
Mammary Gland Structures Are Not Affected by an Increased Growth Rate of Yearling Ewes Post-Weaning but Are Associated with Growth Rates of Singletons

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
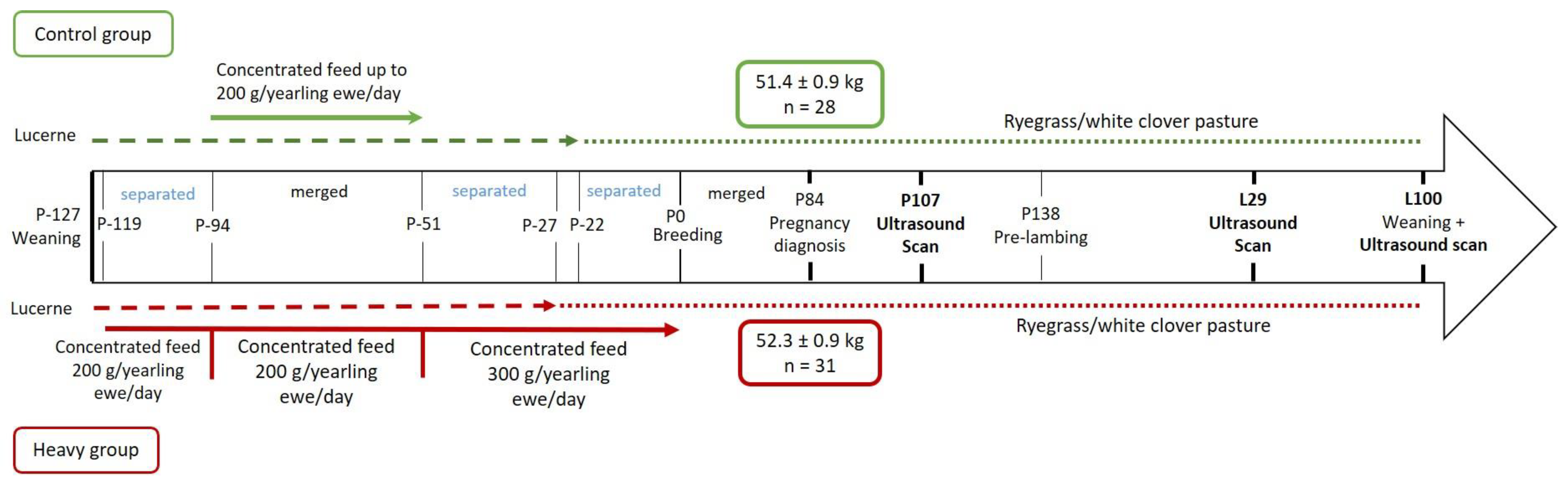
2.1. Experimental Design
2.1.1. Background
2.1.2. Present Study
2.2. Animal Measurements
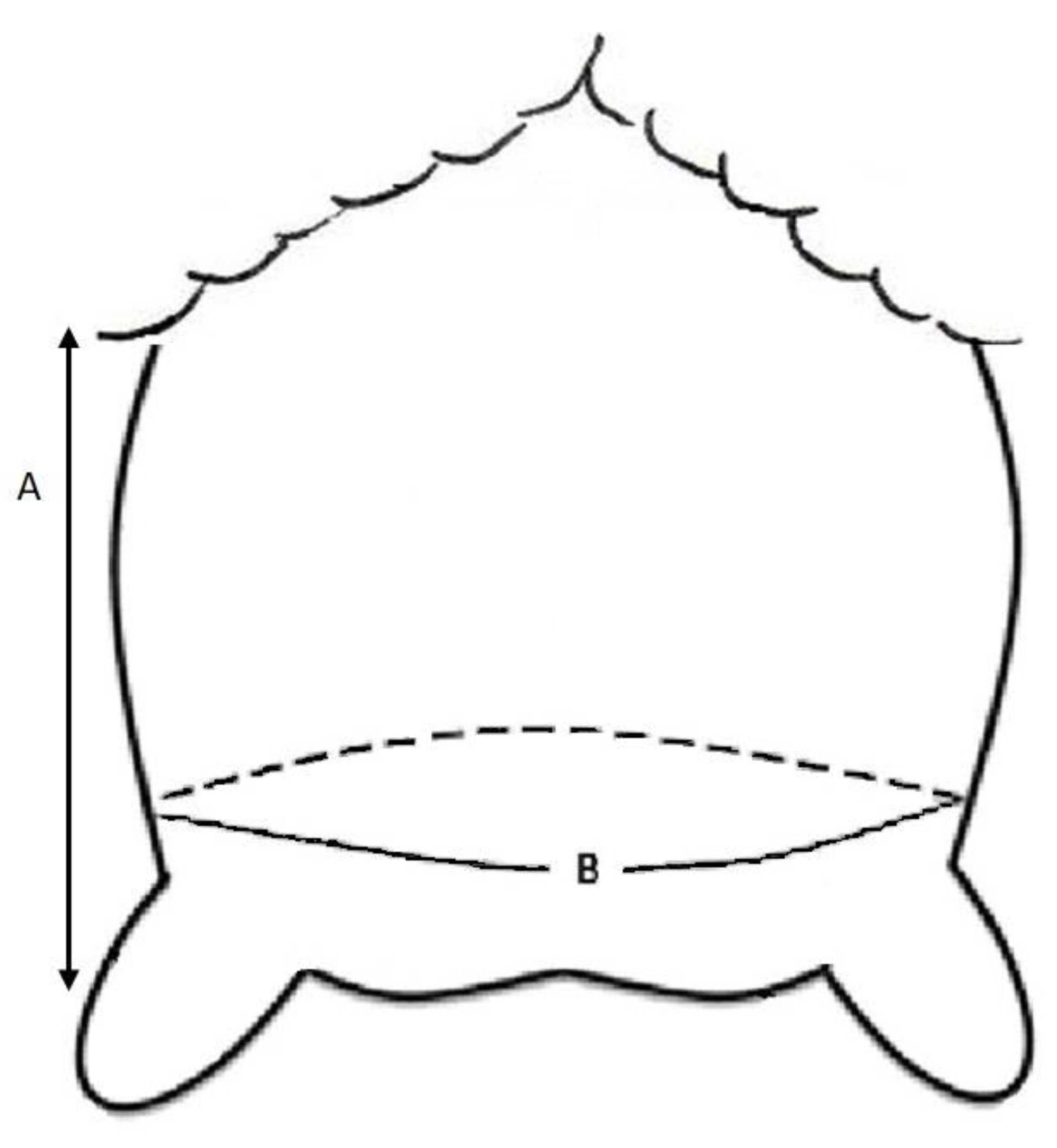
2.2.1. Udder Score and Morphology
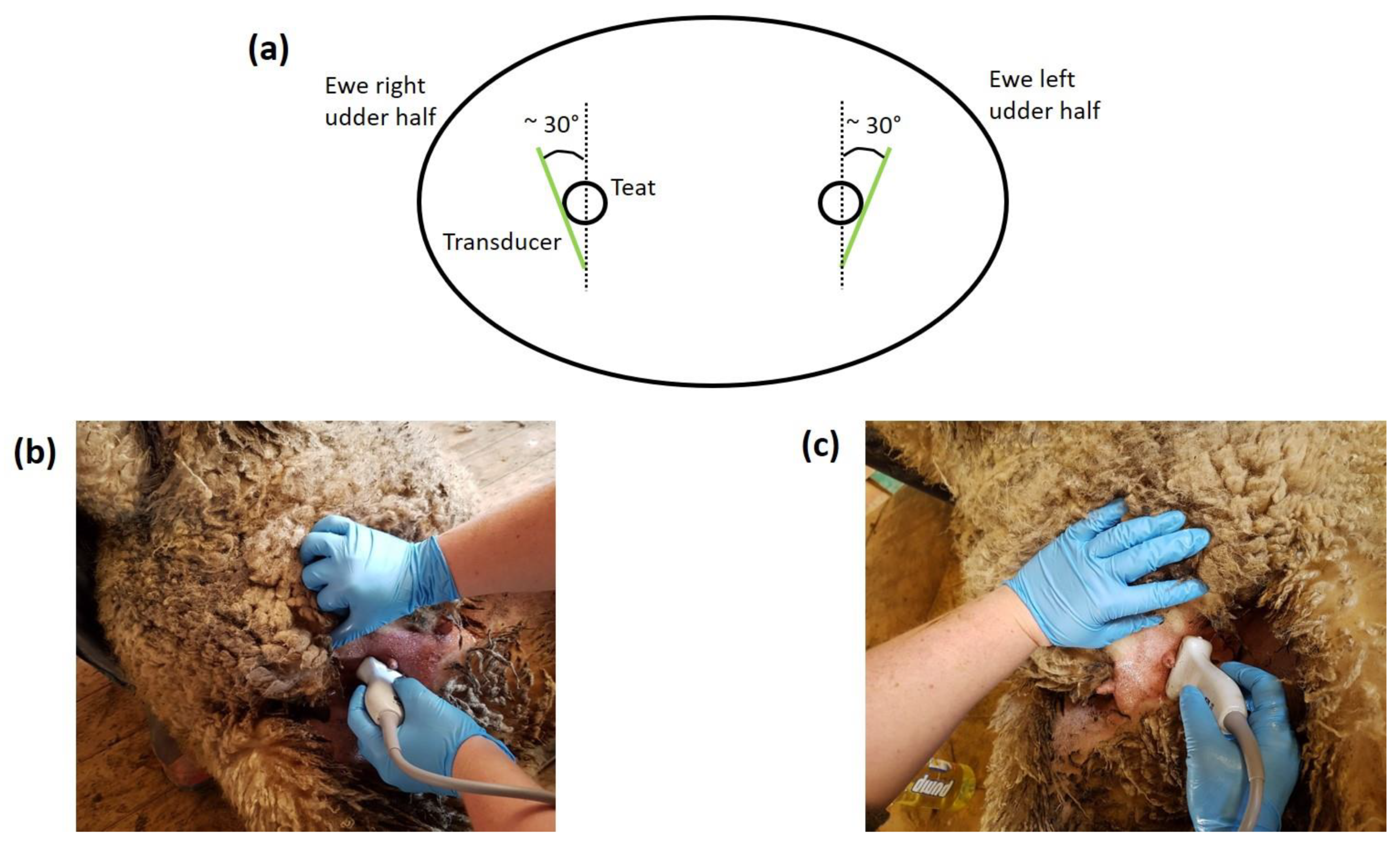
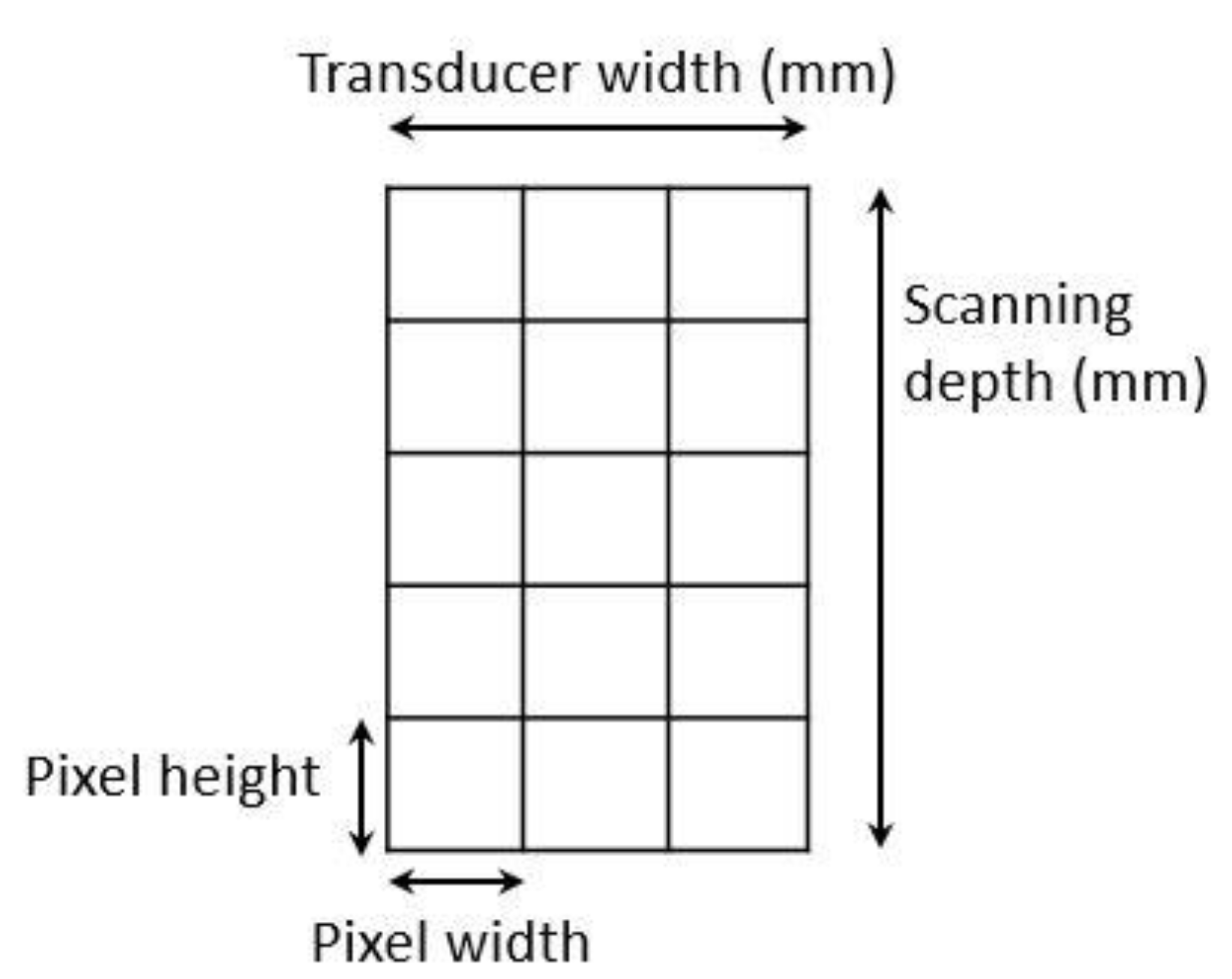
2.2.2. Ultrasound Scanning
2.3. Statistical Analysis
3. Results
3.1. Growth and Live-Weight
3.2. Udder Scores and Morphology
3.3. Ultrasound Measurements
3.4. Correlations between Udder Measurements, Ewe Live-Weight, Ewe BCS and Lamb Growth
3.5. Multiple Regression of Lamb Growth
4. Discussion
4.1. Treatment Effects
4.2. Associations between Udder Measurements and Lamb Growth
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Drawing Templates of Mammary Ultrasound Images in Late Pregnancy, Early Lactation and Weaning in Yearling Ewes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Point and Scanning Depth | Raw Image | Measurement of Udder Structures | Depth (mm) |
|---|---|---|---|
| 113 days of pregnancy (P113) Scanning depth 4.7 cm |  |  | MT generous: 28.9 mm MT conservative: 25.9 mm Gland cistern: 3.3 mm Parenchyma: 10.9 mm Fat Pad: 10.0 mm |
| 109 days of pregnancy (P109) Scanning depth 4.7 cm |  |  | MT generous: 36.8 mm MT conservative: 27.2 mm Gland cistern: 4.7 mm Parenchyma: 15.4 mm Fat Pad: 5.5 mm |
| 107 days of pregnancy (P107) Scanning depth 4.7 cm |  |  | MT generous: 31.7 mm MT conservative: 22.3 mm Gland cistern: 4.5 mm Parenchyma: 8.3 mm Fat Pad: 7.8 mm |
| 107 days of pregnancy (P107) Scanning depth 4.7 cm |  |  | MT generous: 32.2 mm MT conservative: 25.3 mm Gland cistern: 3.6 mm Parenchyma: 12.4 mm Fat Pad: 7.9 mm |
| Time Point and Scanning Depth | Raw Image | Measurement of Udder Structures | Depth (mm) |
|---|---|---|---|
| 32 days of lactation (L32) Scanning depth 7.3 cm |  |  | MT generous: 70.6 mm MT conservative: 67.2 mm Gland cistern: 17.6 mm Parenchyma: 49.6 mm |
| 26 days of lactation (L26) Scanning depth 7.3 cm |  |  | MT generous: 68.9 mm MT conservative: 63.9 mm Gland cistern: 22.8 mm Parenchyma: 41.1 mm |
| 34 days of lactation (L34) Scanning depth 7.3 cm |  |  | MT generous: 70.8 mm MT conservative: 67.4 mm Gland cistern: 14.5 mm Parenchyma: 52.9 mm |
| 26 days of lactation (L26) Scanning depth 7.3 cm |  |  | MT generous: 68.5 mm MT conservative: 65.0 mm Gland cistern: 8.1 mm Parenchyma: 56.9 mm |
| Time Point and Scanning Depth | Raw Image | Measurement of Udder Structures | Depth (mm) |
|---|---|---|---|
| 104 days of lactation (L104) Scanning depth 5.9 cm |  |  | MT generous: 53 mm MT conservative: 50.4 mm Gland cistern: 11.9 mm Parenchyma: 15.9 mm Fat Pad: 19.7 mm |
| 106 days of lactation (L106) Scanning depth 5.9 cm |  |  | MT generous: 54.4 mm MT conservative: 50.1 mm Gland cistern: 10.5 mm Parenchyma: 21.1 mm Fat Pad: 15.3 mm |
| 100 days of lactation (L100) Scanning depth 5.9 cm |  |  | MT generous: 31.8 mm MT conservative: 28.2 mm Gland cistern: 3 mm Parenchyma: 20.1mm Fat Pad: 8 mm |
| 105 days of lactation (L105) Scanning depth 5.9 cm |  |  | MT generous: 48.2 mm MT conservative: 39.5 mm Gland cistern: 4.4 mm Parenchyma: 25.1 mm Fat conservative: 11.9 mm Fat generous: 19 mm |
References
- Dýrmundsson, Ó.R. Puberty and early reproductive performance in sheep. I. Ewe lambs. Commonw. Bur. Anim. Breed. Genet. 1973, 41, 273–289. [Google Scholar]
- Kenyon, P.R.; Thompson, A.N.; Morris, S.T. Breeding ewe lambs successfully to improve lifetime performance. Small Rumin. Res. 2014, 118, 2–15. [Google Scholar] [CrossRef]
- Haslin, E.; Corner-Thomas, R.A.; Kenyon, P.R.; Morris, S.T.; Pettigrew, E.J.; Hickson, R.E.; Blair, H.T. Brief Communication: Impacts of live weight of ewe lambs at mating on their reproductive performance. N. Z. J. Anim. Sci. Prod. 2019, 79, 87–90. [Google Scholar]
- Tolman, B.; McKusick, B.C. The effect of growth rate on mammary gland development in ewe lambs: A review. In Proceedings of the Dairy Sheep Symposium, Eau Claire, WI, USA, 1–3 November 2001. [Google Scholar]
- Villeneuve, L.; Cinq-Mars, D.; Lacasse, P. Effects of restricted feeding of prepubertal ewe lambs on growth performance and mammary gland development. Animal 2010, 4, 944–950. [Google Scholar] [CrossRef] [Green Version]
- McCann, M.A.; Goode, L.; Harvey, R.W.; Caruolo, E.V.; Mann, D.L. Effects of rapid weight gain to puberty on reproduction, mammary development and lactation in ewe lambs. Theriogenology 1989, 32, 55–68. [Google Scholar] [CrossRef]
- Anderson, R.R. Mammary gland growth in sheep. J. Anim. Sci. 1975, 41, 118–123. [Google Scholar] [CrossRef]
- Johnsson, I.D.; Hart, I.C. Pre-pubertal mammogenesis in the sheep 1. The effects of level of nutrition on growth and mammary development in female lambs. Anim. Sci. 1985, 41, 323–332. [Google Scholar] [CrossRef]
- Hovey, R.C.; Mcfadden, T.B.; Akers, R.M. Regulation of mammary gland growth and morphogenesis by the mammary fat pad: A species comparison. J. Mammary Gland. Biol. Neoplasia 1999, 4, 53–68. [Google Scholar] [CrossRef]
- Berryhill, G.E.; Trott, J.E.; Derpinghaus, A.L.; Hovey, R.C. TRIENNAL LACTATION SYMPOSIUM/BOLFA: Dietary regulation of allometric ductal growth in the mammary glands. J. Anim. Sci. 2017, 95, 5664–5674. [Google Scholar] [CrossRef] [Green Version]
- Umberger, S.H.; Goode, L.; Caruolo, E.V.; Harvey, R.W.; Britt, J.H.; Linnerud, A.C. Effects of accelerated growth during rearing on reproduction and lactation in ewes lambing at 13 to 15 months of age. Theriogenology 1985, 23, 555–564. [Google Scholar] [CrossRef]
- Barbagianni, M.S.; Mavrogianni, V.S.; Vasileiou, N.G.C.; Fthenakis, G.C.; Petridis, I.G. Ultrasonographic examination of the udder in sheep. Small Rumin. Res. 2017, 152, 86–99. [Google Scholar] [CrossRef]
- Petridis, I.G.; Gouletsou, P.G.; Barbagianni, M.S.; Amiridis, G.S.; Brozos, C.; Valasi, I.; Fthenakis, G.C. Ultrasonographic findings in the ovine udder during involution. J. Dairy Res. 2014, 81, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Barbagianni, M.S.; Gouletsou, P.G.; Valasi, I.; Petridis, I.G.; Giannenas, I.; Fthenakis, G.C. Ultrasonographic findings in the ovine udder during lactogenesis in healthy ewes or ewes with pregnancy toxaemia. J. Dairy Res. 2015, 82, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Caja, G.; Such, X.; Ruberte, J.; Carretero, A.; Navarro, M. The use of ultrasonography in the study of mammary gland cisterns during lactation in sheep. Proc. Eur. Soc. Anim. Prod. 1999, 95, 91–93. [Google Scholar]
- Rovai, M.; Caja, G.; Such, X. Evaluation of udder cisterns and effects on milk yield of dairy ewes. J. Dairy Sci. 2008, 91, 4622–4629. [Google Scholar] [CrossRef] [Green Version]
- Carta, A.; Casu, S.; Salaris, S. Invited review: Current state of genetic improvement in dairy sheep. J. Dairy Sci. 2009, 92, 5814–5833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pourlis, A. Ovine mammary morphology and associations with milk production, milkability and animal selection. Small Rumin. Res. 2020, 184. [Google Scholar] [CrossRef]
- Ruberte, J.; Carretero, A.; Fernandez, M.; Navarro, M.; Caja, G.; Kirchner, F.; Such, X. Ultrasound mammography in the lactating ewe and its correspondence to anatomical section. Small Rumin. Res. 1994, 13, 199–204. [Google Scholar] [CrossRef]
- Haslin, E.; Corner-Thomas, R.A.; Kenyon, P.R.; Morris, S.T.; Blair, H.T. Effects of heavier live weight of ewe lambs at mating on fertility, lambing percentage, subsequent live weight and the performance of their progeny. N. Z. J. Agric. Res. 2020, 1–15. [Google Scholar] [CrossRef]
- Corner, R.A.; Mulvaney, F.J.; Morris, S.T.; West, D.M.; Morel, P.C.H.; Kenyon, P.R. A comparison of the reproductive performance of ewe lambs and mature ewes. Small Rumin. Res. 2013, 114, 126–133. [Google Scholar] [CrossRef]
- Nowak, R.; Keller, M.; Lévy, F. Mother-Young Relationships in Sheep: A Model for a Multidisciplinary Approach of the Study of Attachment in Mammals. J. Neuroendocrinol. 2011, 23, 1042–1063. [Google Scholar] [CrossRef]
- Griffiths, K.J.; Ridler, A.L.; Compton, C.W.R.; Corner-Thomas, R.A.; Kenyon, P.R. Investigating associations between lamb survival to weaning and dam udder and teat scores. N. Z. Vet. J. 2019, 67, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Sezenler, T.; Ceyhan, A.; Yüksel, M.A.; Önaldı, A.; Yıldırır, M. Effect of parity and type of lambing on performance and udder traits of Bandirma ewes. Indian J. Anim. Sci. 2016, 86, 575–577. [Google Scholar]
- Ayadi, M.; Such, X.; Ezzehizi, N.; Zouari, M.; Najar, T.; M’Rad, M.B.; Casals, R. Relationship between mammary morphology traits and milk yield of Sicilo-Sarde dairy sheep in Tunisia. Small Rumin. Res. 2011, 96, 41–45. [Google Scholar] [CrossRef]
- Albino, R.L.; Marcondes, M.I.; Akers, R.M.; Detmann, E.; Carvalho, B.C.; Silva, T.E. Mammary gland development of dairy heifers fed diets containing increasing levels of metabolisable protein: Metabolisable energy. J. Dairy Res. 2015, 82, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Albino, R.L.; Guimarães, S.E.F.; Daniels, K.M.; Fontes, M.M.S.; Machado, A.F.; Dos Santos, G.B.; Marcondes, M.I. Mammary gland ultrasonography to evaluate mammary parenchymal composition in prepubertal heifers. J. Dairy Sci. 2017, 100, 1588–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, T.; Rasband, W.S. ImageJ User Guide-IJ 1.46. Available online: Imagej.nih.gov/ij/docs/guide/ (accessed on 22 November 2019).
- Abràmoff, M.D.; Magalhães, P.J.; Ram, S.J. Image processing with ImageJ. Biophotonics Int. 2004, 11, 36–42. [Google Scholar]
- Molenaar, A.J.; Maclean, P.H.; Gilmour, M.L.; Draganova, I.G.; Symes, C.W.; Margerison, J.K.; McMahon, C.D. Effect of whole-milk allowance on liveweight gain and growth of parenchyma and fat pads in the mammary glands of dairy heifers at weaning. J. Dairy Sci. 2020, 103, 5061–5069. [Google Scholar] [CrossRef] [PubMed]
- Ojala, T.; Pietikainen, M.; Maenpaa, T. Multiresolution gray-scale and rotation invariant texture classification with local binary patterns. IEEE Trans. Pattern Anal. Mach. Intell. 2002, 24, 971–987. [Google Scholar] [CrossRef]
- Strzetelski, J.; Bilik, K.; Niwińska, B.; Skrzyński, G.; Łuczyńska, E. Ultrasound evaluation of the mammary gland structure in preparturient heifers vs. performance of first calvers. J. Anim. Feed Sci. 2004, 13, 7–10. [Google Scholar] [CrossRef]
- Pettigrew, E.; Hickson, R.; Morris, S.; Kenyon, P.; Corner-Thomas, R.; Haslin, E.; Blair, H. The Effect of Age of Dam and Birth Rank on the Reproductive Performance of Ewes as One-and Two-Year-Olds. Animals 2021, 11, 770. [Google Scholar] [CrossRef]
- Dohoo, I.R.; Martin, W.; Stryhn, H.E. Veterinary Epidemiologic Research; AVC Inc.: Charlottetown, EI, Canada, 2003. [Google Scholar]
- Haslin, E.; Corner-Thomas, R.A.; Kenyon, P.R.; Morris, S.T.; Blair, H.T. Impacts of a heavier live weight at breeding on the morphology of mammary glands of non-dairy ewe lambs [Abstract]. Anim. Prod. Sci. 2021, 61, 165. [Google Scholar]
- Makovicky, P.; Milerski, M.; Margetín, M.; Makovicky, P.; Nagy, M. Genetic parameters for the size of udder cisterns in ewes diagnosed by ultrasonography among breeds: Improved Valachian, Tsigai, Lacaune and their crosses. Arch. Zootec. 2015, 64, 403–408. [Google Scholar]
- Jena, M.K.; Jaswal, S.; Kumar, S.; Mohanty, A.K. Molecular mechanism of mammary gland involution: An update. Dev. Biol. 2018, 445, 145–155. [Google Scholar] [CrossRef]
- Akers, R.M. Overview of Mammary Development. In Lactation and the Mammary Gland; Akers, R.M., Ed.; Blackwell Publishing Company: Ames, IA, USA, 2002; pp. 3–44. [Google Scholar]
- Colville, T. Chapter 18. Pregnancy, Development, and Lactation. In Clinical Anatomy and Physiology for Veterinary Technicians, 2nd ed.; Colville, T., Bassert, J.M., Eds.; Elsevier: St Louis, MO, USA, 2007; pp. 405–413. [Google Scholar]
- Boutinaud, M.; Guinard-Flament, J. The number and activity of mammary epithelial cells, determining factors for milk production. Reprod. Nutr. Dev. 2004, 44, 499–508. [Google Scholar] [CrossRef]
- Capuco, A.V.; Wood, D.L.; Baldwin, R.; Mcleod, K.; Paape, M.J. Mammary cell number, proliferation, and apoptosis during a bovine lactation: Relation to milk production and effect of bST. J. Dairy Sci. 2001, 84, 2177–2187. [Google Scholar] [CrossRef]
- Castillo, V.; Such, X.; Caja, G.; Salama, A.A.K.; Albanell, E.; Casals, R. Changes in alveolar and cisternal compartments induced by milking interval in the udder of dairy ewes. J. Dairy Sci. 2008, 91, 3403–3411. [Google Scholar] [CrossRef]





| Score | Description |
|---|---|
| Udder palpation a | |
| 1 | Diffuse soft consistency |
| 2 | Diffuse firm consistency |
| 3 | Soft consistency with nodule(s)—lumps or grainy texture |
| 4 | Firm consistency with nodule(s)—lumps or grainy texture |
| 5 | Diffuse hard consistency |
| Teat palpation a | |
| 1 | Soft consistency |
| 2 | Thickened teat orifice |
| 3 | Hard consistency |
| 4 | Teat orifice obstruction |
| Parameters | Pregnancy (P107) | Pregnancy (P138) | Early Lactation (L29) | Weaning (L100) |
|---|---|---|---|---|
| Ewe live-weight (kg) | ||||
| Control (n = 24) | 52.3 ± 0.81 a | 58.5 ± 0.87 b | 61.7 ± 1.21 c | 62.0 ± 1.26 c |
| Heavy (n = 24) | 52.4 ± 0.81 a | 57.8 ± 0.87 b | 60.8 ± 1.21 c | 60.2 ± 1.26 c |
| Ewe BCS | ||||
| Control (n = 24) | 2.77 (2.64–2.90) b | 2.69 (2.53–2.85) a | 2.65 (2.48–2.82) a | |
| Heavy (n = 24) | 2.69 (2.58–2.80) b | 2.46 (2.30–2.63) a | 2.42 (2.24–2.60) a | |
| Lamb live-weight (kg) | ||||
| Control (n = 24) | 15.0 ± 0.52 a | 29.9 ± 0.68 b | ||
| Heavy (n = 24) | 15.1 ± 0.52 a | 29.5 ± 0.68 b |
| Descriptor | Pregnancy (P107) | Early Lactation (L29) | Weaning (L100) | p Values (Time) | p Values (Treatment) | p Values (Treatment × Time) |
|---|---|---|---|---|---|---|
| Control group (n = 24) | ||||||
| GC (mm) | 3.57 ± 0.88 a | 16.5 ± 0.88 c | 9.62 ± 0.87 b | <0.001 | 0.082 | 0.472 |
| PAR (mm) | 10.2 ± 0.41 a | 46.1 ± 1.32 c | 18.9 ± 0.74 b | <0.001 | 0.600 | 0.002 |
| FP (mm) | 10.3 ± 0.45 a | - 1 | RS 14.7 ± 1.36 *,b and LS 19.7 ± 1.28 *,b | <0.001 | 0.886 | 0.970 |
| MTc (mm) | 25.6 ± 0.56 a | 63.1 ± 1.06 c | 44.2 ± 0.98 b | <0.001 | 0.091 | 0.063 |
| MTg (mm) | 32.2 ± 0.94 a | 65.5 ± 0.93 c | 49.4 ± 0.96 b | <0.001 | 0.151 | 0.519 |
| ROI (grey scale value) | 88.0 ± 2.10 | 93.6 ± 2.79 | 88.3 ± 2.23 | 0.390 | 0.544 | 0.401 |
| Heavy group (n = 24) | ||||||
| GC (mm) | 4.37 ± 0.88 a | 18.2 ± 0.87 c | 12.0 ± 0.87 b | <0.001 | 0.082 | 0.472 |
| PAR (mm) | 8.81 ± 0.41 a | 45.8 ± 1.31 c | 21.6 ± 0.73 b | <0.001 | 0.600 | 0.002 |
| FP (mm) | 10.4 ± 0.45 a | - 1 | RS 16.2 ± 1.43 *,b and LS 18.7 ± 1.31 *,b | <0.001 | 0.886 | 0.970 |
| MTc (mm) | 25.2 ± 0.56 a | 63.5 ± 1.06 c | 47.7 ± 0.94 b | <0.001 | 0.091 | 0.063 |
| MTg (mm) | 32.3 ± 0.93 a | 67.0 ± 0.93 c | 51.4 ± 0.93 b | <0.001 | 0.151 | 0.519 |
| ROI (grey scale value) | 91.7 ± 2.10 | 91.5 ± 2.78 | 90.7 ± 2.22 | 0.390 | 0.544 | 0.401 |
| Descriptor | UC | UH | GC | PAR | FP | Ewe LW (P107) | Ewe BCS (P138) | Birth to L100 | Birth to L29 | L29 to L100 |
|---|---|---|---|---|---|---|---|---|---|---|
| UV | 0.738 *** | 0.910 *** | 0.193 | 0.243 | −0.213 | −0.016 | −0.038 | 0.041 | −0.163 | 0.175 |
| UC | 0.408 ** | 0.251 | 0.315 * | −0.229 | −0.062 | −0.114 | −0.041 | −0.056 | −0.009 | |
| UH | 0.109 | 0.175 | −0.168 | −0.013 | −0.007 | 0.052 | −0.176 | 0.200 | ||
| GC | 0.040 | −0.448 ** | −0.026 | −0.153 | −0.110 | −0.156 | −0.0009 | |||
| PAR | −0.625 *** | −0.369 ** | −0.314 * | 0.373 ** | 0.091 | 0.331 * | ||||
| FP | 0.257 | 0.373 ** | −0.374 ** | −0.177 | −0.246 | |||||
| Ewe LW (P107) | 0.413 ** | 0.005 | 0.126 | −0.115 | ||||||
| Ewe BCS (P138) | −0.017 | 0.019 | −0.030 | |||||||
| Birth to L100 | 0.480 *** | 0.649 *** | ||||||||
| Birth to L29 | −0.351 * |
| Descriptor | UH | UC | GC | PAR | Ewe LW (L29) | Ewe BCS (L29) | Birth to L100 | Birth to L29 | L29 to L100 |
|---|---|---|---|---|---|---|---|---|---|
| UV | 0.648 *** | 0.649 *** | 0.006 | −0.012 | 0.254 | −0.144 | −0.100 | −0.010 | −0.085 |
| UH | −0.100 | 0.170 | −0.145 | 0.056 | −0.097 | −0.052 | 0.007 | −0.043 | |
| UC | −0.160 | 0.128 | 0.289 * | −0.052 | −0.003 | 0.008 | −0.009 | ||
| GC | −0.976 *** | 0.011 | −0.017 | 0.298 * | 0.284 * | 0.071 | |||
| PAR | 0.032 | 0.029 | −0.324 * | −0.272 | −0.105 | ||||
| Ewe LW (L29) | 0.500 *** | 0.180 | 0.113 | 0.108 | |||||
| Ewe BCS (L29) | 0.171 | 0.183 | 0.093 |
| Descriptor | UH | UC | GC | PAR | FP Right | FP Left | Ewe LW (L100) | Ewe BCS (L100) | Birth to L100 | Birth to L29 | L29 to L100 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| UV | 0.771 *** | 0.876 *** | 0.022 | −0.181 | 0.161 | 0.317 * | −0.091 | −0.112 | 0.138 | −0.088 | 0.226 |
| UH | 0.394 ** | 0.150 | −0.152 | 0.111 | 0.309 * | −0.206 | 0.007 | 0.006 | −0.137 | 0.135 | |
| UC | −0.021 | −0.121 | 0.153 | 0.200 | 0.0003 | −0.101 | 0.204 | −0.058 | 0.264 | ||
| GC | −0.291 * | −0.226 | −0.132 | 0.009 | −0.068 | 0.111 | 0.194 | −0.045 | |||
| PAR | −0.210 | −0.333 * | 0.144 | −0.062 | 0.106 | −0.105 | 0.216 | ||||
| FP Right | −0.230 | −0.214 | −0.067 | 0.051 | 0.344 * | −0.233 | |||||
| FP Left | −0.151 | −0.026 | 0.122 | −0.280 | 0.379 * | ||||||
| Ewe LW (L100) | 0.664 *** | 0.040 | 0.072 | −0.017 | |||||||
| Ewe BCS (L100) | −0.202 | −0.171 | −0.068 |
| Descriptor | Minimum | 10th Percentile | Mean | 90th Percentile | Maximum |
|---|---|---|---|---|---|
| GC (mm) | |||||
| P107 | 1.9 | 2.4 | 4.0 | 7.1 | 8.3 |
| L29 | 4.2 | 8.5 | 17.2 | 23.8 | 30.7 |
| L100 | 4.3 | 5.4 | 10.8 | 16.2 | 25.8 |
| PAR (mm) | |||||
| P107 | 5.7 | 6.8 | 9.5 | 12.1 | 17.2 |
| L29 | 28.8 | 37.2 | 46.0 | 56.6 | 60.6 |
| L100 | 12.3 | 16.6 | 20.2 | 26.3 | 29.9 |
| FP (mm) | |||||
| P107 | 6.0 | 8.2 | 10.4 | 13.2 | 15.8 |
| L100-Left | 2.3 | 10.1 | 19.2 | 27.1 | 30.9 |
| L100-Right | 4.0 | 7.5 | 15.4 | 24.5 | 28.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haslin, E.; Corner-Thomas, R.A.; Kenyon, P.R.; Molenaar, A.J.; Morris, S.T.; Blair, H.T. Mammary Gland Structures Are Not Affected by an Increased Growth Rate of Yearling Ewes Post-Weaning but Are Associated with Growth Rates of Singletons. Animals 2021, 11, 884. https://doi.org/10.3390/ani11030884
Haslin E, Corner-Thomas RA, Kenyon PR, Molenaar AJ, Morris ST, Blair HT. Mammary Gland Structures Are Not Affected by an Increased Growth Rate of Yearling Ewes Post-Weaning but Are Associated with Growth Rates of Singletons. Animals. 2021; 11(3):884. https://doi.org/10.3390/ani11030884
Chicago/Turabian StyleHaslin, Emmanuelle, Rene A. Corner-Thomas, Paul R. Kenyon, Adrian J. Molenaar, Stephen T. Morris, and Hugh T. Blair. 2021. "Mammary Gland Structures Are Not Affected by an Increased Growth Rate of Yearling Ewes Post-Weaning but Are Associated with Growth Rates of Singletons" Animals 11, no. 3: 884. https://doi.org/10.3390/ani11030884
APA StyleHaslin, E., Corner-Thomas, R. A., Kenyon, P. R., Molenaar, A. J., Morris, S. T., & Blair, H. T. (2021). Mammary Gland Structures Are Not Affected by an Increased Growth Rate of Yearling Ewes Post-Weaning but Are Associated with Growth Rates of Singletons. Animals, 11(3), 884. https://doi.org/10.3390/ani11030884