanagawa, H.; Sato, K. A first case of XY/XYY mosaic bull. Jpn. J. Vet. Res. 1981, 29, 94–96. [Google Scholar] [PubMed]
- Alam, M.G.S.; Hurtado, R. Testicular hypoplasia syndrome due to chromosomal aberration (Trisomy 61/XXY). Indian Vet. J. 1982, 2, 55–60. [Google Scholar]
- Schmutz, S.M.; Barth, A.D.; Moker, J.S. A Klinefelter bull with a 1;29 translocation born to a fertile 61,XXX cow. Can. Vet. J. 1994, 35, 182–184. [Google Scholar]
- Molteni, L.; Macchi, A.D.G.; Meggiolaro, D.; Sironi, G.; Enice, F.; Popescu, P. New cases of XXY constitution in cattle. Anim. Reprod. Sci. 1999, 55, 107–113. [Google Scholar] [CrossRef]
- Joerg, H.; Janett, F.; Schlatt, S.; Mueller, S.; Graphodatskaya, D.; Suwattana, D.; Asai, M.; Stranzinger, G. Germ cell transplan-tation in an azoospermic Klinefelter bull. Biol. Reprod. 2003, 69, 1940–4194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaszczak, K.; Parada, R.; Wardecka, B.; Niemczewski, C. A note on analysis of chromosome constitution in tissues, quality of semen and DNA microsatellite loci in bull with 60,XY/61,XXY karyotype. J. Anim. Feed. Sci. 2003, 12, 521–527. [Google Scholar] [CrossRef] [Green Version]
- Słota, E.; Kozubska-Sobocińska, A.; Kościelny, M.; Danielak-Czech, B.; Rejduch, B. Detection of the XXY trisomy in a bull by using sex chromosome painting probes. J. Appl. Genet. 2003, 44, 379–382. [Google Scholar]
- Ducos, A.; Revay, T.; Kovacs, A.; Hidas, A.; Pinton, A.; Bonnet-Garnier, A.; Molteni, L.; Slota, E.; Switonski, M.; Arruga, M.V.; et al. Cytogenetic screening of livestock populations in Europe: An overview. Cytogenet. Genome Res. 2008, 120, 26–41. [Google Scholar] [CrossRef]
- Patel, R.; Singh, K.; Soni, K.; Chauhan, J. Novel cytogenetic finding: An unusual X;X-translocation in Mehsana buffalo (Bubalus bubalis). Cytogenet. Genome Res. 2006, 115, 186–188. [Google Scholar] [CrossRef]
- Bruere, A.N.; Marshall, R.B.; Ward, D.P.J. Testicular hypoplasia and xxy sex chromosome complement in two rams: The ovine counterpart of klinefelter’s syndrome in man. J. Reprod. Fertil. 1969, 19, 103–108. [Google Scholar] [CrossRef] [Green Version]
- Moraes, J.C.; Mattevi, M.S.; Ferreira, J.M. Chromosome studies in Brazilian rams. Vet. Rec. 1980, 107, 489–490. [Google Scholar] [CrossRef]
- Takebayashi, S.G.; Jorg, W. Testicular hypoplasia in a horned goat with 61, XXY/60,XY karyotype. Jpn. J. Genet. 1986, 61, 177–181. [Google Scholar] [CrossRef]
- Bhatia, S.; Shanker, V. First report of a XX/XXY fertile goat buck. Vet. Rec. 1992, 130, 271–272. [Google Scholar] [CrossRef] [PubMed]
- Henricson, B.; Åkesson, A. Two Heifers with Gonadal Dysgenesis and the Sex Chromosomal Constitution XY. Acta Vet. Scand. 1967, 8, 262–272. [Google Scholar] [CrossRef]
- Chapman, H.; Bruère, A.; Jaine, P. XY gonadal dysgenesis in a Charolais heifer. Anim. Reprod. Sci. 1978, 1, 9–18. [Google Scholar] [CrossRef]
- Sharma, A.K.; Vijaykumar, N.K.; Khar, S.K.; Verma, S.K.; Nigam, J.M. XY gonadal dysgenesis in a heifer. Vet. Rec. 1980, 107, 328–330. [Google Scholar] [CrossRef]
- Macmillan, K.L.; Fielden, E.D.; McNatty, K.P.; Henderson, H.V. LH concentrations in two cattle with XY gonadal dysgenesis. J. Reprod. Fertil. 1984, 71, 525–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondoh, S.; Miyake, Y.; Nakahori, Y.; Nakagome, Y.; Kaneda, Y. Cytogenetical and molecular biological studies on a bovine XY female. J. Vet. Med. Sci. 1992, 54, 1077–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hare, J.E.; Baird, J.D.; Duignan, P.; Saunders, J.; Floetenmeyer, R.; Basrur, P.K. XY gonadal dysgenesis and tetralogy of Fallot in an Angus calf. Can. Vet. J. 1994, 35, 510–512. [Google Scholar]
- Kawakura, K.; Miyake, Y.I.; Murakami, R.K.; Kondoh, S.; Hirata, T.I.; Kaneda, Y. Deletion of the SRY region on the Y chro-mosome detected in bovine gonadal hypoplasia (XY female) by PCR. Cytogenet. Cell Genet. 1996, 72, 183–184. [Google Scholar] [CrossRef] [PubMed]
- Kawakura, K.; Miyake, Y.-I.; Murakami, R.; Kondoh, S.; Hirata, T.-I.; Kaneda, Y. Abnormal structure of the Y chromosome detected in bovine gonadal hypoplasia (XY female) by FISH. Cytogenet. Genome Res. 1997, 76, 36–38. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, M.; Sorensen, A.M., Jr. Some cytogenetic aspects of intersexuality in the bovine. J. Anim. Sci. 1971, 32, 1219–1228. [Google Scholar] [CrossRef]
- Nicolae, I.; Popescu, C.P. Cytogenetic studies on Romanian cattle breeds. Arch. Zootec. 2001, 50, 355–361. [Google Scholar]
- Iannuzzi, L.; Di Meo, G.P.; Perucatti, A.; Di Palo, R.; Zicarelli, L. 50,XY gonadal dysgenesis (Swyer’s syndrome) in a female river buffalo (Bubalus bubalis). Vet. Rec. 2001, 148, 634–635. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, L.; Monteagudo, L.; De Jalon, J.G.; Tejedor, M.; Ramos, J.; Lacasta, D. A Case of Ovine Female XY Sex Reversal Syndrome Not Related to Anomalies in the Sex-Determining Region Y (SRY). Cytogenet. Genome Res. 2009, 126, 329–332. [Google Scholar] [CrossRef]
- Albarella, S.; D’Anza, E.; Galdiero, G.; Esposito, L.; De Biase, D.; Paciello, O.; Ciotola, F.; Peretti, V. Cytogenetic Analyses in Ewes with Congenital Abnormalities of the Genital Apparatus. Animals 2019, 9, 776. [Google Scholar] [CrossRef] [Green Version]
- Sulimovici, S.; Weissenberg, R.; Lunenfeld, B.; Padeh, B.; Soller, M. Testicular testosterone biosynthesis in male Saanen goats with XX sex chromosomes. Clin. Genet. 2008, 13, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Pailhoux, E.; Cribiu, E.P.; Chaffaux, S.; Darre, R.; Fellous, M.; Cotinot, C. Molecular analysis of 60,XX pseudohermaphrodite polled goats for the presence of SRY and ZFY genes. J. Reprod. Fertil. 1994, 100, 491–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Just, W.; Almeida, C.C.; Goldshmidt, B.; Vogel, W. The Male Pseudohermaphrodite XX Polled Goat is Zfy and Sry Negative. Hereditas 2004, 120, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Vaiman, D.; Koutita, O.; Oustry, A.; Elsen, J.-M.; Manfredi, E.; Fellous, M.; Cribiu, E.P. Genetic mapping of the autosomal region involved in XX sex-reversal and horn development in goats. Mamm. Genome 1996, 7, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Payen, E.J.; Cotinot, C.Y. Sequence evolution of SRY gene within Bovidae family. Mamm. Genome 1994, 5, 723–725. [Google Scholar] [CrossRef]
- Biason-Lauber, A.; Konrad, D.; Meyer, M.; Debeaufort, C.; Schoenle, E.J. Ovaries and Female Phenotype in a Girl with 46,XY Karyotype and Mutations in the CBX2 Gene. Am. J. Hum. Genet. 2009, 84, 658–663. [Google Scholar] [CrossRef] [Green Version]
- Pailhoux, E.; Vigier, B.; Vaiman, D.; Schibler, L.; Vaiman, A.; Cribiu, E.; Nezer, C.; Georges, M.; Sundström, J.; Pelliniemi, L.J.; et al. Contribution of domestic animals to the identification of new genes involved in sex determination. J. Exp. Zoo. 2001, 290, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Pepene, C.E.; Coman, I.; Mihu, D.; Militaru, M.; Duncea, I. Infertility in a new 46, XX male with positive SRY confirmed by fluorescence in situ hybridization: A case report. Clin. Exp. Obstet. Gynecol. 2008, 35, 299–300. [Google Scholar]
- Nebesio, T.D.; Torres-Martinez, W.; Rink, R.C.; Eugster, E.A. Spurious Case of XX Maleness in A Patient With A History of Wiskott-Aldrich Syndrome. Endocr. Pr. 2011, 17, e1–e3. [Google Scholar] [CrossRef]
- Di Meo, G.P.; Perucatti, A.; Floriot, S.; Incarnato, D.; Rullo, R.; Jambrenghi, A.C.; Ferretti, L.; Vonghia, G.; Cribiu, E.; Eggen, A.; et al. Chromosome evolution and improved cytogenetic maps of the Y chromosome in cattle, zebu, river buffalo, sheep and goat. Chromosom. Res. 2005, 13, 349–355. [Google Scholar] [CrossRef]
- Das, P.; Chowdhary, B.; Raudsepp, T. Characterization of the Bovine Pseudoautosomal Region and Comparison with Sheep, Goat, and Other Mammalian Pseudoautosomal Regions. Cytogenet. Genome Res. 2009, 126, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Raudsepp, T.; Das, P.; Avila, F.; Chowdhary, B. The Pseudoautosomal Region and Sex Chromosome Aneuploidies in Domestic Species. Sex. Dev. 2012, 6, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Villagómez, D.; Parma, P.; Radi, O.; Di Meo, G.; Pinton, A.; Iannuzzi, L.; King, W. Classical and Molecular Cytogenetics of Disorders of Sex Development in Domestic Animals. Cytogenet. Genome Res. 2009, 126, 110–131. [Google Scholar] [CrossRef]
- Favetta, L.; Villagómez, D.; Iannuzzi, L.; Di Meo, G.; Webb, A.; Crain, S.; King, W. Disorders of Sexual Development and Abnormal Early Development in Domestic Food-Producing Mammals: The Role of Chromosome Abnormalities, Environment and Stress Factors. Sex. Dev. 2012, 6, 18–32. [Google Scholar] [CrossRef]
- Parma, P.; Veyrunes, F.; Pailhoux, E. Sex Reversal in Non-Human Placental Mammals. Sex. Dev. 2016, 10, 326–344. [Google Scholar] [CrossRef]
- Padula, A. The freemartin syndrome: An update. Anim. Reprod. Sci. 2005, 87, 93–109. [Google Scholar] [CrossRef] [PubMed]
- Komisarek, J.; Dorynek, Z. Genetic aspects of twinning in cattle. J. Appl. Genet. 2002, 43, 55–68. [Google Scholar]
- Greene, W.; Dunn, H.; Foote, R. Sex-chromosome ratios in cattle and their relationship to reproductive development in freemartins. Cytogenet. Genome Res. 1977, 18, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, G.; Perucatti, A.; Di Palo, R.; Iannuzzi, A.; Ciotola, F.; Peretti, V.; Neglia, G.; Campanile, G.; Zicarelli, L. Sex chromosome abnormalities and sterility in river buffalo. Cytogenet. Genome Res. 2008, 120, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Ruvinsky, A.; Spicer, L.J. Developmental genetics: Sex determination and differentiation. In The Genetics of Cattle; Fries, R., Ruvinsky, A., Eds.; CARI Puhlishing and CAB International: Wallingford, UK, 1999; pp. 456–461. [Google Scholar]
- Cabianca, G.; Rota, A.; Cozzi, B.; Ballarin, C. Expression of AMH in Female Fetal Intersex Gonads in the Bovine. Anat. Histol. Embryol. 2006, 36, 24–26. [Google Scholar] [CrossRef] [PubMed]
- Dunn, H.O.; McEntee, K.; Hall, C.E.; Johnson, R.H., Jr.; Stone, W.H. Cytogenetic and reproductive studies of bulls born co-twin with freemartins. J. Reprod. Fertil. 1979, 57, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustavsson, I. Chromosome aberrations and their influence on the reproductive performance of domestic animals Y: A review. Z. Tierz. Ziichtungsbiol. 1980, 97, 176–195. [Google Scholar] [CrossRef]
- Bongso, T.A.; Jainudeen, M.R.; Lee, J.Y. Testicular hypoplasia in a bull with XX/XY chimerism. Cornell Vet. 1981, 71, 376–382. [Google Scholar]
- Seguin, B.E.; Zhang, T.Q.; Buoen, L.C.; Weber, A.F.; Ruth, G.R. Cytogenetic survey of Holstein bulls at a commercial artificial insemination company to determine prevalence of bulls with centric fusion and chimeric anomalies. J. Am. Vet. Med. Assoc. 2000, 216, 65–67. [Google Scholar] [CrossRef]
- McDaneld, T.G.; Kuehn, L.A.; Thomas, M.G.; Snelling, W.M.; Sonstegard, T.S.; Matukumalli, L.K.; Smith, T.P.L.; Pollak, E.J.; Keele, J.W. Y are you not pregnant: Identification of Y chromosome segments in female cattle with decreased reproductive efficiency1,2,3,4. J. Anim. Sci. 2012, 90, 2142–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Payen, E.; Pailhoux, E.; Merhi, R.A.; Gianquinto, L.; Kirszenbaum, M.; Locatelli, A.; Cotinot, C. Characterization of ovine SRY transcript and developmental expression of genes involved in sexual differentiation. Int. J. Dev. Biol. 1996, 40, 567–575. [Google Scholar]
- Bosu, W.T.; Basrur, P.K. Morphological and hormonal features of an ovine and a caprine intersex. Can. J. Comp. Med.: Rev. Can. de Med. Comp. 1984, 48, 402–409. [Google Scholar]
- Smith, M.C.; Dunn, H.O. Freemartin condition in a goat. J. Am. Vet. Med. Assoc. 1981, 178, 735–737. [Google Scholar] [PubMed]
- Bongso, T.; Robinson, E.; Fatimah, I.; Abeynayake, P. Foetal membrane fusion and its developmental consequences in goat twins. Br. Vet. J. 1986, 142, 59–64. [Google Scholar] [CrossRef]
- Santucciu, C.; Iannuzzi, L.; Fogu, G.; Bonelli, P.; Bogliolo, L.; Rosati, I.; Ledda, S.; Zedda, M.T.; Pau, S. Clinical and cytogenetic studies in intersex ewes. Caryologia 2006, 59, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Di Meo, G.; Neglia, G.; Perucatti, A.; Genualdo, V.; Iannuzzi, A.; Crocco, D.; Incarnato, D.; Romano, G.; Parma, P. Numerical Sex Chromosome Aberrations and Abnormal Sex Development in Horse and Sheep. Sex. Dev. 2009, 3, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Szczerbal, I.; Komosa, M.; Nowacka-Woszuk, J.; Uzar, T.; Houszka, M.; Semrau, J.; Musial, M.; Barczykowski, M.; Lukomska, A.; Switonski, M. A Disorder of Sex Development in a Holstein–Friesian Heifer with a Rare Mosaicism (60,XX/90,XXY): A Genetic, Anatomical, and Histological Study. Animals 2021, 11, 285. [Google Scholar] [CrossRef] [PubMed]
- Meinecke, B.; Kuiper, H.; Wohlsein, P.; Wehrend, A.; Meinecke-Tillmann, S.; Drögemüller, C.; Bürstel, D.; Ebeling, S. A Diploid-Triploid (60,XX/90,XXY) Intersex in a Holstein Heifer. Sex. Dev. 2006, 1, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.M., Jr.; Hoehn, H.; Lin, M.S.; Smith, D.W. Diploid-triploid mixoploidy: Clinical and cytogenetic aspects. Pediatrics 1981, 68, 23–28. [Google Scholar] [PubMed]
- Oktem, O.; Paduch, D.A.; Xu, K.; Mielnik, A.; Oktay, K. Normal Female Phenotype and Ovarian Development Despite the Ovarian Expression of the Sex-Determining Region of Y Chromosome (SRY) in a 46,XX/69,XXY Diploid/Triploid Mosaic Child Conceived afterin VitroFertilization–Intracytoplasmic Sperm Injection. J. Clin. Endocrinol. Metab. 2007, 92, 1008–1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Schepper, G.G.; Aalbers, J.G.; Brake, J.H.T. Double reciprocal translocation heterozygosity in a bull. Vet. Rec. 1982, 110, 197–199. [Google Scholar] [CrossRef] [PubMed]
- Mayr, B.; Krutzler, H.; Auer, H.; Schleger, W. Reciprocal translocation 60,XY, t(8;15) (21;24) in cattle. J. Reprod. Fertil. 1983, 69, 629–630. [Google Scholar] [CrossRef] [Green Version]
- Christensen, K.; Agerholm, J.S.; Larsen, B. Dairy breed bull with complex chromosome translocation: Fertility and linkage studies. J. Reprod. Fertil. 2008, 117, 199–202. [Google Scholar] [CrossRef]
- Kovacs, A.; Villagómez, D.; Gustavsson, I.; Lindblad, K.; Foote, R.; Howard, T. Synaptonemal complex analysis of a three-breakpoint translocation in a subfertile bull. Cytogenet. Genome Res. 1992, 61, 195–201. [Google Scholar] [CrossRef]
- De Giovanni, A.; Succi, G.; Molteni, L.; Castiglioni, M. A new autosomal translocation in “Alpine grey cattle”. Ann. Genet. Sel. Anim. 1979, 11, 115–120. [Google Scholar] [CrossRef]
- Ansari, H.; Jung, H.; Hediger, R.; Fries, R.; König, H.; Stranzinger, G. A balanced autosomal reciprocal translocation in an azoospermic bull. Cytogenet. Genome Res. 1993, 62, 117–123. [Google Scholar] [CrossRef]
- Villagómez, D.; Andersson, M.; Gustavsson, I.; Plöen, L. Synaptonemal complex analysis of a reciprocal translocation, rcp(20;24) (q17;q25), in a subfertile bull. Cytogenet. Genome Res. 1993, 62, 124–130. [Google Scholar] [CrossRef]
- Mayr, B.; Korb, H.; Kiendler, S.; Brem, G. Reciprocal X;1 translocation in a calf. Genet. Sel. Evol. 1998, 30, 305–308. [Google Scholar] [CrossRef]
- Ducos, A.; Dumont, P.; Seguela, A.; Pinton, A.; Berland, H.; Brun-Baronnat, C.; Darre, A.; Guienne, B.M.-L.; Humblot, P.; Boichard, D.; et al. A new reciprocal translocation in a subfertile bull. Genet. Sel. Evol. 2000, 32, 589–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannuzzi, L.; Molteni, L.; Di Meo, G.; Perucatti, A.; Lorenzi, L.; Incarnato, D.; De Giovanni, A.; Succi, G.; Gustavsson, I. A new balanced autosomal reciprocal translocation in cattle revealed by banding techniques and human-painting probes. Cytogenet. Cell Genet. 2001, 94, 225–228. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Molteni, L.; Di Meo, G.; De Giovanni, A.; Perucatti, A.; Succi, G.; Incarnato, D.; Eggen, A.; Cribiu, E. A case of azoospermia in a bull carrying a Y-autosome reciprocal translocation. Cytogenet. Cell Genet. 2001, 95, 225–227. [Google Scholar] [CrossRef] [PubMed]
- Molteni, L.; Perucatti, A.; Iannuzzi, A.; Di Meo, G.; De Lorenzi, L.; De Giovanni, A.; Incarnato, D.; Succi, G.; Cribiu, E.; Eggen, A.; et al. A new case of reciprocal translocation in a young bull: Rcp(11;21)(q28;q12). Cytogenet. Genome Res. 2007, 116, 80–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lorenzi, L.; De Giovanni, A.; Molteni, L.; Denis, C.; Eggen, A.; Parma, P. Characterization of a balanced reciprocal trans-location, rcp(9;11)(q27;q11) in cattle. Cytogenet. Genome Res. 2007, 119, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Switonski, M.; Andersson, M.; Nowacka-Woszuk, J.; Szczerbal, I.; Sosnowski, J.; Kopp, C.; Cernohorska, H.; Rubes, J. Identi-fication of a new reciprocal translocation in an AI bull by synaptonemal complex analysis, followed by chromosome painting. Cytogenet. Genome Res. 2008, 121, 245–248. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzi, L.; Kopecna, O.; Gimelli, S.; Cernohorska, H.; Zannotti, M.; Béna, F.; Molteni, L.; Rubes, J.; Parma, P. Reciprocal Translocation t(4;7)(q14;q28) in Cattle: Molecular Characterization. Cytogenet. Genome Res. 2010, 129, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Switonski, M.; Szczerbal, I.; Krumrych, W.; Nowacka-Woszuk, J. A case of Y-autosome reciprocal translocation in a Hol-stein-Friesian bull. Cytogenet. Genome Res. 2011, 132, 22–25. [Google Scholar] [CrossRef] [PubMed]
- Kochneva, M.L.; Zhidenova, A.N.; Biltueva, L.S.; Kiseleva, Y.T. A new cse of reciprocal translocation rcp(13;26) in cattle. Biologiya 2011, 6, 84–89. [Google Scholar]
- Biltueva, L.; Kulemzina, A.; Vorobieva, N.; Perelman, P.; Kochneva, M.; Zhidenova, A.; Graphodatsky, A. A New Case of an Inherited Reciprocal Translocation in Cattle: Rcp(13;26)(q24;q11). Cytogenet. Genome Res. 2014, 144, 208–211. [Google Scholar] [CrossRef] [PubMed]
- Jennings, R.L.; Griffin, D.K.; O’Connor, R.E. A New Approach for Accurate Detection of Chromosome Rearrangements That Affect Fertility in Cattle. Animals 2020, 10, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glahn-Luft, B.; Schneider, H.; Schneider, J.; Wassmuth, R. Agnathie beim Schaf mit Chromosomenaberrationen und Hb-Mangel [Agnathia in the sheep associated with chromosome aberration and Hb deficiency]. Dtsch. Tierarztl. Wochenschr. 1978, 85, 472–474. [Google Scholar]
- Anamthawat-Jonsson, K.; Long, S.; Basrur, P.; Adalsteinsson, S. Reciprocal translocation (13;20)(q12;q22) in an Icelandic sheep. Res. Vet. Sci. 1992, 52, 367–370. [Google Scholar] [CrossRef]
- Popescu, C.P.; Tixier, M. L’incidence des anomalies chromosomiques chez les animaux de ferme et leurs conséquences économiques [The frequency of chromosome abnormalities in farm animals and their economic consequences]. Ann. Genet. 1984, 27, 69–72. [Google Scholar]
- Popescu, C.P. Conséquences des anomalies de la structure chromosomique chez les animaux domestiques [Consequences of abnormalities of chromosome structure in domestic animals]. Reprod. Nutr. Dev. 1990, 30 (Suppl. 1), 105s–116s. [Google Scholar] [CrossRef]
- Slota, E.; Danielak, B.; Kozubska, A. Structural rearrangement of metacentric chromosomes in the ram of Polish wrzosówka breed. In Proceedings of the 7th European Colloquium on Cytogenetics Dom. Anim, Warsaw, Poland, 23–26 July 1986; p. 37. [Google Scholar]
- Iannuzzi, A.; Perucatti, A.; Genualdo, V.; De Lorenzi, L.; Di Berardino, D.; Parma, P. Cytogenetic Elaboration of a Novel Reciprocal Translocation in Sheep. Cytogenet. Genome Res. 2013, 139, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, A.; Perucatti, A.; Genualdo, V.; Pauciullo, A.; Incarnato, D.; Musilova, P.; Rubes, J.; Iannuzzi, C. The Utility of Chromosome Microdissection in Clinical Cytogenetics: A New Reciprocal Translocation in Sheep. Cytogenet. Genome Res. 2014, 142, 174–178. [Google Scholar] [CrossRef]
- Switonski, M.; Stranzinger, G. Studies of synaptonemal complexes in farm mammals—A review. J. Hered. 1998, 89, 473–480. [Google Scholar] [CrossRef] [Green Version]
- Basrur, P.K.; Koykul, W.; Baguma-Nibasheka, M.; King, W.A.; Ambady, S.; de León, F.A.P. Synaptic pattern of sex com-plements and sperm head malformation in X-autosome translocation carrier bulls. Mol. Reprod. Dev. 2001, 59, 67–77. [Google Scholar] [CrossRef]
- Rho, G.-J.; Coppola, G.; Sosnowski, J.; Kasimanickam, R.; Johnson, W.H.; Semple, E.; Mastromonaco, G.F.; Betts, D.H.; Koch, T.G.; Weese, S.; et al. Use of Somatic Cell Nuclear Transfer to Study Meiosis in Female Cattle Carrying A Sex-Dependent Fertility-Impairing X-Chromosome Abnormality. Cloning Stem Cells 2007, 9, 118–129. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzi, L.; Morando, P.; Planas, J.; Zannotti, M.; Molteni, L.; Parma, P. Reciprocal translocations in cattle: Frequency es-timation. J. Anim. Breed. Genet. 2012, 129, 409–416. [Google Scholar] [CrossRef] [Green Version]
- Wilch, E.S.; Morton, C.C. Historical and Clinical Perspectives on Chromosomal Translocations. Adv. Exp. Med. Biol. 2018, 1044, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lojda, L.; Rubes, J.; Staisksova, M.; Havrandsova, J. Chromosomal findings in some reproductive disorders in bulls. In Proceedings of the 8th International Congress Animal Reproduction Artificial Insemination, Krakow, Poland, 12–16 July 1976; Volume 158, p. 141. [Google Scholar]
- Garick, D.J.; Ruvinsky, A. The Gentic of Cattle; CAB International: Wallingford, UK, 2015. [Google Scholar]
- Miyake, Y.; Murakami, R.K.; Kaneda, Y. Inheritance of the Robertsonian translocation (1/21) in the Holstein-Friesian cattle. I. Chromosome analysis. J. Vet. Med. Sci. 1991, 53, 113–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce, P.; Ansari, H.; Maher, D.; Amarante, M.; Monro, T.; Hendrikse, W. 1/25 translocations in Blonde d’Aquitaine cattle in New Zealand. N. Z. Vet. J. 1997, 45, 69–71. [Google Scholar] [CrossRef]
- Stranzinger, G.F.; Förster, M. Autosomale Chromosomentranslokationen beim Fleck- und Braunvieh [Autosomal chromo-sometranslocation of piebald cattle and brown cattle (author’s transl)]. Experientia 1976, 32, 24–27. (In German) [Google Scholar] [CrossRef]
- Miyake, Y.; Kaneda, Y. A new type of Robertsonian translocation (1/26) in a bull with unilateral cryptorchidism, probably occurring de novo. Nihon Juigaku Zasshi 1987, 49, 1015–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eldridge, F.E. High frequency of a Robertsonian translocation in a herd of British White cattle. Vet. Rec. 1975, 97, 71–73. [Google Scholar] [CrossRef]
- Pollock, D.L.; Bowman, J.C. A Robertsonian Translocation in British Friesian Cattle. J. Reprod. Fertil. 1974, 40, 423–432. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Yamamoto, Y.; Amano, T.; Yamagata, T.; Dang, V.-C.; Matsuda, Y.; Namikawa, T. A Robertsonian Translocation, Rob(2;28), Found in Vietnamese Cattle. Hereditas 2000, 133, 19–23. [Google Scholar] [CrossRef]
- Popescu, C.P. Observations sur le caryotype normal et anormal des bovins [Normal and abnormal karyotypes of cattle]. Can. Vet. J. 1977, 18, 143–149. [Google Scholar] [PubMed]
- Barasc, H.; Mouney-Bonnet, N.; Peigney, C.; Calgaro, A.; Revel, C.; Mary, N.; Ducos, A.; Pinton, A. Analysis of Meiotic Seg-regation Pattern and Interchromosomal Effects in a Bull Heterozygous for a 3/16 Robertsonian Translocation. Cytogenet. Genome Res. 2018, 156, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Bouvet, A.; Popescu, C.P.; De Giovanni-Macchi, A.M.; Colombo, G.; Molteni, L. Synaptonemal complexes analysis in a bull carrying a 4;8 Robertsonian translocation. Ann. Génét. 1989, 32, 193–199. [Google Scholar] [PubMed]
- Bahri-Darwich, I.; Cribiu, E.; Berland, H.; Darré, R. A new Robertsonian translocation in Blonde d’Aquitaine cattle, rob(4;10). Genet. Sel. Evol. 1993, 25, 413–419. [Google Scholar] [CrossRef]
- Papp, M.; Kovacs, A. 5/18 dicentric Robertsonian translocation in a Simmental bull. In Proceedings of the 4th Europe Colloquium Cytogen Domestic Animals, Milano Gargnano, Italy, 7–11 June 1980; p. 51. [Google Scholar]
- Slota, E.; Switonski, M. A new Robertsonian translocation 5;22 in cattle. Studies of banded chromosomes and synaptonemal complexes. Genet. Pol. 1992, 33, 227–231. [Google Scholar]
- Di Meo, G.P.; Molteni, L.; Perucatti, A.; De Giovanni, A.; Incarnato, D.; Succi, G.; Schibler, L.; Cribiu, E.P.; Iannuzzi, L. Chro-mosomal characterization of three centric fusion translocations in cattle using G-, R- and C-banding and FISH technique. Caryologia 2000, 53, 213–218. [Google Scholar] [CrossRef]
- Tateno, H.; Miyake, Y.I.; Mori, H.; Kamiguchi, Y.; Mikamo, K. Sperm chromosome study of two bulls heterozygous for dif-ferent Robertsonian translocations. Hereditas 1994, 120, 7–11. [Google Scholar] [CrossRef]
- Hanada, H.; Muramatsu, S.; Abe, T.; Fukushima, T. Robertsonian chromosome polymorphism found in a local herd of the Japanese Black cattle. Ann. Genet. Sel. Anim. 1981, 13, 205–211. [Google Scholar] [CrossRef]
- Cribiu, E.P.; Matejka, M.; Darre, R.; Durand, V.; Berland, H.M.; Bouvet, A. Identification of chromosomes involved in a Rob-ertsonian translocation in cattle. Genet. Sel. Evol. 1989, 21, 555–560. [Google Scholar] [CrossRef]
- Pinheiro, L.E.L.; Ferrari, L. A new type of Robertsonian translocation in cattle. In Proceedings of the 5th Encontro de Pesquisas Veterinarias, Jaboticabal, Brazil, 6–7 November 1980; p. 161. [Google Scholar]
- Kovács, A.; Papp, M. Report on chromosomal examination of A.I. bulls in Hungary. Ann. Génét. Sél. Anim. 1977, 9, 528. [Google Scholar] [CrossRef]
- Holecková, B.; Sutiaková, I.; Pijáková, N. Robertsonian translocation in a cattle population. Vet. Med. 1995, 40, 33–34. [Google Scholar]
- Molteni, L.; Giovanni-Macchi, A.; Succi, G.; Cremonesi, F.; Stacchezzini, S.; Meo, G.P.; Iannuzzi, L. A New Centric Fusion Translocation in Cattle: Rob (13;19). Hereditas 2004, 129, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Kovács, A.; Mészáros, I.; Sellyei, M.; Vass, L. Mosaic centromeric fusion in a Holstein-Friesian bull. Acta Boil. Acad. Sci. Hung. 1973, 24, 215–220. [Google Scholar]
- Slota, E.; Danielak, B.; Kozubska, A. The Robertsonian translocation in cattle quintuplets. In Proceedings of the 8th European Colloquium on Cytogenetics of Domestic Animals, Bristol, UK, 9–22 July 1988; pp. 122–124. [Google Scholar]
- De Giovanni Macchi, A.; Molteni, L.; Parma, P.; Laurelli, A. Identification of a new Robertsonian translocation in the Marchigiana breed. In Proceedings of the 8th North American Colloquium on Domestic Animal Cytogenetics and Gene Mapping, Guelph, ON, Canada, 13–16 July 1993; p. 175. [Google Scholar]
- De Lorenzi, L.; Molteni, L.; De Giovanni, A.; Parma, P. A new case of rob(14;17) in cattle. Cytogenet. Genome Res. 2008, 120, 144–146. [Google Scholar] [CrossRef] [PubMed]
- Logue, D.N.; Harvey, M.J. A 14/20 Robertsonian translocation in Swiss Simmental cattle. Res. Vet. Sci. 1978, 25, 7–12. [Google Scholar] [CrossRef]
- Schmutz, S.; Moker, J.; Pawlyshyn, V.; Haugen, B.; Clark, E. Fertility effects of the 14;20 Robertsonian translocation in cattle. Theriogenology 1997, 47, 815–823. [Google Scholar] [CrossRef]
- Weber, A.F.; Buoen, L.C.; Zhang, T.; Ruth, G.R. Prevalence of the 14/20 centric fusion chromosomal aberration in US Simmental cattle. J. Am. Vet. Med. Assoc. 1992, 200, 1216–1219. [Google Scholar] [PubMed]
- Di Berardino, D.; Iannuzzi, L.; Ferrara, L.; Matassino, D. A new case of Robertsonian translocation in cattle. J. Hered. 1979, 70, 436–438. [Google Scholar] [CrossRef]
- Ellsworth, S.M.; Paul, S.R.; Bunch, T.D. A 14/28 dicentric Robertsonian translocation in a Holstein cow. Theriogenology 1979, 11, 165–171. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Rangel-Figueiredo, T.; Di Meo, G.; Ferrara, L. A new Robertsonian translocation in cattle, rob(15;25). Cytogenet. Genome Res. 1992, 59, 280–283. [Google Scholar] [CrossRef]
- Iannuzzi, L.; Rangel-Figueiredo, T.; Meo, G.P.; Ferrara, L. A New Centric Fusion Translocation in Cattle, Rob(16;18). Hereditas 2004, 119, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Rubes, J.; Musilová, P.; Borkovec, L.; Borkovcová, Z.; Svecová, D.; Urbanová, J. A new Robertsonian translocation in cattle, rob(16;20). Hereditas 1996, 124, 275–279. [Google Scholar] [CrossRef]
- Rybar, R.; Horakova, J.; Machatkova, M.; Hanzalova, K.; Rubes, J. Embryos produced in vitro from bulls carrying 16;20 and 1;29 Robertsonian translocations: Detection of translocations in embryos by fluorescence in situ hybridization. Zygote 2005, 13, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Pinton, A.; Ducos, A.; Berland, H.; Séguéla, A.; Blanc, M.F.; Darré, A.; Mimar, S.; Darré, R. A new Robertsonian translocation in Holstein-Friesian cattle. Genet. Sel. Evol. 1997, 29, 523–526. [Google Scholar] [CrossRef]
- Berland, H.M.; Cribiu, E.P.; Darre, R.; Boscher, J.; Popescu, C.P.; Sharma, A. A New Case of Robertsonian Translocation in Cattle. J. Hered. 1988, 79, 33–36. [Google Scholar] [CrossRef]
- De Lorenzi, L.; Molteni, L.; Denis, C.; Eggen, A.; Parma, P. A new case of centric fusion in cattle: Rob(21;23). Anim. Genet. 2008, 39, 454–455. [Google Scholar] [CrossRef] [PubMed]
- Molteni, L.; Meggiolaro, D.; Macchi, A.D.G.; De Lorenzi, L.; Crepaldi, P.; Stacchezzini, S.; Cremonesi, F.; Ferrara, F. Fertility of cryopreserved sperm in three bulls with different Robertsonian translocations. Anim. Reprod. Sci. 2005, 86, 27–36. [Google Scholar] [CrossRef]
- Bongso, A.; Basrur, P.K. Chromosome anomalies in Canadian Guernsey bulls. Cornell Vet. 1976, 66, 476–489. [Google Scholar] [PubMed]
- Di Meo, G.; Perucatti, A.; Genualdo, V.; Iannuzzi, A.; Sarubbi, F.; Jambrenghi, A.C.; Incarnato, D.; Peretti, V.; Vonghia, G.; Iannuzzi, L. A Rare Case of Centric Fission and Fusion in a River Buffalo (Bubalus bubalis, 2n = 50) Cow with Reduced Fertility. Cytogenet. Genome Res. 2011, 132, 26–30. [Google Scholar] [CrossRef]
- Albarella, S.; Ciotola, F.; Coletta, A.; Genualdo, V.; Iannuzzi, L.; Peretti, V. A new translocation t(1p;18) in an Italian Medi-terranean river buffalo (Bubalus bubalis, 2n = 50) bull: Cytogenetic, fertility and inheritance studies. Cytogenet. Genome Res. 2013, 139, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Bruère, A.; Mills, R.A. Observations on the incidence of Robertsonian translocations and associated testicular changes in a flock of New Zealand Romney sheep. Cytogenet. Genome Res. 1971, 10, 260–272. [Google Scholar] [CrossRef] [PubMed]
- Ansari, H.A.; Pearce, P.D.; Maher, D.W.; Malcolm, A.A.; Broad, T.E. Resolving ambiguities in the karyotype of domestic sheep (Ovis aries). Chromosoma 1993, 102, 340–347. [Google Scholar] [CrossRef] [PubMed]
- Bruère, A.; Chapman, H.M.; Wyllie, D.R. Chromosome polymorphism and its possible implications in the select Drysdale breed of sheep. Cytogenet. Genome Res. 1972, 11, 233–246. [Google Scholar] [CrossRef]
- Pearce, P.D.; Ansari, H.A.; Maher, D.W.; Malcolm, A.A.; Stewart-Scott, I.A.; Broad, T.E. New Robertsonian translocation chromosomes in domestic sheep (Ovis aries). Cytogenet. Genome Res. 1994, 67, 137–140. [Google Scholar] [CrossRef]
- Glahn-Luft, B.; Wassmuth, R. The influence of 1/20 translocation in sheep on the efficiency of reproduction. In Proceedings of the 31st Annual Meeting of the European Association for Animal Production, München, Germany, 1–4 September 1980. [Google Scholar]
- Chaves, R.; Adega, F.; Wienberg, J.; Guedes-Pinto, H.; Heslop-Harrison, J.S. Molecular cytogenetic analysis and centromeric satellite organization of a novel 8;11 translocation in sheep: A possible intermediate in biarmed chromosome evolution. Mamm. Genome 2003, 14, 706–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popescu, C.P. The mode of transmission of a centric fusion to the offspring of a buck (Capra hircus L.). Ann. Genet. Sel. Anim. 1972, 4, 355–361. [Google Scholar] [PubMed]
- Evans, H.J.; Buckland, R.A.; Sumner, A.T. Chromosome homology and heterochromatin in goat, sheep and ox studied by banding techniques. Chromosoma 1973, 42, 383–402. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, H.; Jorge, W.; Cury, P. Distribution of a Robertsonian translocation in goats. Small Rumin. Res. 1992, 8, 345–352. [Google Scholar] [CrossRef]
- Elminger, B.; Stranzinger, C. Identification of a centromeric fusion in the G-banding karyotype of a Saanen goat. In Proceedings of the 5th European Colloquium on Cytogenetic of Domestic Animals, Milano Gargano, Italy, 7–11 June 1982; pp. 407–409. [Google Scholar]
- Dolf, J.; Hediger, R. Comparison of centric fusion in a Toggenburg and Saanen goat. In Proceedings of the 6th European Colloquium on Cy-togenetic of Domestic Animals, Zürich, Switzerland, 16–20 July 1984; pp. 311–312. [Google Scholar]
- Burguete, I.; Di Berardino, D.; Lioi, M.B.; Taibi, L.; Matassino, D. Cytogenetic observations on a Robertsonian translocation in Saanen goats. Genet. Sel. Evol. 1987, 19, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Guillemot, E.; Gary, F.; Berland, H.M.; Berthelot, X.; Durand, V.; Darre, R.; Cribiu, E.P. Effects of 6/15 Robertsonian translo-cation in Saanen goats. Reprod. Domest. Anim. 1993, 28, 28–32. [Google Scholar] [CrossRef]
- Da Mota, L.S.L.S.; da Silva, R.A.B. Centric fusion in goats (Capra hircus): Identification of a 6/15 translocation by high resolution chromosome banding. Genet. Mol. Biol. 1998, 21. [Google Scholar] [CrossRef]
- Moreno-Millan, M.; Rodero-Franganillo, A. A new Robertsonian translocation in an intersex goat: Morphometric determi-nation. Arch. Zootec. 1990, 39, 263–270. [Google Scholar]
- De Lorenzi, L.; Genualdo, V.; Gimelli, S.; Rossi, E.; Perucatti, A.; Iannuzzi, A.; Zannotti, M.; Malagutti, L.; Molteni, L.; Iannuzzi, L.; et al. Genomic analysis of cattle rob(1;29). Chromosom. Res. 2012, 20, 815–823. [Google Scholar] [CrossRef]
- Di Meo, G.P.; Perucatti, A.; Chaves, R.; Adega, F.; De Lorenzi, L.; Molteni, L.; De Giovanni, A.; Incarnato, D.; Guedes-Pinto, H.; Eggen, A.; et al. Cattle rob(1;29) originating from complex chromosome rearrangements as revealed by both banding and FISH-mapping techniques. Chromosom. Res. 2006, 14, 649–655. [Google Scholar] [CrossRef]
- Chaves, R.; Heslop-Harrsion, J.S.; Guedes-Pinto, H. Centromeric heterochromatin in the cattle rob(1;29) translocation: Al-pha-satellite I sequences, in-situ MspI digestion patterns, chromomycin staining and C-bands. Chromosom. Res. 2000, 8, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Popescu, C.P.; Pech, A. Une bibliographie sur la translocation 1/29 de bovins dans le monde (1964–1990). Ann. Zootech. 1991, 40, 271–305. [Google Scholar] [CrossRef]
- Iannuzzi, A.; Di Meo, G.; Jambrenghi, A.C.; Vonghia, G.; Iannuzzi, L.; Rangel-Figueiredo, T. Frequency and distribution of rob(1;29) in eight Portuguese cattle breeds. Cytogenet. Genome Res. 2008, 120, 147–149. [Google Scholar] [CrossRef]
- Switoński, M.; Gustavsson, I.; Plöen, L. The nature of the 1;29 translocation in cattle as revealed by synaptonemal complex analysis using electron microscopy. Cytogenet. Genome Res. 1987, 44, 103–111. [Google Scholar] [CrossRef]
- Bonnet-Garnier, A.; Pinton, A.; Berland, H.M.; Khireddine, B.; Eggen, A.; Yerle, M.; Darré, R.; Ducos, A. Sperm nuclei analysis of 1/29 Robertsonian translocation carrier bulls using fluorescence in situ. Cytogenet. Genome Res. 2006, 112, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Di Dio, C.; Longobardi, V.; Zullo, G.; Parma, P.; Pauciullo, A.; Perucatti, A.; Higgins, J.; Iannuzzi, A. Analysis of meiotic segregation by triple-color fish on both total and motile sperm fractions in a t(1p;18) river buffalo bull. PLoS ONE 2020, 15, e0232592. [Google Scholar] [CrossRef]
- Rubes, J.; Machatková, M.; Jokesová, E.; Zudová, D. A potential relationship between the 16;20 and 14;20 Robertsonian translocations and low in vitro embryo development. Theriogenology 1999, 52, 171–180. [Google Scholar] [CrossRef]
- Vozdova, M.; Kubíčková, S.; Cernohorska, H.; Rubeš, J. Detection of translocation rob(1;29) in bull sperm using a specific DNA probe. Cytogenet. Genome Res. 2008, 120, 102–105. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L. Standard karyotype of the river buffalo (Bubalus bubalis L., 2n = 50). Report of the committee for the standardization of banded karyotypes of the river buffalo. Cytogenet. Cell Genet. 1994, 67, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Broad, T.E.; McLean, N.; Grimaldi, D.; Monk, N. A novel occurrence in sheep (Ovis aries) of four homozygous Robertsonian translocations. Hereditas 2000, 132, 165–166. [Google Scholar] [CrossRef]
- Broad, T.E.; Hayes, H.; Long, S.E. Cytogenetics: Physical maps. In The Genetics of Sheep; Piper, L., Ruvinsky, A., Eds.; CAB International: Wallingford, UK, 1997; pp. 241–296. [Google Scholar]
- Vallenzasca, C.; Martignoni, M.; Galli, A. Finding of a bull with Y;17 translocation. Hereditas 1990, 113, 63–67. [Google Scholar] [CrossRef]
- Gallagher, D.S., Jr.; Basrur, P.K.; Womack, J.E. Identification of an autosome to X chromosome translocation in the domestic cow. J. Hered. 1992, 83, 451–453. [Google Scholar] [CrossRef]
- Pinton, A.; Ducos, A.; Yerle, M. Chromosomal rearrangements in cattle and pigs revealed by chromosome microdissection and chromosome painting. Genet. Sel. Evol. 2003, 35, 1–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popescu, C.; Boscher, J. New data on pericentric inversion in cattle (Bos taurus L). Ann. Génét. Sél. Anim. 1976, 8, 443. [Google Scholar] [CrossRef]
- Switoński, M. A pericentric inversion in an X chromosome in the cow. J. Hered. 1987, 78, 58–59. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, L.; Di Meo, G.; Perucatti, A.; Eggen, A.; Incarnato, D.; Sarubbi, F.; Cribiu, E. A pericentric inversion in the cattle Y chromosome. Cytogenet. Cell Genet. 2001, 94, 202–205. [Google Scholar] [CrossRef] [PubMed]
- De Lorenzi, L.; Iannuzzi, A.; Rossi, E.; Bonacina, S.; Parma, P. Centromere Repositioning in Cattle (Bos taurus) Chromosome 17. Cytogenet. Genome Res. 2017, 151, 191–197. [Google Scholar] [CrossRef]
- Hansen, K.M. Bovine tandem fusion and fertility. Hereditas 1969, 63, 453–454. [Google Scholar]
- Pinheiro, L.E.; Carvalho, T.B.; Oliveira, D.A.; Popescu, C.P.; Basrur, P.K. A 4/21 tandem fusion in cattle. Hereditas 1995, 122, 99–102. [Google Scholar] [CrossRef]
- Halnan, C.R. Autosomal deletion and infertility in cattle. Vet. Rec. 1972, 91, 572. [Google Scholar] [CrossRef] [PubMed]
- Uzar, T.; Szczerbal, I.; Serwanska-Leja, K.; Nowacka-Woszuk, J.; Gogulski, M.; Bugaj, S.; Switonski, M.; Komosa, M. Congenital Malformations in a Holstein-Fresian Calf with a Unique Mosaic Karyotype: A Case Report. Animals 2020, 10, 1615. [Google Scholar] [CrossRef] [PubMed]
- Braun, U.; Ansari, H.A.; Hediger, R.; Süss, U.; Ehrensperger, F. Hypotrichose und Oligodontie, verbunden mit einer Xq-Deletion, bei einem Kalb der Schweizerischen Fleckviehrasse [Hypotrichosis and oligodontia, combined with an Xq-deletion, in a calf of the Swiss Holstein breed]. Tierarztl. Prax. 1988, 16, 39–44. [Google Scholar] [PubMed]
- Sharshov, A.A.; Grafodatskiĭ, A.S. Novaia perestroĭka khromosom u krupnogo rogatogo skota (A new chromosomal rear-rangement in cattle). Tsitol. Genet. 1990, 24, 30–33. [Google Scholar]
- De Lorenzi, L.; Rossi, E.; Genualdo, V.; Gimelli, S.; Lasagna, E.; Iannuzzi, A.; Parma, P.; Perucatti, A. Molecular Characterization of Xp Chromosome Deletion in a Fertile Cow. Sex. Dev. 2012, 6, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Lasagna, E.; (University of Perugia, Perugia, Italy). Personal communication, 2020.
- Iannuzzi, L.; Di Meo, G.; Perucatti, A.; Rullo, R.; Incarnato, D.; Longeri, M.; Bongioni, G.; Molteni, L.; Galli, A.; Zanotti, M.; et al. Comparative FISH-mapping of the survival of motor neuron gene (SMN) in domestic bovids. Cytogenet. Genome Res. 2003, 102, 39–41. [Google Scholar] [CrossRef]
- De Lorenzi, L.; Arrighi, S.; Rossi, E.; Grignani, P.; Previderè, C.; Bonacina, S.; Cremonesi, F.; Parma, P. XY (SRY-positive) Ovarian Disorder of Sex Development in Cattle. Sex. Dev. 2018, 12, 196–203. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chromosome Abnormalities | ||
|---|---|---|
| Numerical | Structural | |
| Autosomes | Sex Chromosomes | |
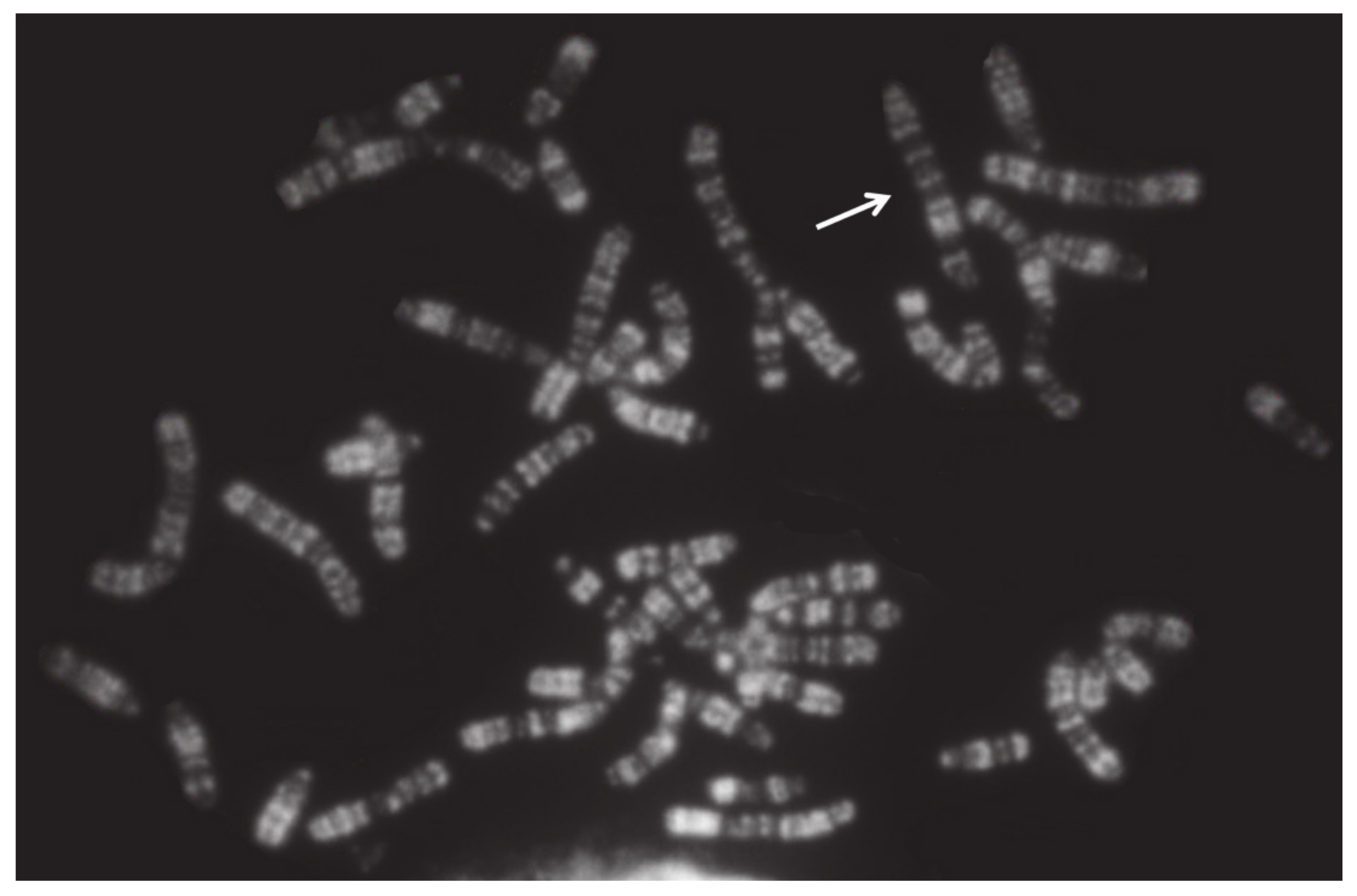
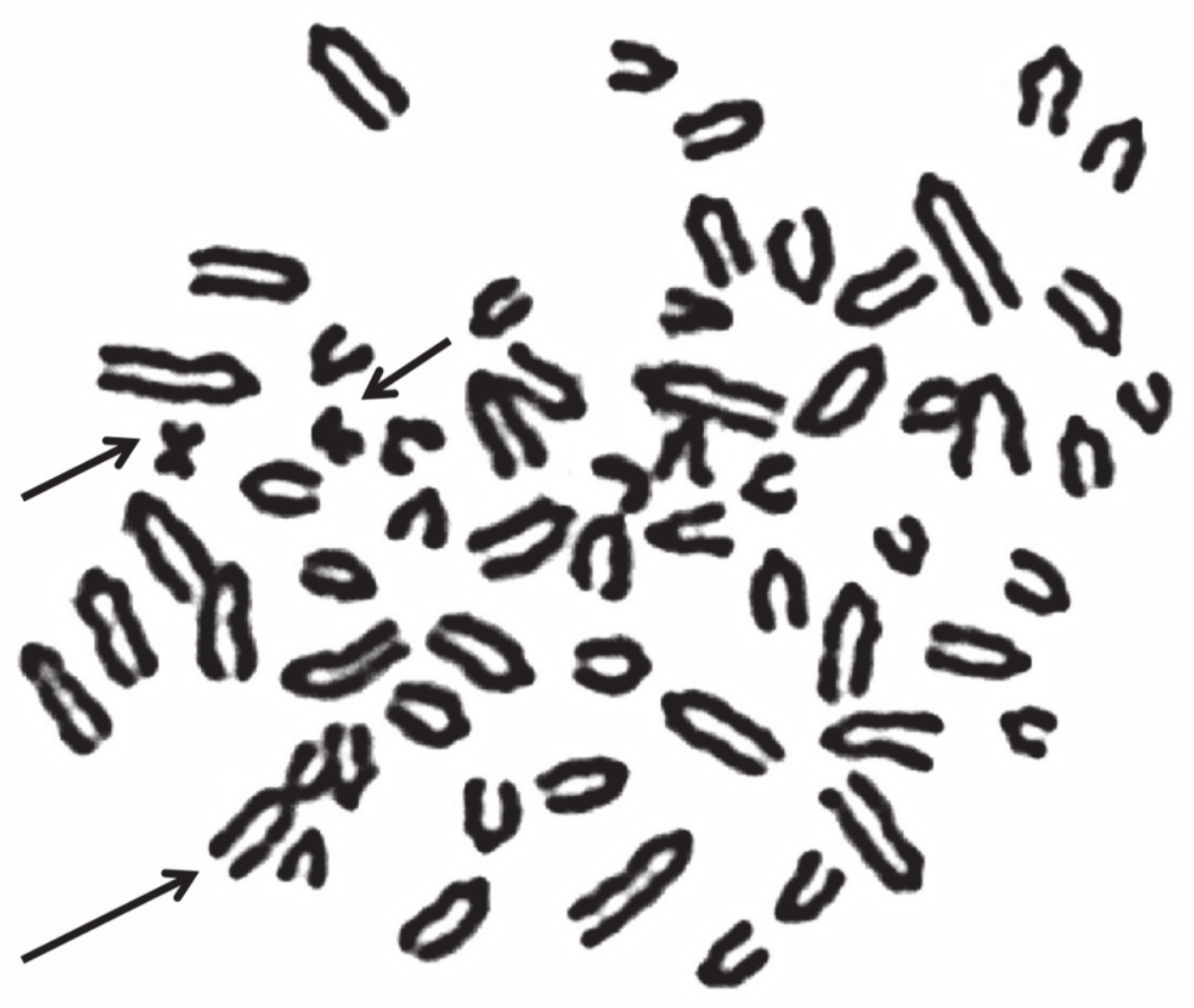
| Very rare (the animal body conformation being abnormal; these abnormalities are eliminated directly by the breeders) | More tolerated by the species but almost all related to sterility or low fertility, especially in the females Generally not visible in the carriers (normal body conformation and external genitalia) | Deviation from the normal chromosome shape or gene order Very important for the (a) high percentage of carriers (i.e., cattle rob1;29); (b) normal body conformation; (c) because they escape the normal breeding selection They can be balanced (translocations and inversions) or unbalanced (deletions, insertions, and duplications) |
| Chromosome Involved | Phenotype | References |
|---|---|---|
| Large Autosome | Male calf with extreme brachygnathia inferior | [21] |
| 12 | Anatomical defect, lethal | [22,23] |
| 16 (TAN,1;16) | Anatomical defects | [24] |
| 18 (?) | Anatomical defects | [25] |
| 19 | Anatomical defects (BI) | [26] |
| 20 | Sterile cow | [27] |
| Malformed calf, absence of external genitalia | [28] | |
| Malformed fetus, cranial defects | [29] | |
| Fetus with pulmonary hypoplasia and anasarca syndrome (genomic analysis) | [30] | |
| 21 (?) | Anatomical defects | [31] |
| 21 | Newborn Hereford with a cleft palate, hydrocephalus, a cardiac interventricular septal defect, and arthrogryposis | [32] |
| 22 | Anatomical defects (no lethality) [33] Multiple malformations, including hypoplasia of palpebral fissures, cleft palate, kyphoscoliosis, and arthrogryposis | [32,33,34] |
| 21 and 27 | Fetuses | [35,36] |
| 22 1 | Anatomical defects | [18] |
| Anatomical defects | [19] | |
| 24 | Malformed heifer (slight prognathia, heart defects, slow growth rate) | [37] |
| 26 | Sterility, growth retardation | [38] |
| 25 +;11− | Anatomical defects | [39] |
| 28 1 | Anatomical defects | [20] |
| 29 | Malformed female calf showing dwarfism with severe facial anomalies (genomic analysis) | [40] |
| Species | Phenotype | Reference |
|---|---|---|
| Cattle | Meiotic disturbances, familiar disposition, infertility | [47] |
| Infertility | [48] | |
| Infertility | [22] | |
| Infertility | [49] | |
| Continuous estrus | [50] | |
| Infertility | [51] | |
| Infertility, 2 cases | [52] | |
| R. Buffalo | Sterile (damages to internal sex structures) | [53] |
| Sterile (damages to internal sex structures) | [54] | |
| Sterile (damages to internal sex structures), male traits | [55] |
| Species | Phenotype | Reference |
|---|---|---|
| Cattle | Gonadal disgenesis (sterility) | [57] |
| Gonadal disgenesis (sterility) | [58] | |
| Body smaller in size, the uterus and uterine tubes appeared immature and inactive. | [59] | |
| Infertile heifer (XY/X0/Y-isochromosome) | [60] | |
| R. Buffalo | Gonadal disgenesis (sterility) | [61] |
| Gonadal disgenesis (sterility) | [62] | |
| Gonadal disgenesis (sterility) | [63] | |
| Sheep | Normal phenotype and external genitalia, no nursing of offspring | [64] |
| Gonadal dygenesis in the X0/XX karyotype | [65] | |
| Dizygotic sheep twins with internal sex damages and mammary gland development very limited | [66] | |
| Goat | Gonadal dysgenesis (XO/XX/XXX mixoploidy) | [67] |
| Species | Phenotype | References |
|---|---|---|
| Cattle | Testicular hypoplasia in a mosaicism case XY/XX/XXY | [69] |
| Testicular hypoplasia | [70] | |
| Testicular hypoplasia | [22] | |
| Intersexuality in a mosaicism case XX/XXY | [71] | |
| Bilateral testicular hypoplasia | [72] | |
| Testicular hypoplasia | [11] | |
| Testicular hypoplasia in a mosaicism case XX/XYY | [73] | |
| Masculinization effects in a mosaicism case XX/XXY | [70] | |
| Testicular hypoplasia | [74] | |
| Testicular hypoplasia (XXY + rob(1;29)) | [75] | |
| 2 cases (testicular hypoplasia with degradation of seminiferous tubules in one examined case) | [76] | |
| Azospermic bull | [77] | |
| Testicular hypoplasia in a bull with mosaicism (XY/XYY) | [78] | |
| Testicular hypoplasia | [79] | |
| Testicular hypoplasia | [80] | |
| Testicular hypoplasia in 3 cases | [52] | |
| Young male excluded for reproduction being mosaic for XY/XYY | Present Study | |
| R. Buffalo | Testicular hypoplasia in a case of 2n = 50,Y, rob(X;X) | [81] |
| Sheep | 2 cases in rams showing hypoplastic testis | [82] |
| Ram with no particular phenotypic effects (XX/XYY mosaicism) | [83] | |
| Goat | Testicular hypoplasia in a case of XXY/XY mosaicism | [84] |
| XX/XXY fertile buck | [85] |
| Species | Sex Chrom. | Phenotype/Effects on Fertility | Reference |
|---|---|---|---|
| Cattle | XY | Female (2) with reproductive defects | [86] |
| XY | Female with internal sex anatomical defects and no estrus | [87] | |
| XY | Female with no estrus and streak gonads | [88] | |
| XY | Female with hypoplastic ovaries | [89] | |
| XY | Single birth female with normal internal sex adducts but feeble estrus | [27] | |
| XY | Female normal gonads and genital development with AMGY and ZFY genes present (no SRY determination) | [90] | |
| XY | Female with hypoplastic gonads (the right one resembled an ovary and the left one an undeveloped testis) | [91] | |
| XY | Females (3) with no estrus and abnormal Y (Yp-iso) | [92,93] | |
| XX | Male with both testis and ovotestis development | [94] | |
| XX | Male XX + rob(1;29) apparently with the normal reproductive parameters but eliminated for rob(1;29) | [95] | |
| R. buffalo | XY | Females (2) sterile with abnormal internal sex adducts (one case with SRY-positive) | [55,96] |
| Sheep | XY | Sterile ewe with streak gonads, SRY+ | [97] |
| XY | Ewe with a longer ano-vulvar distance, enlarged clitoris, two testes-like structures at the inguinal level | [98] | |
| Goat | XX | Testicular biosynthesis of testosterone | [99] |
| XX | Males intersex, SRY-, Polled Intersex Syndrome (PIS) | [100,101,102] |
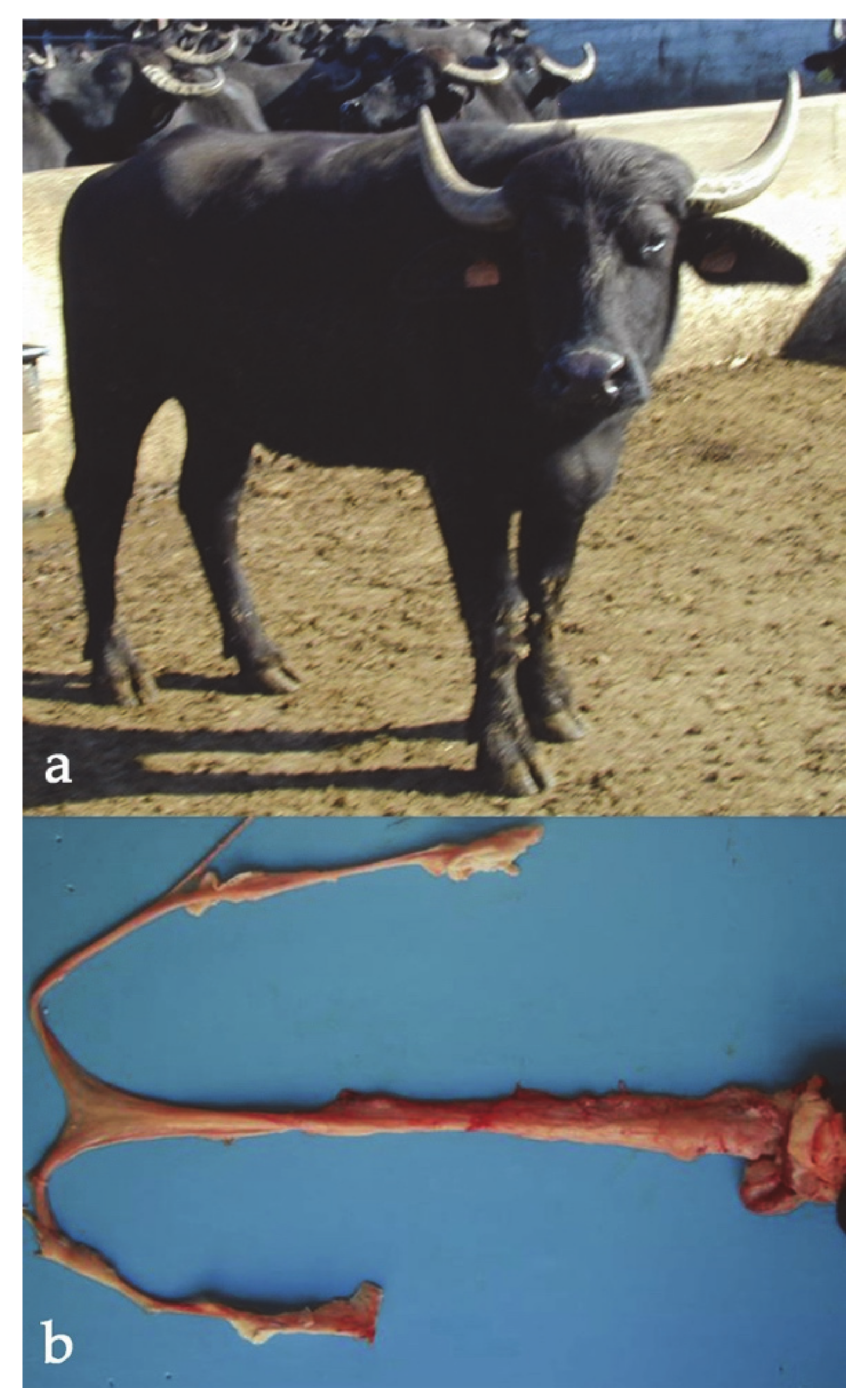
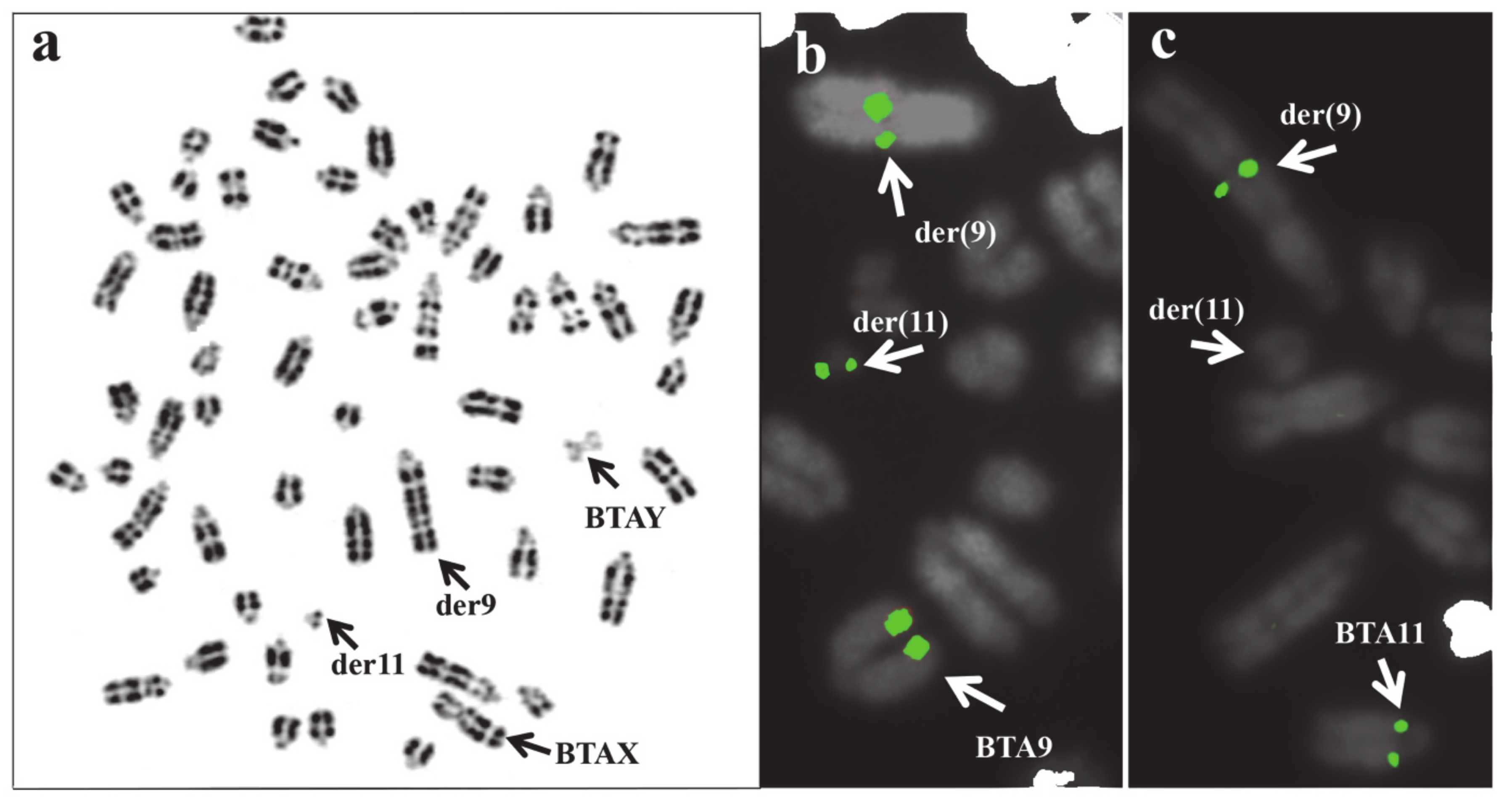
| Species | Rcp/Chrom. Involved | Phenotype | Reference |
|---|---|---|---|
| Cattle | double rcp(2q−;20q +, 8q-;27q +) | reduced fertility | [135] |
| rcp(8;15) (q21;q24) | reduced fertility | [136] | |
| rcp(1;8) (q44:q16) | 2 males and 3 females, reduced fertility | [137] | |
| rcp(1;8;9) (q43;q13;q26) | subfertile bull subfertile bulls (n = 3) | [138,139] | |
| rcp(8;13) (q11;q24) | azoospemic bull | [140] | |
| rcp(20;24) (q17;q25) | subfertile bull | [141] | |
| rcp(X;1) (42;13) | normal female calf with mosaicism XX/XY | [142] | |
| rcp(12;17) (q22;q14) | subfertile bull | [143] | |
| rcp(1;5) (q21;q35) | azoospermic bull and its dam (reduced fertility) | [144] | |
| rcp(Y;9) (q12.3;q2.1) | azoospermic bull | [145] | |
| rcp(11;21) (q28;q12) | bull, no libido, rare spermatozoa | [146] | |
| rcp(9;11) (q27;q11) | male addressed to reproduction | [147] | |
| rcp(2;4) (q45;q34) | bull (post mortem SC-analysis) | [148] | |
| rcp(4;7) (q14;q28) | bull, balanced, cyto-genomic analysis (CGH-arrays) | [149] | |
| rcp(Y;21) (p11;q11) | bull testosterone negative | [150] | |
| rcp(11;25) (q24;q11) | cow with reduced fertility | [41] | |
| rcp(13;26) | cow with reduced fertility | [151] | |
| rcp(5;6) (q13;q34) | bull, balanced, cyto-genomic analysis (CGH-arrays) | [16] | |
| rcp(13;26) (q24;q11) | dam and calf, balanced | [152] | |
| rcp(12;23) | two subfertile bulls | [153] | |
| Sheep | rcp(1p;19q) | low fertility | [154] |
| rcp(13;20) (q12;q22) | low fertility | [155] | |
| rcp(2q;3q) | low fertility | [156,157] | |
| rcp(2p−;3q +) | low fertility | [80,158] | |
| rcp(4q;12q) (q13;q25) | low fertility | [159] | |
| rcp(18;23) (q14;q26) | low fertility | [160] | |
| rcp(13;20) (q12;q22) | poor fertility | [155] |
| Species | Rob/Chrom. | Breed/Country | Reference | |
|---|---|---|---|---|
| Cattle | 1 | 4 | Czech Republic | [166] |
| - | 7 | Not reported | [167] | |
| - | Blond D’Aquitaine, France | [80] | ||
| - | 21 | Friesian | [168] | |
| - | 22 | Czech Republic | [166] | |
| - | 23 | Czech Republic | [166] | |
| - | 25 | Blonde d’Aquitaine, N.Z. Piebald cattle Germany | [169,170] | |
| - | 26 | Friesian, Japan | [171] | |
| - | 27 | British Friesian | [172] | |
| - | 28 | Czech Republic | [166] | |
| 2 | 4 | Friesian, England | [173] | |
| - | 8 | Friesian, England | [167] | |
| - | 27 | Not reported | [167] | |
| - | 28 | Vietnamese cattle | [174] | |
| 3 | 4 | Limousine, France | [175] | |
| - | 12 | Blond D’Aquitaine, France | [80] | |
| - | 16 | Montbéliarde, France | [176] | |
| - | 27 | Black spotted, Romania | [95] | |
| 4 | 4 | Czech Republic | [167] | |
| - | 8 | Chianina, Italy | [177] | |
| - | 10 | Blonde d’Aquitaine, France | [178] | |
| 5 | 18 | Simmenthal, Hungary | [179] | |
| - | 21 | Japanese Black, Japan | [167] | |
| - | 22 | Polish Red White, Poland | [180] | |
| - | 23 | Brown, Romania | [95] | |
| 6 | 8 | Chianina, Italy | [177,181] | |
| - | 28 | Czech Republic | [166] | |
| 7 | 21 | Japanese Black Cattle, Japan | [182,183] | |
| 8 | 9 | Brown Swiss, Switzerland | [167] | |
| - | 23 | Ukrainian Grey | [167] | |
| 9 | 23 | Blonde d’Aquitaine, France | [184] | |
| 10 | 15 | Pitangueiras, Spain | [185] | |
| 11 | 16 | Simmenthal, Hungary | [186] | |
| - | 21 | Brown, Romania | [95] | |
| - | 22 | Czech Republic | [167] | |
| 12 | 12 | Simmenthal, Germany | [167] | |
| - | 15 | Friesian, Argentina | [167] | |
| 13 | 14 | Friesian, Slovakia | [187] | |
| - | 19 | Marchigiana, Italy | [188] | |
| - | 21 | Friesian, Hungary | [189] | |
| - | 24 | Red &White, Poland. Not reported | [80,187,190] | |
| 14 | 17 | Marchigiana, Italy | [191,192] | |
| - | 19 | Braunvieh, Switzerland | [167] | |
| - | 20 | Simmenthal, Switzerland, USA. Spotted, Romania | [95,193,194,195] | |
| - | 21 | Simmental, Hungary | [167] | |
| - | 24 | Podolian, Italy | [196] | |
| - | 28 | Friesian, USA | [197] | |
| 15 | 25 | Barrosã, Portugal | [198] | |
| 16 | 18 | Barrosã, Portugal | [199] | |
| - | 19 | Marchigiana, Italy | [167] | |
| - | 20 | Simmenthal, Czeck Rep. | [200,201] | |
| - | 21 | RedPied, Czeck Rep. | [167] | |
| 19 | 21 | Friesian, France | [202] | |
| 20 | 20 | Simmenthal, Germany | [167] | |
| 21 | 27 | Blonde d’Aquitaine, France | [203] | |
| 21 | 23 | Maremmana, Italy | [204] | |
| - | 29 | Blonde d’Aquitaine, France | [80] | |
| 24 | 27 | Friesian, Egypt | [167] | |
| 25 | 27 | Alpine Grey, Italy | [139] | |
| 26 | 29 | Alpine Grey, Italy | [139,181,205] | |
| 27 | 29 | Guernsey, Canada | [206] | |
| R. buffalo | 1p | 23 | Ital. Mediterranean, Italy | [207] |
| 1p | 18 | Ital. Mediterranean, Italy | [208] | |
| X | X | Murrah, India | [81] | |
| Sheep | 6 | 24 | (t1) New Zeland Romney, NZ | [209,210] |
| 9 | 10 | (t2) New Zeland Romney, NZ | [210,211] | |
| 7 | 25 | (t3) New Zeland Romney, New Zeland | [210,211] | |
| 5 | 8 | (t4) New Zeland Romney, New Zeland | [212] | |
| 8 | 22 | (t5) New Zeland Romney, New Zeland | [212] | |
| 1 | 20 | Undefined Race, Germany | [213] | |
| 8 | 11 | Churra da Terra Quente, Portugal | [214] | |
| Goat | 2 | 13 | Undefined Race, France | [215] |
| 3 | 7 | - | [161] | |
| 5 | 15 | Saanen, Scotland. Saanen, Brazil | [216,217] | |
| 6 | 17 | Saanen, Switzerland. Saanen, Germany | [218,219] | |
| 6 | 15 | Saanen, Italy. Saanen, France. Saanen, Brazil | [220,221,222] | |
| 10 | 12 | Malaguena, Spain | [223] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iannuzzi, A.; Parma, P.; Iannuzzi, L. Chromosome Abnormalities and Fertility in Domestic Bovids: A Review. Animals 2021, 11, 802. https://doi.org/10.3390/ani11030802
Iannuzzi A, Parma P, Iannuzzi L. Chromosome Abnormalities and Fertility in Domestic Bovids: A Review. Animals. 2021; 11(3):802. https://doi.org/10.3390/ani11030802
Chicago/Turabian StyleIannuzzi, Alessandra, Pietro Parma, and Leopoldo Iannuzzi. 2021. "Chromosome Abnormalities and Fertility in Domestic Bovids: A Review" Animals 11, no. 3: 802. https://doi.org/10.3390/ani11030802
APA StyleIannuzzi, A., Parma, P., & Iannuzzi, L. (2021). Chromosome Abnormalities and Fertility in Domestic Bovids: A Review. Animals, 11(3), 802. https://doi.org/10.3390/ani11030802