
Genetic Analysis of Major Production and Reproduction Traits of Korean Duroc, Landrace and Yorkshire Pigs


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Phenotypes
2.2. Animal Pedigree
2.3. Statistical Analyses
3. Results
3.1. Descriptive Statistics
3.2. Variance Components and Genetic Parameter Estimates
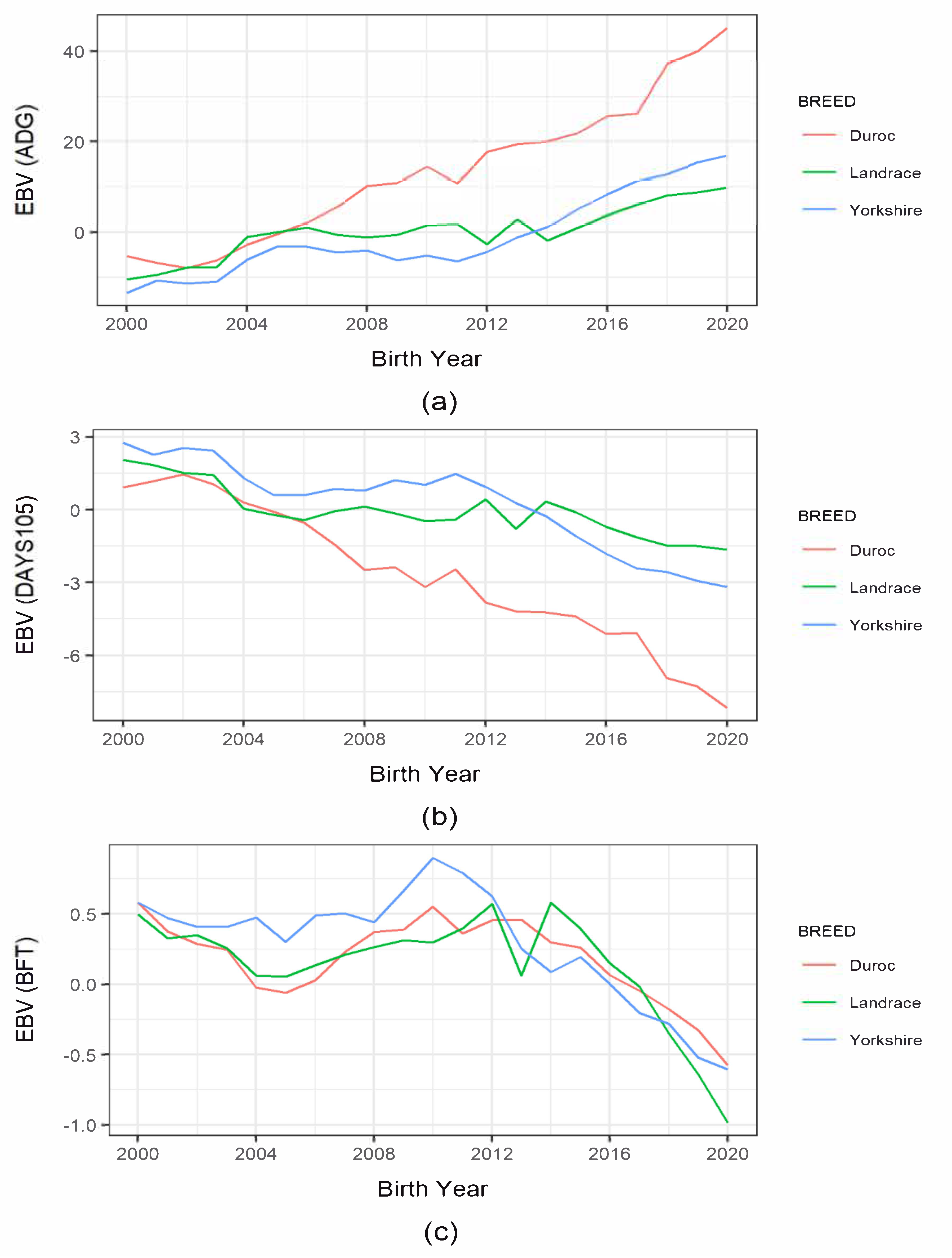
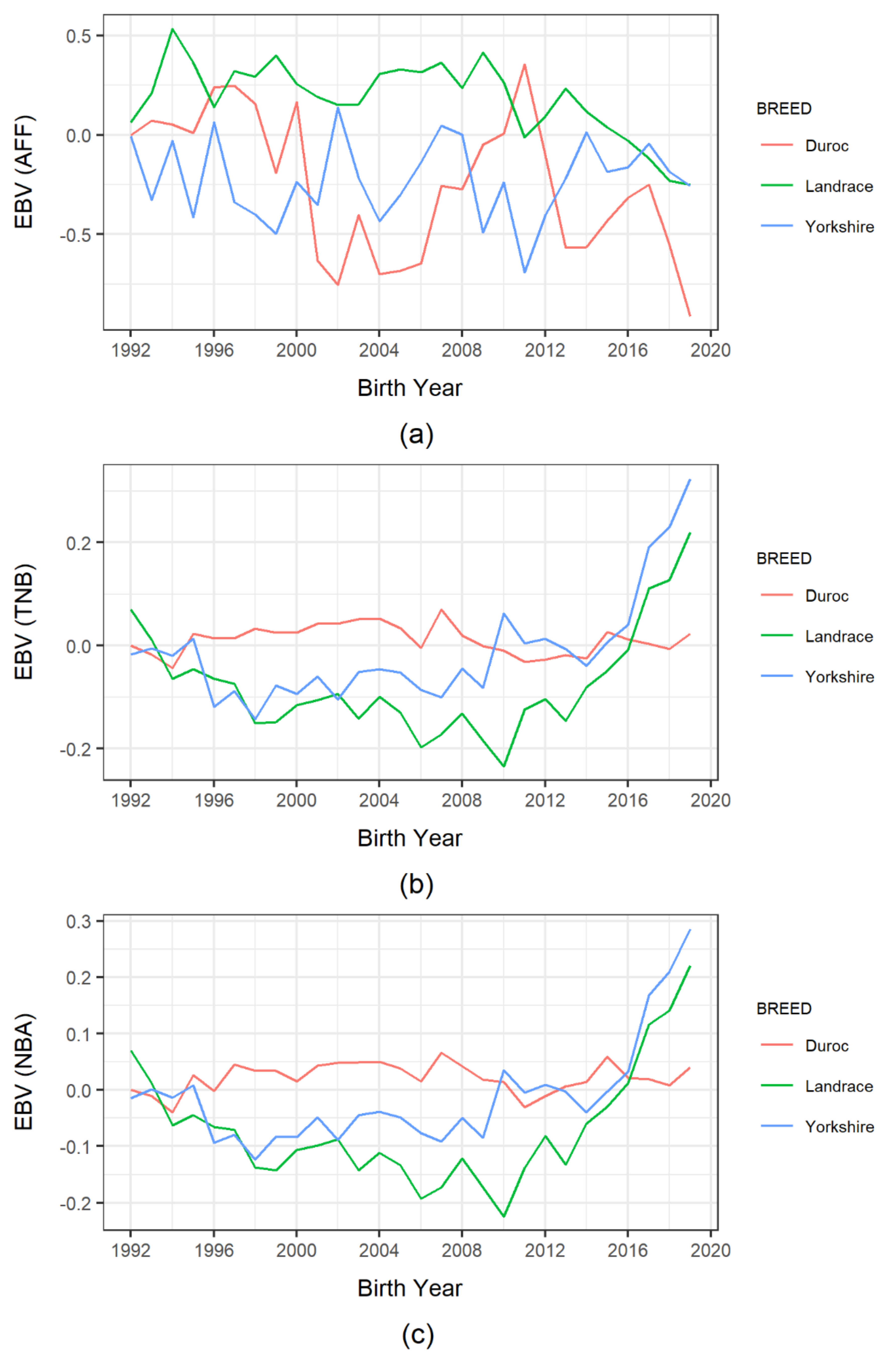
3.3. Genetic Trends of Production and Reproduction Traits
4. Discussion
4.1. Heritability of Production Traits
4.2. Genetic Correlations among Production Traits
4.3. Heritability of Reproduction Traits
4.4. Genetic Correlations among Reproduction Traits
4.5. Genetic Trends in Production and Reproduction Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiao, S.; Maltecca, C.; Gray, K.A.; Cassady, J.P. Feed intake, average daily gain, feed efficiency, and real-time ultrasound traits in Duroc pigs: II. genomewide association. J. Anim. Sci. 2014, 92, 2846–2860. [Google Scholar] [CrossRef] [PubMed]
- Edwards, W.M.; van der Sluis, G.T.; Stevermer, E.J. Determinants of profit- ability in farrow-to-finish swine production. North Cent. J. Agric. Econ. 1989, 11, 17–25. [Google Scholar]
- Klimas, R.; Klimiene, A. Genetic trend and relationship of meatiness traits of different breed pigs raised in lithuania. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 2009, 63, 66–69. [Google Scholar] [CrossRef] [Green Version]
- Merks, J.W.M.; Mathur, P.K.; Knol, E.F. New phenotypes for new breeding goals in pigs. Animal 2012, 6, 535–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoque, M.A.; Suzuki, K.; Kadowaki, H.; Shibata, T.; Oikawa, T. Genetic parameters for feed efficiency traits and their relationships with growth and carcass traits in Duroc pigs. J. Anim. Breed. Genet. 2007, 124, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Stalder, K.J.; Lacy, R.C.; Cross, T.L.; Conatser, G.E. Financial impact of average parity of culled females in a breed-to-wean swine operation using replacement gilt net present value analysis. J. Swine Health Prod. 2003, 11, 69–74. [Google Scholar]
- Tummaruk, P.; Lundeheim, N.; Einarsson, S.; Dalin, A.M. Reproductive performance of purebred Hampshire sows in Sweden. Livest. Prod. Sci. 2001, 68, 67–77. [Google Scholar] [CrossRef]
- Serenius, T.; Stalder, K.J. Length of productive life of crossbred sows is affected by farm management, leg conformation, sow’s own prolificacy, sow’s origin parity and genetics. Animal 2007, 1, 745–750. [Google Scholar] [CrossRef] [Green Version]
- Sobczyńska, M.; Blicharski, T.; Tyra, M. Relationships between longevity, lifetime productivity, carcass traits and conformation in Polish maternal pig breeds. J. Anim. Breed. Genet. 2013, 130, 361–371. [Google Scholar] [CrossRef]
- Noppibool, U.; Elzo, M.A.; Koonawootrittriron, S.; Suwanasopee, T. Estimation of Genetic Parameters and Trends for Length of Productive Life and Lifetime Production Traits in a Commercial Landrace and Yorkshire Swine Population in Northern Thailand. Asian Australas. J. Anim. Sci. 2016, 29, 1222–1228. [Google Scholar] [CrossRef] [Green Version]
- Iida, R.; Piñeiro, C.; Koketsu, Y. High lifetime and reproductive performance of sows on southern European Union commercial farms can be predicted by high numbers of pigs born alive in parity one. J. Anim. Sci. 2015, 93, 2501–2508. [Google Scholar] [CrossRef]
- Sasaki, Y.; Koketsu, Y. Sows having high lifetime efficiency and high longevity associated with herd productivity in commercial herds. Livest. Sci. 2008, 118, 140–146. [Google Scholar] [CrossRef]
- Choy, Y.H.; Mahboob, A.; Cho, C.I.; Choi, J.G.; Choi, I.S.; Choi, T.J.; Cho, K.H.; Park, B.H. Genetic parameters of pre-adjusted body weight growth and ultrasound measures of body tissue development in three seedstock pig breed populations in Korea. Asian Australas. J. Anim. Sci. 2015, 28, 1696–1702. [Google Scholar] [CrossRef] [Green Version]
- Lopez, B.I.M.; Song, C.; Seo, K. Genetic parameters and trends for production traits and their relationship with litter traits in Landrace and Yorkshire pigs. Anim. Sci. J. 2018, 89, 1381–1388. [Google Scholar] [CrossRef]
- Lopez, B.I.; Viterbo, V.; Song, C.W.; Seo, K.S. Estimation of genetic parameters and accuracy of genomic prediction for production traits in duroc pigs. Czech J. Anim. Sci. 2019, 64, 160–165. [Google Scholar] [CrossRef] [Green Version]
- Falconer, D.S. Heritability. In Introduction to Quantitative Genetics; Longman Scientific and Technical: New York, NY, USA, 1989; p. 164. [Google Scholar]
- Wray, N.; Visscher, P. Estimating trait heritability. Nat. Educ. 2008, 1, 29. [Google Scholar]
- Misztal, I.; Tsuruta, S.; Lourenco, D.; Aguilar, I.; Legarra, A.; Vitezica, Z.Z. Manual for BLUPF90 Family of Programs. Available online: https://nce.ads.uga.edu/wiki/lib/exe/fetch.%0Aphp?media=blupf90_all2.pdf (accessed on 14 April 2021).
- Houle, D.; Meyer, K. Estimating sampling error of evolutionary statistics based on genetic covariance matrices using maximum likelihood. J. Evol. Biol. 2015, 28, 1542–1549. [Google Scholar] [CrossRef]
- Houle, D. Comparing evolvability and variability of quantitative traits. Genetics 1992, 130, 195–204. [Google Scholar] [CrossRef]
- Clutter, A.C. Genetics of performance traits. In The Genetics of the Pigs; Rothschild, M.F., Ruvinsky, A., Eds.; CAB International: Wallingford, UK, 2011; pp. 325–354. [Google Scholar]
- Su, G.; Christensen, O.F.; Ostersen, T.; Henryon, M.; Lund, M.S. Estimating Additive and Non-Additive Genetic Variances and Predicting Genetic Merits Using Genome-Wide Dense Single Nucleotide Polymorphism Markers. PLoS ONE 2012, 7, e0045293. [Google Scholar] [CrossRef]
- Sirichokchatchawan, T.; Imboonta, N. Genetic parameters for residual feed intake, feed efficiency and average daily gain in landrace pigs. Thai J. Vet. Med. 2015, 45, 543–549. [Google Scholar]
- Suzuki, K.; Irie, M.; Kadowaki, H.; Shibata, T.; Kumagai, M.; Nishida, A. Genetic parameter estimates of meat quality traits in Duroc pigs selected for average daily gain, longissimus muscle area, backfat thickness, and intramuscular fat content. J. Anim. Sci. 2005, 83, 2058–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoque, M.A.; Kadowaki, H.; Shibata, T.; Suzuki, K. Maternal and direct genetic parameters for production traits and maternal correlations among production and feed efficiency traits in duroc pigs. Asian Australas. J. Anim. Sci. 2008, 21, 961–966. [Google Scholar] [CrossRef]
- Hoque, M.A.; Kadowaki, H.; Shibata, T.; Oikawa, T.; Suzuki, K. Genetic parameters for measures of residual feed intake and growth traits in seven generations of Duroc pigs. Livest. Sci. 2009, 121, 45–49. [Google Scholar] [CrossRef]
- Willson, H.E.; Rojas de Oliveira, H.; Schinckel, A.P.; Grossi, D.; Brito, L.F. Estimation of Genetic Parameters for Pork Quality, Novel Carcass, Primal-Cut and Growth Traits in Duroc Pigs. Animals 2020, 10, 779. [Google Scholar] [CrossRef]
- Miar, Y.; Plastow, G.; Bruce, H.; Moore, S.; Manafiazar, G.; Kemp, R.; Charagu, P.; Huisman, A.; Van Haandel, B.; Zhang, C.; et al. Genetic and phenotypic correlations between performance traits with meat quality and carcass characteristics in commercial crossbred pigs. PLoS ONE 2014, 9, e0110105. [Google Scholar] [CrossRef]
- Koots, K.R.; Gibson, J.P. How precise are genetic correlation estimates? In Proceedings of the 5th World Congress on Genetics Applied to Livestock Production, University of, Guelph, ON, Canada, 7–12 August 1994; Volume 18, pp. 353–360. [Google Scholar]
- Choi, J.G.; Cho, C.I.; Choi, I.S.; Lee, S.S.; Choi, T.J.; Cho, K.H.; Park, B.H.; Choy, Y.H. Genetic parameter estimation in seedstock swine population for growth performances. Asian Australas. J. Anim. Sci. 2013, 26, 470–475. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.I.; Sohn, Y.G.; Jung, J.H.; Park, Y.I. Genetic parameter estimates for backfat thickness at three different sites and growth rate in swine. Asian Australas. J. Anim. Sci. 2004, 17, 305–308. [Google Scholar] [CrossRef]
- Banville, M.; Riquet, J.; Bahon, D.; Sourdioux, M.; Canario, L. Genetic parameters for litter size, piglet growth and sow’s early growth and body composition in the Chinese-European line Tai Zumu. J. Anim. Breed. Genet. 2015, 132, 328–337. [Google Scholar] [CrossRef]
- Ito, T.; Fukawa, K.; Kamikawa, M.; Nikaidou, S.; Taniguchi, M.; Arakawa, A.; Tanaka, G.; Mikawa, S.; Furukawa, T.; Hirose, K. Effects of correcting missing daily feed intake values on the genetic parameters and estimated breeding values for feeding traits in pigs. Anim. Sci. J. 2018, 89, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Dube, B.; Mulugeta, S.D.; Dzama, K. Genetic relationship between growth and carcass traits in Large White pigs. S. Afr. J. Anim. Sci. 2013, 43, 482–492. [Google Scholar] [CrossRef] [Green Version]
- Herrera-Cáceres, W.; Sánchez, J.P. Selection for feed efficiency using the social effects animal model in growing Duroc pigs: Evaluation by simulation. Genet. Sel. Evol. 2020, 52, 1–10. [Google Scholar] [CrossRef]
- Saintilan, R.; Mérour, I.; Brossard, L.; Tribout, T.; Dourmad, J.Y.; Sellier, P.; Bidanel, J.; van Milgen, J.; Gilbert, H. Genetics of residual feed intake in growing pigs: Relationships with production traits, and nitrogen and phosphorus excretion traits. J. Anim. Sci. 2013, 91, 2542–2554. [Google Scholar] [CrossRef] [Green Version]
- Ohnishi, C.; Satoh, M. Estimation of genetic parameters for performance and body measurement traits in Duroc pigs selected for average daily gain, loin muscle area, and backfat thickness. Livest. Sci. 2018, 214, 161–166. [Google Scholar] [CrossRef]
- Akanno, E.C.; Schenkel, F.S.; Quinton, V.M.; Friendship, R.M.; Robinson, J.A.B. Meta-analysis of genetic parameter estimates for reproduction, growth and carcass traits of pigs in the tropics. Livest. Sci. 2013, 152, 101–113. [Google Scholar] [CrossRef]
- Chang, H.L.; Lai, Y.Y.; Wu, M.C.; Sasaki, O. Genetic correlations between male reproductive traits and growth traits in growth performance tested Duroc, Landrace and Yorkshire breed boars. Anim. Sci. J. 2017, 88, 1258–1268. [Google Scholar] [CrossRef]
- Kerssen, N.; Ducro, B.J.; Hermesch, S. Genetic analyses of sow longevity traits, age at first farrowing and first-litter characteristics. In Proceedings of the the 23rd Association for the Advancement of Animal Breeding and Genetics Conference, Armidale, NSW, Australia, 27 October–1 November 2019; Volume 23, pp. 167–170. [Google Scholar]
- Engblom, L.; Calderón Díaz, J.A.; Nikkilä, M.; Gray, K.; Harms, P.; Fix, J.; Tsuruta, S.; Mabry, J.; Stalder, K. Genetic analysis of sow longevity and sow lifetime reproductive traits using censored data. J. Anim. Breed. Genet. 2016, 133, 138–144. [Google Scholar] [CrossRef]
- Serenius, T.; Stalder, K.J.; Fernando, R.L. Genetic associations of sow longevity with age at first farrowing, number of piglets weaned, and wean to insemination interval in the Finnish Landrace swine population1. J. Anim. Sci. 2008, 86, 3324–3329. [Google Scholar] [CrossRef] [Green Version]
- Paixão, G.; Martins, Â.; Esteves, A.; Payan-Carreira, R.; Carolino, N. Genetic parameters for reproductive, longevity and lifetime production traits in Bísaro pigs. Livest. Sci. 2019, 225, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Cavalcante-Neto, A.; Lui, J.F.; Sarmento, J.L.R.; Ribeiro, M.N.; Fonseca, C.; Tonhati, H.; Monteiro, J.M.C.; Lara, M.A.C.; Queiroz, S.A. Genetic and environmental effects on age at first farrowing in sows in southeastern Brazil. Genet. Mol. Res. 2011, 10, 2860–2866. [Google Scholar] [CrossRef]
- Ogawa, S.; Konta, A.; Kimata, M.; Ishii, K.; Uemoto, Y.; Satoh, M. Estimation of genetic parameters for farrowing traits in purebred Landrace and Large White pigs. Anim. Sci. J. 2019, 90, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Uzzaman, M.R.; Park, J.E.; Lee, K.T.; Cho, E.S.; Choi, B.H.; Kim, T.H. A genome-wide association study of reproductive traits in a Yorkshire pig population. Livest. Sci. 2018, 209, 67–72. [Google Scholar] [CrossRef]
- Hanenberg, E.H.A.T.; Knol, E.F.; Merks, J.W.M. Estimates of genetic parameters for reproduction traits at different parities in Dutch Landrace pigs. Livest. Prod. Sci. 2001, 69, 179–186. [Google Scholar] [CrossRef]
- Lee, J.H.; Song, K.D.; Lee, H.K.; Cho, K.H.; Park, H.C.; Park, K. Do Genetic parameters of reproductive and meat quality traits in Korean berkshire pigs. Asian Australas. J. Anim. Sci. 2015, 28, 1388–1393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Tan, C.; Hu, X.; Wang, A.; Wu, Z. Genetic parameters for reproductive traits at different parities in Large White Pigs. J. Anim. Sci. 2018, 96, 1215–1220. [Google Scholar] [CrossRef]
- Roehe, R.; Kennedy, B.W. Estimation of genetic parameters for litter size in Canadian Yorkshire and Landrace swine with each parity of farrowing treated as a different trait. J. Anim. Sci. 1995, 73, 2959. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Yang, Y.; Liao, K.; Liu, B.; Chen, Y.; Shen, L.; Chen, L.; Jiang, A.; Liu, Y.; Li, Q.; et al. Genetic parameter estimation for reproductive traits in QingYu pigs and comparison of carcass and meat quality traits to Berkshire×QingYu crossbred pigs. Asian Australas. J. Anim. Sci. 2020, 33, 1224–1232. [Google Scholar] [CrossRef] [Green Version]
- Camargo, E.G.; Marques, D.B.D.; de Figueiredo, E.A.P.; E Silva, F.F.; Lopes, P.S. Genetic study of litter size and litter uniformity in Landrace pigs. Rev. Bras. Zootec. 2020, 49. [Google Scholar] [CrossRef]
- Canario, L.; Lundgren, H.; Haandlykken, M.; Rydhmer, L. Genetics of growth in piglets and the association with homogeneity of body weight within litters. J. Anim. Sci. 2010, 88, 1240–1247. [Google Scholar] [CrossRef]
- Wolf, J.; Žáková, E.; Groeneveld, E. Within-litter variation of birth weight in hyperprolific Czech Large White sows and its relation to litter size traits, stillborn piglets and losses until weaning. Livest. Sci. 2008, 115, 195–205. [Google Scholar] [CrossRef]
- Roehe, R. Genetic determination of individual birth weight and its association with sow productivity traits using Bayesian analyses. J. Anim. Sci. 1999, 77, 330. [Google Scholar] [CrossRef]
- Serenius, T.; Sevón-aimonen, M.-L.; Kause, A.; Mäntysaari, E.A.; Mäki-tanila, A. Selection potential of different prolificacy traits in the finnish landrace and large white populations. Acta Agric. Scand. Sect. A Anim. Sci. 2004, 54, 36–43. [Google Scholar] [CrossRef]
- KAIA (Korea Animal Improvement Association). 2020 Annual Report (In Korean: 2020년도 종축개량사업보고서); Korea Animal Improvement Association: Seoul, Korea, 2020; p. 156. [Google Scholar]



{kind=link}
{kind=link}
| Breed | Trait | N | Mean | SD | Minimum Value | Maximum Value | CV |
|---|---|---|---|---|---|---|---|
| Duroc | ADG | 115,501 | 666.11 | 60.42 | 470.00 | 906.80 | 9.07 |
| DAYS105 | 115,501 | 156.31 | 12.79 | 115.42 | 214.17 | 8.18 | |
| BFT | 115,501 | 12.55 | 2.37 | 1.83 | 24.10 | 18.89 | |
| Landrace | ADG | 116,870 | 643.07 | 56.82 | 468.00 | 908.20 | 8.84 |
| DAYS105 | 116,870 | 161.21 | 12.98 | 113.60 | 215.33 | 8.05 | |
| BFT | 116,870 | 12.68 | 2.50 | 1.88 | 24.18 | 19.75 | |
| Yorkshire | ADG | 368,021 | 641.37 | 57.35 | 468.00 | 908.20 | 8.94 |
| DAYS105 | 368,021 | 161.36 | 13.10 | 115.78 | 215.33 | 8.12 | |
| BFT | 368,021 | 13.27 | 2.53 | 1.83 | 24.20 | 19.07 |
| Breed | Trait | N | Mean | SD | Minimum Value | Maximum Value | CV |
|---|---|---|---|---|---|---|---|
| Duroc | AFF | 8906 | 370.86 | 23.66 | 324 | 438 | 6.38 |
| TNB | 8906 | 9.28 | 2.64 | 1 | 20 | 28.43 | |
| NBA | 8906 | 8.28 | 2.55 | 1 | 19 | 30.77 | |
| Landrace | AFF | 11,758 | 362.73 | 23.08 | 324 | 438 | 6.36 |
| TNB | 11,758 | 11.53 | 3.18 | 1 | 20 | 27.60 | |
| NBA | 11,757 | 10.63 | 2.98 | 0 | 20 | 28.02 | |
| Yorkshire | AFF | 40,171 | 368.82 | 22.95 | 324 | 438 | 6.22 |
| TNB | 40,171 | 12.07 | 3.40 | 1 | 20 | 28.13 | |
| NBA | 40,167 | 11.04 | 3.18 | 0 | 20 | 28.85 |
| Breed | Trait | Genetic Parameter Estimates | Genetic Variance | Residual Variance | CVg | ||
|---|---|---|---|---|---|---|---|
| ADG | DAYS105 | BFT | |||||
| Duroc | ADG | 0.36 ± 0.01 | −0.97 ± 0.00 | −0.01 ± 0.02 | 868.90 | 1578.00 | 0.04 |
| DAYS105 | −0.97 ± 0.00 | 0.44 ± 0.01 | −0.01 ± 0.02 | 41.80 | 53.54 | 0.04 | |
| BFT | −0.10 ± 0.00 | 0.07 ± 0.00 | 0.38 ± 0.01 | 1.44 | 2.31 | 0.10 | |
| Landrace | ADG | 0.36 ± 0.01 | −0.97 ± 0.00 | 0.00 ± 0.02 | 776.60 | 1386.00 | 0.04 |
| DAYS105 | −0.97 ± 0.00 | 0.42 ± 0.01 | −0.01 ± 0.02 | 38.78 | 54.55 | 0.04 | |
| BFT | −0.05 ± 0.00 | 0.04 ± 0.00 | 0.48 ± 0.01 | 2.39 | 2.59 | 0.12 | |
| Yorkshire | ADG | 0.34 ± 0.00 | −0.97 ± 0.00 | 0.04 ± 0.01 | 720.90 | 1382.00 | 0.04 |
| DAYS105 | −0.97 ± 0.00 | 0.41 ± 0.00 | −0.06 ± 0.01 | 38.30 | 54.81 | 0.04 | |
| BFT | −0.03 ± 0.00 | 0.01 ± 0.00 | 0.45 ± 0.00 | 2.26 | 2.74 | 0.11 | |
| Breed | Trait | Genetic Parameter Estimates | Genetic Variance | Residual Variance | CVg | ||
|---|---|---|---|---|---|---|---|
| AFF | TNB | NBA | |||||
| Duroc | AFF | 0.14 ± 0.02 | −0.02 ± 0.15 | −0.13 ± 0.15 | 39.96 | 248.00 | 0.02 |
| TNB | 0.02 ± 0.01 | 0.09 ± 0.02 | 0.90 ± 0.04 | 0.56 | 5.83 | 0.08 | |
| NBA | 0.00 ± 0.01 | 0.86 ± 0.00 | 0.09 ± 0.02 | 0.52 | 5.42 | 0.09 | |
| Landrace | AFF | 0.07 ± 0.02 | −0.21 ± 0.14 | −0.23 ± 0.14 | 21.08 | 261.40 | 0.01 |
| TNB | −0.01 ± 0.01 | 0.10 ± 0.02 | 0.94 ± 0.02 | 0.87 | 7.83 | 0.08 | |
| NBA | −0.03 ± 0.01 | 0.90 ± 0.00 | 0.10 ± 0.02 | 0.77 | 7.08 | 0.08 | |
| Yorkshire | AFF | 0.12 ± 0.01 | −0.01 ± 0.06 | −0.04 ± 0.06 | 37.88 | 277.00 | 0.02 |
| TNB | 0.00 ± 0.01 | 0.11 ± 0.01 | 0.96 ± 0.01 | 0.99 | 8.35 | 0.08 | |
| NBA | 0.00 ± 0.01 | 0.90 ± 0.00 | 0.09 ± 0.01 | 0.77 | 7.75 | 0.08 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alam, M.; Chang, H.-K.; Lee, S.-S.; Choi, T.-J. Genetic Analysis of Major Production and Reproduction Traits of Korean Duroc, Landrace and Yorkshire Pigs. Animals 2021, 11, 1321. https://doi.org/10.3390/ani11051321
Alam M, Chang H-K, Lee S-S, Choi T-J. Genetic Analysis of Major Production and Reproduction Traits of Korean Duroc, Landrace and Yorkshire Pigs. Animals. 2021; 11(5):1321. https://doi.org/10.3390/ani11051321
Chicago/Turabian StyleAlam, Mahboob, Hyuk-Kee Chang, Seung-Soo Lee, and Tae-Jeong Choi. 2021. "Genetic Analysis of Major Production and Reproduction Traits of Korean Duroc, Landrace and Yorkshire Pigs" Animals 11, no. 5: 1321. https://doi.org/10.3390/ani11051321
APA StyleAlam, M., Chang, H. -K., Lee, S. -S., & Choi, T. -J. (2021). Genetic Analysis of Major Production and Reproduction Traits of Korean Duroc, Landrace and Yorkshire Pigs. Animals, 11(5), 1321. https://doi.org/10.3390/ani11051321