
The Use of Molecular Profiling to Track Equine Reinfection Rates of Cyathostomin Species Following Anthelmintic Administration


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Fecal Sample Collection
2.3. Fecal Egg Count Reduction and Egg Reappearance Period Tests
2.4. DNA Extraction and Sequencing
2.5. Phylogenetic Analysis
2.6. Statistical Analysis and Species Frequency Reductions
3. Results
3.1. Sequencing Results
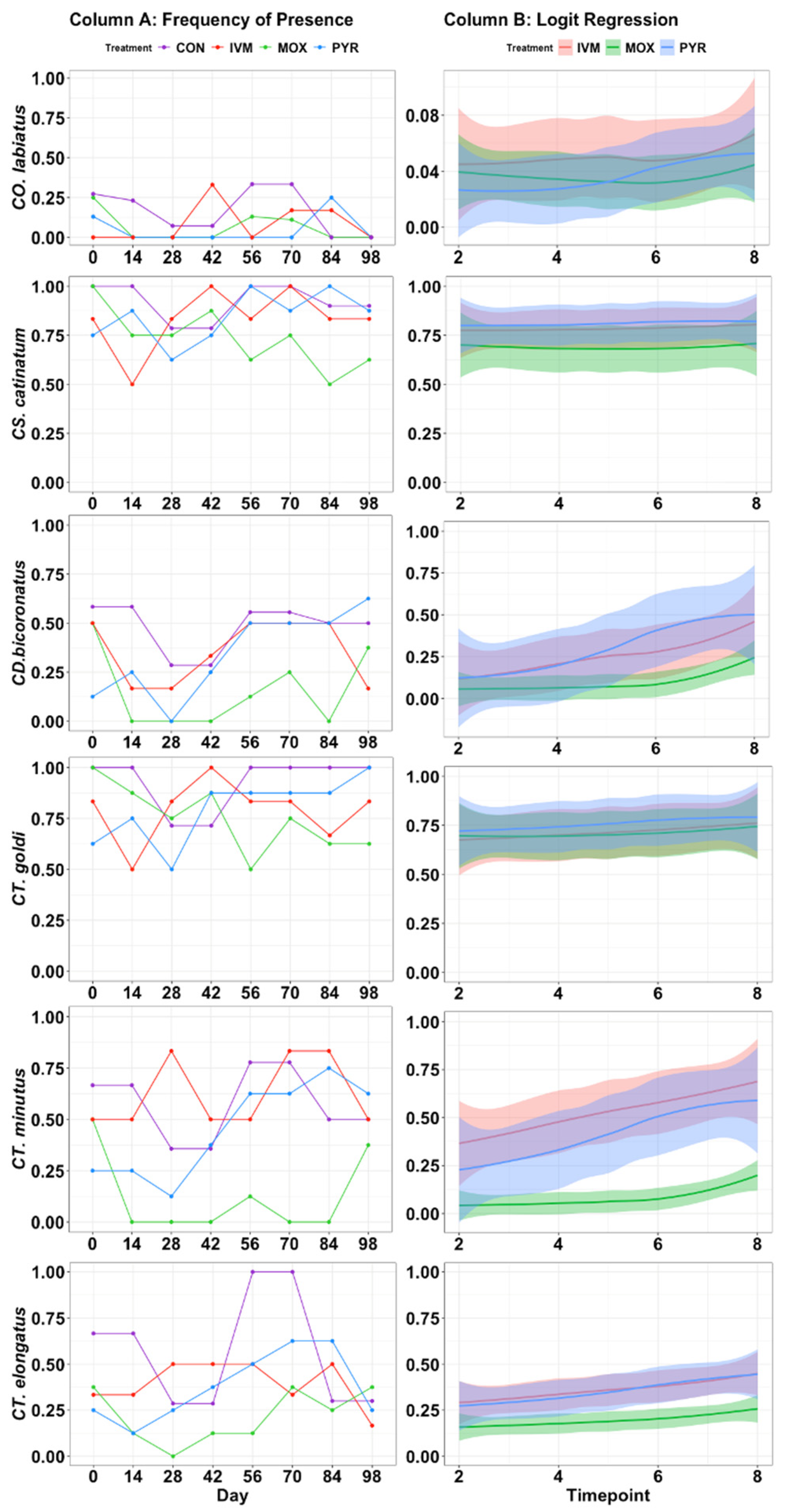
3.2. Fecal Egg and Species Count Reductions
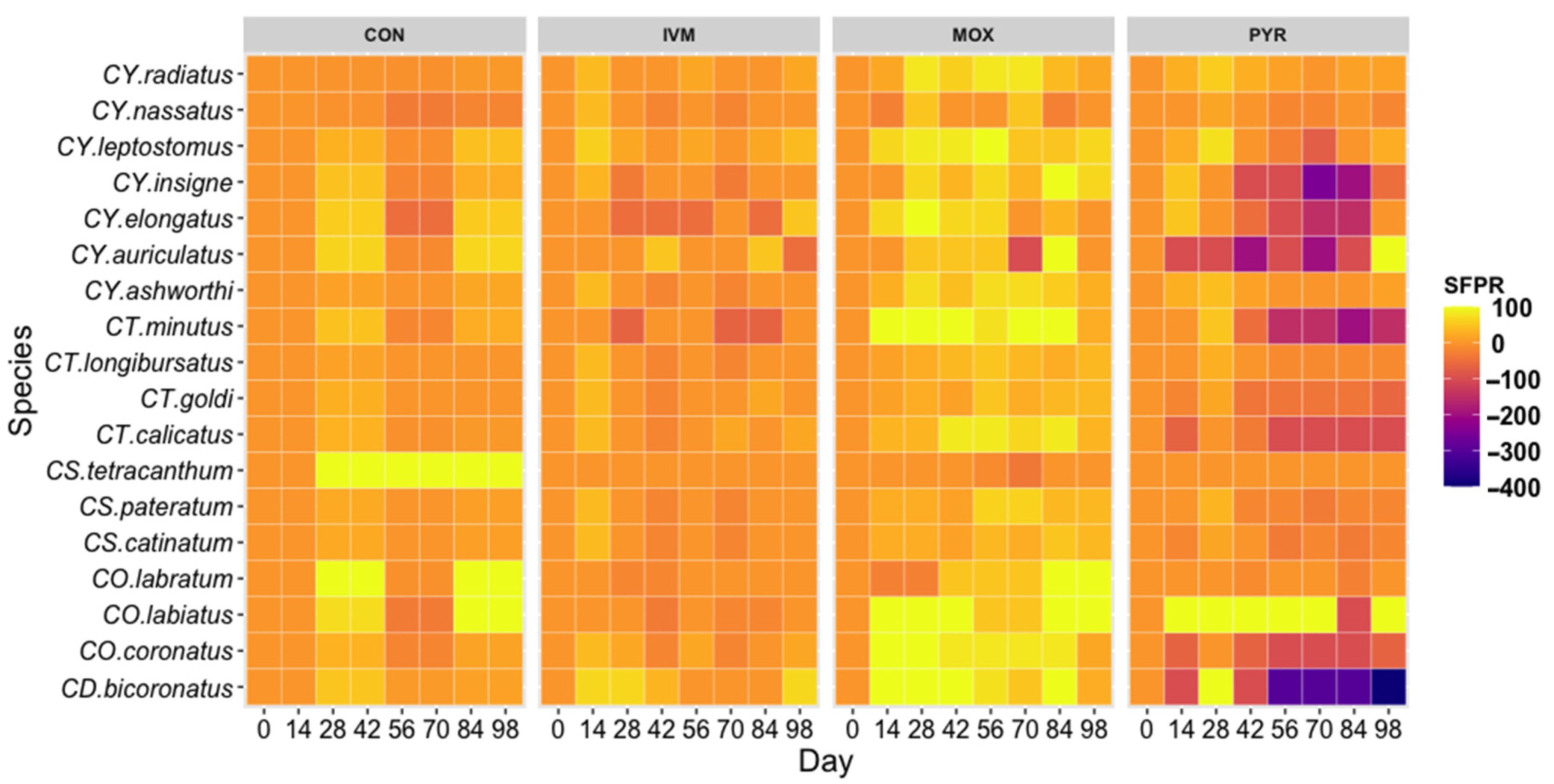
3.3. Anthelmintic Resistance
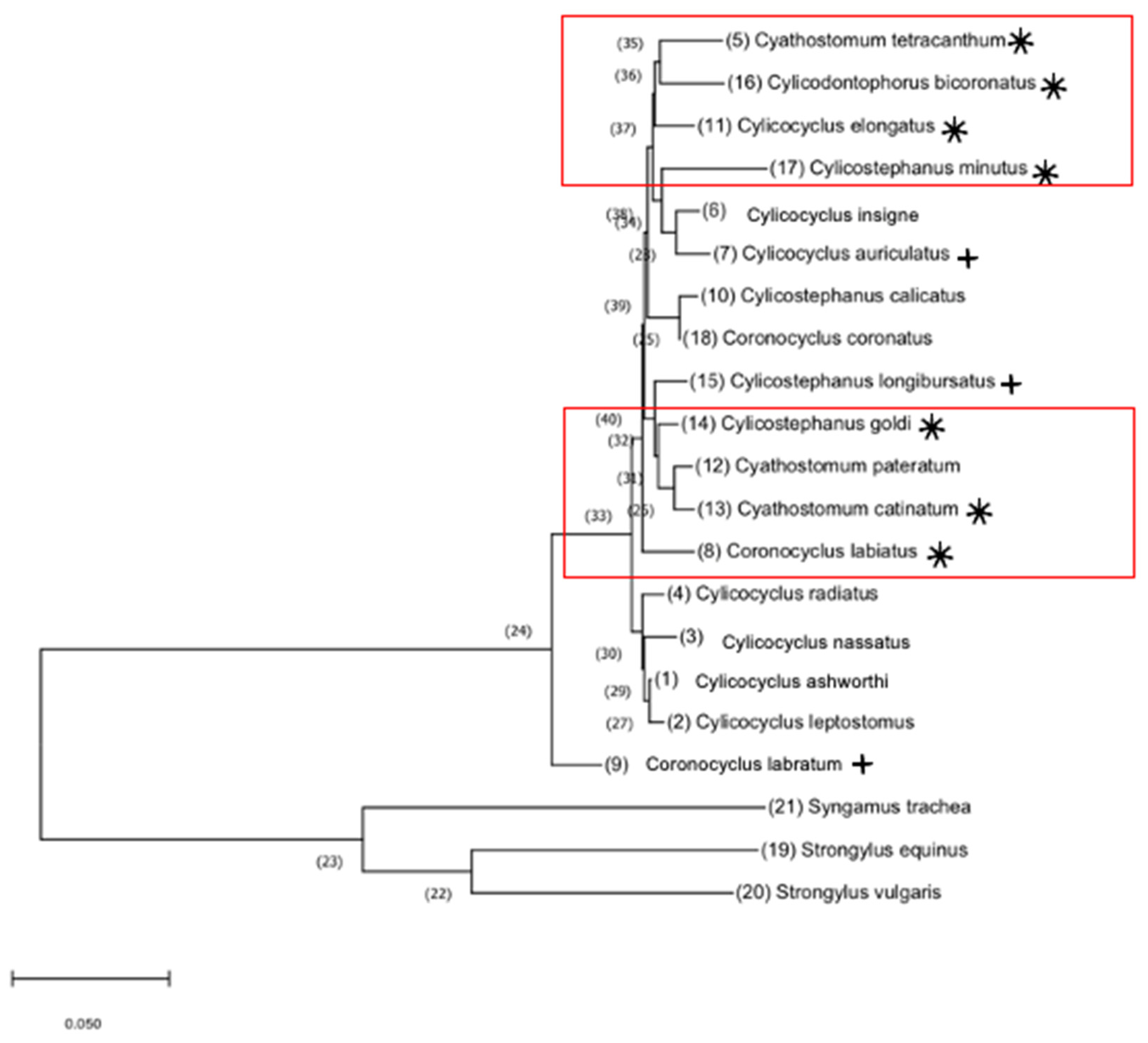
3.4. Phylogeny
4. Discussion
4.1. Fecal Egg Count Reductions and Species Frequency of Presence Reductions
4.2. Anthelmintic Resistance
4.3. Phylogeny
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nielsen, M.K.; Kaplan, R.M.; Thamsborg, S.M.; Monrad, J.; Olsen, S.N. Climatic Influences on Development and Survival of Free-Living Stages of Equine Strongyles: Implications for Worm Control Strategies and Managing Anthelmintic Resistance. Vet. J. 2007, 174, 23–32. [Google Scholar] [CrossRef]
- Morariu, S.; Mederle, N.; Badea, C.; Dărăbuş, G.; Ferrari, N.; Genchi, C. The Prevalence, Abundance and Distribution of Cyathostomins (Small Stongyles) in Horses from Western Romania. Vet. Parasitol. 2016, 223, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Mfitilodze, M.W.; Hutchinson, G.W. Prevalence and Abundance of Equine Strongyles (Nematoda: Strongyloidea) in Tropical Australia. J. Parasitol. 1990, 76, 487–494. [Google Scholar] [CrossRef]
- Bucknell, D.G.; Gasser, R.B.; Beveridge, I. The Prevalence and Epidemiology of Gastrointestinal Parasites of Horses in Victoria, Australia. Int. J. Parasitol. 1995, 25, 711–724. [Google Scholar] [CrossRef]
- Chapman, M.R.; French, D.D.; Klei, T.R. Gastrointestinal helminths of ponies in louisiana: A comparison of species currently prevalent with those present 20 years ago. J. Parasitol. 2002, 88, 1130–1134. [Google Scholar] [CrossRef]
- Love, S.; Murphy, D.; Mellor, D. Pathogenicity of Cyathostome Infection. Vet. Parasitol. 1999, 85, 113–122. [Google Scholar] [CrossRef]
- Garcia, A.; Brady, H.A.; Nichols, W.T.; Prien, S. Equine Cyathostomin Resistance to Fenbendazole in Texas Horse Facilities. J. Equine Vet. Sci. 2013, 33, 223–228. [Google Scholar] [CrossRef]
- Peregrine, A.S.; McEwen, B.; Bienzle, D.; Koch, T.G.; Weese, J.S. Larval Cyathostominosis in Horses in Ontario: An Emerging Disease? Can. Vet. J. 2006, 47, 80–82. [Google Scholar]
- Uhlinger, C. Equine Small Strongyles: Epidemiology, Pathology, and Control. Compend. Contin. Educ. Pract. Vet. 1991, 863. Available online: https://agris.fao.org/agris-search/search.do?recordID=US9143595. (accessed on 10 February 2021).
- Murphy, D.; Love, S. The Pathogenic Effects of Experimental Cyathostome Infections in Ponies. Vet. Parasitol. 1997, 70, 99–110. [Google Scholar] [CrossRef]
- Mair, T.S.; Cripps, P.J.; Ricketts, S.W. Diagnostic and Prognostic Value of Serum Protein Electrophoresis in Horses with Chronic Diarrhoea. Equine Vet. J. 1993, 25, 324–326. [Google Scholar] [CrossRef]
- Gokbulut, C.; McKellar, Q.A. Anthelmintic Drugs Used in Equine Species. Vet. Parasitol. 2018, 261, 27–52. [Google Scholar] [CrossRef]
- Kaplan, R.M. Anthelmintic Resistance in Nematodes of Horses. Vet. Res. 2002, 33, 491–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Samson-Himmelstjerna, G.; Fritzen, B.; Demeler, J.; Schürmann, S.; Rohn, K.; Schnieder, T.; Epe, C. Cases of Reduced Cyathostomin Egg-Reappearance Period and Failure of Parascaris Equorum Egg Count Reduction Following Ivermectin Treatment as Well as Survey on Pyrantel Efficacy on German Horse Farms. Vet. Parasitol. 2007, 144, 74–80. [Google Scholar] [CrossRef]
- Kaplan, R.M.; Klei, T.R.; Lyons, E.T.; Lester, G.; Courtney, C.H.; French, D.D.; Tolliver, S.C.; Vidyashankar, A.N.; Zhao, Y. Prevalence of Anthelmintic Resistant Cyathostomes on Horse Farms. J. Am. Vet. Med. Assoc. 2004, 225, 903–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, M.K.; Banahan, M.; Kaplan, R.M. Importation of Macrocyclic Lactone Resistant Cyathostomins on a US Thoroughbred Farm. Int. J. Parasitol. Drugs Drug Resist. 2020, 14, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Traversa, D.; von Samson-Himmelstjerna, G.; Demeler, J.; Milillo, P.; Schürmann, S.; Barnes, H.; Otranto, D.; Perrucci, S.; di Regalbono, A.F.; Beraldo, P.; et al. Anthelmintic Resistance in Cyathostomin Populations from Horse Yards in Italy, United Kingdom and Germany. Parasit. Vectors 2009, 2, S2. [Google Scholar] [CrossRef] [Green Version]
- Peregrine, A.S.; Molento, M.B.; Kaplan, R.M.; Nielsen, M.K. Anthelmintic Resistance in Important Parasites of Horses: Does It Really Matter? Vet. Parasitol. 2014, 201, 1–8. [Google Scholar] [CrossRef]
- Canever, R.J.; Braga, P.R.C.; Boeckh, A.; Grycajuck, M.; Bier, D.; Molento, M.B. Lack of Cyathostomin Sp. Reduction after Anthelmintic Treatment in Horses in Brazil. Vet. Parasitol. 2013, 194, 35–39. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, D.E.; Hutchinson, M.J.; Parker, L.; Gibbons, L.M. Equine Cyathostome Infection: Suppression of Faecal Egg Output with Moxidectin. Vet. Rec. 1995, 137, 545. [Google Scholar] [CrossRef]
- DiPietro, J.A.; Hutchens, D.E.; Lock, T.F.; Walker, K.; Paul, A.J.; Shipley, C.; Rulli, D. Clinical Trial of Moxidectin Oral Gel in Horses. Vet. Parasitol. 1997, 72, 167–177. [Google Scholar] [CrossRef]
- Demeulenaere, D.; Vercruysse, J.; Dorny, P.; Claerebout, E. Comparative Studies of Ivermectin and Moxidectin in the Control of Naturally Acquired Cyathostome Infections in Horses. Vet. Rec. 1997, 141, 383–386. [Google Scholar] [CrossRef]
- Borgsteede, F.H.M.; Boersma, J.H.; Gaasenbeek, C.P.H.; van der Burg, W.P.J. The Reappearance of Eggs in Faeces of Horses after Treatment with Ivermectin. Vet. Q. 1993, 15, 24–26. [Google Scholar] [CrossRef]
- Boersema, J.H.; Eysker, M.; Maas, J.; van der Aar, W.M. Comparison of the Reappearance of Strongyle Eggs in Foals, Yearlings, and Adult Horses after Treatment with Ivermectin or Pyrantel. Vet. Q. 1996, 18, 7–9. [Google Scholar] [CrossRef] [Green Version]
- Boersema, J.H.; Borgsteede, F.H.M.; Eysker, M.; Saedt, I. The Reappearance of Strongyle Eggs in Faeces of Horses Treated with Pyrantel Embonate. Vet. Q. 1995, 17, 18–20. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.K.; Mittel, L.; Grice, A.; Erksine, M.; Graves, E.; Vaala, W.; Tully, R.C.; French, D.D.; Bowman, R.; Kaplan, R.M. AAEP Parasite Control Guidelines 2019. Available online: http://www.aaep.org/custdocs/ParasiteControlGuidelinesFinal.pdf (accessed on 10 February 2021).
- Scare, J.A.; Lyons, E.T.; Wielgus, K.M.; Nielsen, M.K. Combination Deworming for the Control of Double-Resistant Cyathostomin Parasites–Short and Long Term Consequences. Vet. Parasitol. 2018, 251, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Gilleard, J.S.; Beech, R.N. Population Genetics of Anthelmintic Resistance in Parasitic Nematodes. Parasitology 2007, 134.8, 1133. [Google Scholar] [CrossRef] [PubMed]
- Redman, E.; Whitelaw, F.; Tait, A.; Burgess, C.; Bartley, Y.; Skuce, P.J.; Jackson, F.; Gilleard, J.S. The Emergence of Resistance to the Benzimidazole Anthlemintics in Parasitic Nematodes of Livestock Is Characterised by Multiple Independent Hard and Soft Selective Sweeps. PLoS Negl. Trop. Dis. 2015, 9, e0003494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leathwick, D.M.; Sauermann, C.W.; Nielsen, M.K. Managing Anthelmintic Resistance in Cyathostomin Parasites: Investigating the Benefits of Refugia-Based Strategies. Int. J. Parasitol. Drugs Drug Resist. 2019, 10, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Corning, S. Equine Cyathostomins: A Review of Biology, Clinical Significance and Therapy. Parasit. Vectors 2009, 2, S1. [Google Scholar] [CrossRef] [Green Version]
- Bellaw, J.L.; Krebs, K.; Reinemeyer, C.R.; Norris, J.K.; Scare, J.A.; Pagano, S.; Nielsen, M.K. Anthelmintic Therapy of Equine Cyathostomin Nematodes–Larvicidal Efficacy, Egg Reappearance Period, and Drug Resistance. Int. J. Parasitol. 2018, 48, 97–105. [Google Scholar] [CrossRef]
- Hodgkinson, J.E.; Freeman, K.L.; Lichtenfels, J.R.; Palfreman, S.; Love, S.; Matthews, J.B. Identification of Strongyle Eggs from Anthelmintic-Treated Horses Using a PCR-ELISA Based on Intergenic DNA Sequences. Parasitol. Res. 2005, 95, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Traversa, D.; Iorio, R.; Otranto, D.; Giangaspero, A.; Milillo, P.; Klei, T.R. Species-Specific Identification of Equine Cyathostomes Resistant to Fenbendazole and Susceptible to Oxibendazole and Moxidectin by Macroarray Probing. Exp. Parasitol. 2009, 121, 92–95. [Google Scholar] [CrossRef]
- Ionita, M.; Howe, D.K.; Lyons, E.T.; Tolliver, S.C.; Kaplan, R.M.; Mitrea, I.L.; Yeargan, M. Use of a Reverse Line Blot Assay to Survey Small Strongyle (Strongylida: Cyathostominae) Populations in Horses before and after Treatment with Ivermectin. Vet. Parasitol. 2010, 168, 332–337. [Google Scholar] [CrossRef]
- Čerňanská, D.; Paoletti, B.; Kráľová-Hromadová, I.; Iorio, R.; Čudeková, P.; Milillo, P.; Traversa, D. Application of a Reverse Line Blot Hybridisation Assay for the Species-Specific Identification of Cyathostomins (Nematoda, Strongylida) from Benzimidazole-Treated Horses in the Slovak Republic. Vet. Parasitol. 2009, 160, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Kooyman, F.N.J.; van Doorn, D.C.K.; Geurden, T.; Wagenaar, J.A. Semi-Quantitative Differentiation of Cyathostomin Larval Cultures by Reverse Line Blot. Vet. Parasitol. 2016, 216, 59–65. [Google Scholar] [CrossRef]
- Bredtmann, C.M.; Krücken, J.; Murugaiyan, J.; Kuzmina, T.; von Samson-Himmelstjerna, G. Nematode Species Identification—Current Status, Challenges and Future Perspectives for Cyathostomins. Front. Cell. Infect. Microbiol. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Van Doorn, D.C.K.; Ploeger, H.W.; Eysker, M.; Geurden, T.; Wagenaar, J.A.; Kooyman, F.N.J. Cylicocyclus Species Predominate during Shortened Egg Reappearance Period in Horses after Treatment with Ivermectin and Moxidectin. Vet. Parasitol. 2014, 206, 246–252. [Google Scholar] [CrossRef]
- Hodgkinson, J.E.; Lichtenfels, J.R.; Mair, T.S.; Cripps, P.; Freeman, K.L.; Ramsey, Y.H.; Love, S.; Matthews, J.B. A PCR–ELISA for the Identification of Cyathostomin Fourth-Stage Larvae from Clinical Cases of Larval Cyathostominosis. Int. J. Parasitol. 2003, 33, 1427–1435. [Google Scholar] [CrossRef] [Green Version]
- Hodgkinson, J.E.; Love, S.; Lichtenfels, J.R.; Palfreman, S.; Ramsey, Y.H.; Matthews, J.B. Evaluation of the Specificity of Five Oligoprobes for Identification of Cyathostomin Species from Horses. Int. J. Parasitol. 2001, 31, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Chilton, N.B.; Hoste, H.; Hung, G.-C.; Beveridge, I.; Gasser, R.B. The 5.8S RDNA Sequences of 18 Species of Bursate Nematodes (Order Strongylida): Comparison with Rhabditid and Tylenchid Nematodes. Int. J. Parasitol. 1997, 27, 119–124. [Google Scholar] [CrossRef]
- Leathers, D.; Office of The Delaware State Climatologist. Off. Del. State Climatol. Available online: http://climate.udel.edu/search-results/ (accessed on 10 February 2021).
- Larsen, M.L.; Ritz, C.; Petersen, S.L.; Nielsen, M.K. Determination of Ivermectin Efficacy against Cyathostomins and Parascaris Equorum on Horse Farms Using Selective Therapy. Vet. J. 2011, 188, 44–47. [Google Scholar] [CrossRef]
- Collins, A.; Ke, X. Primer1: Primer Design Web Service for Tetra-Primer ARMS-PCR. Open Bioinforma. J. 2012, 6, 55–58. [Google Scholar] [CrossRef]
- Elela, S.A.; Nazar, R.N. Role of the 5.8S RRNA in Ribosome Translocation. Nucleic Acids Res. 1997, 25, 1788–1794. [Google Scholar] [CrossRef] [Green Version]
- Dimitrova, A.D.; Gecheff, K.I.; Ananiev, E.D. Methylation pattern of ribosomal rna genes in nor-deleted and nor- reconstructed barley lines (hordeum vulgare L.). organization of igs in rdna repeat unit. Genet. Plant. Phys. 2012, 2, 3–14. [Google Scholar]
- Magoc, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenfels, J.R.; Kharchenko, V.A.; Dvojnos, G.M. Illustrated Identification Keys to Strongylid Parasites (Strongylidae: Nematoda) of Horses, Zebras and Asses (Equidae). Vet. Parasitol. 2008, 156, 4–161. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecson, B.M.; Barrios, J.A.; Johnson, D.R.; Nelson, K.L. A Real-Time PCR Method for Quantifying Viable Ascaris Eggs Using the First Internally Transcribed Spacer Region of Ribosomal DNA. APPL Env. Microbiol 2006, 72, 9. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundatioon for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Lyons, E.T.; Tolliver, S.C.; Drudge, J.H. Historical Perspective of Cyathostomes: Prevalence, Treatment and Control Programs. Vet. Parasitol. 1999, 85, 97–112. [Google Scholar] [CrossRef]
- Ogbourne, C.P. Observations on the Free-Living Stages of Strongylid Nematodes of the Horse. Parasitology 1972, 64, 461–477. [Google Scholar] [CrossRef]
- Silva, A.V.M.; Costa, H.M.A.; Santos, H.A.; Carvalho, R.O. Cyathostominae (Nematoda) Parasites of Equus Caballus in Some Brazilian States. Vet. Parasitol. 1999, 86, 15–21. [Google Scholar] [CrossRef]
- Kuzmina, T.A.; Kharchenko, V.A.; Starovir, A.I.; Dvojnos, G.M. Analysis of the Strongylid Nematodes (Nematoda: Strongylidae) Community after Deworming of Brood Horses in Ukraine. Vet. Parasitol. 2005, 131, 283–290. [Google Scholar] [CrossRef]
- Reinemeyer, C.R.; Smith, S.A.; Gabel, A.A.; Herd, R.P. The Prevalence and Intensity of Internal Parasites of Horses in the U.S.A. Vet. Parasitol. 1984, 15, 75–83. [Google Scholar] [CrossRef]
- Zanet, S.; Battisti, E.; Labate, F.; Oberto, F.; Ferroglio, E. Reduced Efficacy of Fenbendazole and Pyrantel Pamoate Treatments against Intestinal Nematodes of Stud and Performance Horses. Vet. Sci. 2021, 8, 42. [Google Scholar] [CrossRef]
- Sauermann, C.W.; Leathwick, D.M.; Lieffering, M.; Nielsen, M.K. Climate Change Is Likely to Increase the Development Rate of Anthelmintic Resistance in Equine Cyathostomins in New Zealand. Int. J. Parasitol. Drugs Drug Resist. 2020, 14, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Rupasinghe, D.; Ogbourne, C.P. Laboratory Studies on the Effect of Temperature on the Development of the Free-Living Stages of Some Strongylid Nematodes of the Horse. Z. Für Parasitenkd. 1978, 55, 249–253. [Google Scholar] [CrossRef]
- Mfitilodze, M.W.; Hutchinson, G.W. Development and Survival of Free-Living Stages of Equine Strongyles under Laboratory Conditions. Vet. Parasitol. 1987, 23, 121–133. [Google Scholar] [CrossRef]
- Rossanigo, C.E.; Gruner, L. Moisture and Temperature Requirements in Faeces for the Development of Free-Living Stages of Gastrointestinal Nematodes of Sheep, Cattle and Deer. J. Helminthol. 1995, 69, 357–362. [Google Scholar] [CrossRef]
- Von Ober-Blöbaum, W. Untersuchungen Über Die Einwirkungen Physikalischer Einflü Sse Auf Die Larven von Pferdestrongyliden. Tierartzl Rundsch. 1932, 47, 812–815. [Google Scholar]
- Lucker, J.T. Survival and development at low tempera tures of eggs and preinfective larvae horse strongyles. J. Agric. Res. 1941, 63, 193. [Google Scholar]
- Kaplan, R.M.; West, E.M.; Norat-Collazo, L.M.; Vargas, J. A Combination Treatment Strategy Using Pyrantel Pamoate and Oxibendazole Demonstrates Additive Effects for Controlling Equine Cyathostomins. Equine Vet. Educ. 2014, 26, 485–491. [Google Scholar] [CrossRef]
- Lyons, E.T.; Kuzmina, T.A.; Tolliver, S.C.; Collins, S.S. Observations on Development of Natural Infection and Species Composition of Small Strongyles in Young Equids in Kentucky. Parasitol. Res. 2011, 109, 1529–1535. [Google Scholar] [CrossRef]
- Barnes, E.H.; Dobson, R.J. Population Dynamics of Trichostrongylus Colubriformis in Sheep: Computer Model to Simulate Grazing Systems and the Evolution of Anthelmintie ResistancE. Int. J. Parasitol. 1990, 20, 823–831. [Google Scholar] [CrossRef]
- Dargatz, D.A.; Traub-Dargatz, J.L.; Sangster, N.C. Antimicrobic and Anthelmintic Resistance. Vet. Clin. North. Am. Equine Pract. 2000, 16, 515–536. [Google Scholar] [CrossRef]
- Sangster, N.C. Pharmacology of Anthelmintic Resistance in Cyathostomes: Will It Occur with the Avermectin/Milbemycins? Vet. Parasitol. 1999, 85, 189–204. [Google Scholar] [CrossRef]
- Dowdall, S.M.J.; Matthews, J.B.; Mair, T.; Murphy, D.; Love, S.; Proudman, C.J. Antigen-Specific IgG(T) Responses in Natural and Experimental Cyathostominae Infection in Horses. Vet. Parasitol. 2002, 106, 225–242. [Google Scholar] [CrossRef]
- Stratford, C.H.; McGorum, B.C.; Pickles, K.J.; Matthews, J.B. An Update on Cyathostomins: Anthelmintic Resistance and Diagnostic Tools. Equine Vet. J. 2011, 43, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Lyons, E.T.; Tolliver, S.C.; Drudge, J.H.; Collins, S.S.; Swerczek, T.W. Continuance of Studies on Population S Benzimidazole-Resistant Small Strongyles in a Shetland Pony Herd in Kentucky: Effect of Pyrantel Pamoate (1992–1999). Vet. Parasitol. 2001, 94, 247–256. [Google Scholar] [CrossRef]
- Gokbulut, C.; Nolan, A.M.; Mckellar, Q.A. Plasma Pharmacokinetics and Faecal Excretion of Ivermectin, Doramectin and Moxidectin Following Oral Administration in Horses. Equine Vet. J. 2001, 33, 494–498. [Google Scholar] [CrossRef]
- Dobson, R.J.; Lejambre, L.; Gill, J.H. Management of Anthelmintic Resistance: Inheritance of Resistance and Selection with Persistent Drugs. Int. J. Parasitol. 1996, 26, 993–1000. [Google Scholar] [CrossRef]
- Terrill, T.H.; Kaplan, R.M.; Larsen, M.; Samples, O.M.; Miller, J.E.; Gelaye, S. Anthelmintic Resistance on Goat Farms in Georgia: Efficacy of Anthelmintics against Gastrointestinal Nematodes in Two Selected Goat Herds. Vet. Parasitol. 2001, 97, 261–268. [Google Scholar] [CrossRef]
- Durette-Desset, M.-C.; Beveridge, I.; Spratt, D.M. The Origins and Evolutionary Expansion of the Strongylida (Nematoda). Int. J. Parasitol. 1994, 24, 1139–1165. [Google Scholar] [CrossRef]
- Cwiklinski, K.; Merga, J.Y.; Lake, S.L.; Hartley, C.; Matthews, J.B.; Paterson, S.; Hodgkinson, J.E. Transcriptome Analysis of a Parasitic Clade V Nematode: Comparative Analysis of Potential Molecular Anthelmintic Targets in Cylicostephanus Goldi. Int. J. Parasitol. 2013, 43, 917–927. [Google Scholar] [CrossRef] [Green Version]
- Tandon, R.; LePage, K.T.; Kaplan, R.M. Cloning and Characterization of Genes Encoding α and β Subunits of Glutamate-Gated Chloride Channel Protein in Cylicocyclus Nassatus. Mol. Biochem. Parasitol. 2006, 150, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Dorris, M.; De Ley, P.; Blaxter, M.L. Molecular Analysis of Nematode Diversity and the Evolution of Parasitism. Parasitol. Today 1999, 15, 188–193. [Google Scholar] [CrossRef]
- Blouin, M.S.; Yowell, C.A.; Courtney, C.H.; Dame, J.B. Host Movement and the Genetic Structure of Populations of Parasitic Nematodes. Genetics 1995, 141, 1007–1014. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Horse ID | Sex 1 | Age (Years) | Breed | Weight (kg) | Farm ID 2 |
|---|---|---|---|---|---|
| 1 | G | 10 | Arabian | 456 | 1 |
| 2 | M | 10 | Arabian | 449 | 1 |
| 3 | G | 8 | Quarter Horse | 600 | 1 |
| 4 | M | 10 | Quarter Horse | 534 | 1 |
| 5 | G | 7 | Thoroughbred | 490 | 1 |
| 6 | G | 16 | Standardbred | 470 | 1 |
| 7 * | G | 19 | Saddlebred | 493 | 2 † |
| 8 | M | 18 | Standardbred | 498 | 2 |
| 9 * | G | 33 | Morgan | 392 | 2 |
| Taxa | Accession Number |
|---|---|
| Cylicocyclus (CY) ashworthi | Y08586 |
| Cylicocyclus (CY) leptostomus | KP693432 |
| Cylicocyclus (CY) nassatus | Y08585 |
| Cylicocyclus (CY) radiatus | JQ906423 |
| Cyathostomum (CS) tetracanthum | KF850629 |
| Cylicocyclus (CY) insigne | Y08588 |
| Cylicocyclus (CY) auriculatus | JQ906414 |
| Coronocyclus (CO) labiatus | JN786947 |
| Coronocyclus (CO) labratum | AJ004838 |
| Cylicostephanus (CT) calicatus | KM085356 |
| Cylicocylus (CY) elongatus | JQ906417 |
| Cyathostomum (CS) pateratum | KF850627 |
| Cyathostomum (CS) catinatum | KF850626 |
| Cylicostephanus (CT) goldi | KM085357 |
| Cylicostephanus (CT) longibursatus | KM085358 |
| Cylicodontophorus (CD) bicoronatus | KP693441 |
| Cylicostephanus (CT) minutus | KM085361 |
| Coronocyclus (CO) coronatus | JN786951 |
| Poteriostomum (POT) imparidentatum | KP693433 |
| FECR 1/ERP 2 | Total Number of Species 3 | ||||||
|---|---|---|---|---|---|---|---|
| Day | IVM (%) | MOX (%) | PYR (%) | CON | IVM | MOX | PYR |
| 0 | -- | -- | -- | 13.16 ± 3.7 | 10.5 ± 5.4 | 10.9 ± 4.6 | 7.88 ± 4.6 |
| 14 | 100.00 | 100.00 | 98.37 | 13.16 ± 3.7 | 6.50 ± 7.2 | 7.13 ± 3.9 | 7.88 ± 4.1 |
| 28 | 96.15 | 100.00 | 72.29 * | 9.00 ± 5.0 | 10.7 ± 5.4 | 5.13 ± 3.3 | 5.50 ± 4.8 |
| 42 | 81.73 * | 98.58 | 49.46 | 9.00 ± 5.0 | 12.2 ± 2.6 | 5.63 ± 3.4 | 9.13 ± 4.9 |
| 56 | 53.85 | 96.45 | 7.63 | 14.33 ± 2.3 | 10.2 ± 5.7 | 4.25 ± 5.2 | 10.8 ± 4.1 |
| 70 | 67.31 | 93.85 | −91.29 | 14.33 ± 2.3 | 12.0 ± 3.3 | 6.00 ± 3.7 | 11.8 ± 3.3 |
| 84 | 34.62 | 83.91 * | −152.69 | 10.3 ± 3.7 | 10.7 ± 5.1 | 4.75 ± 3.1 | 11.5 ± 4.4 |
| 98 | 0.00 | 60.25 | −167.36 | 10.3 ± 5.7 | 9.33 ± 4.6 | 6.88 ± 6.2 | 9.75 ± 3.9 |
| p-Value | Spearman Correlation (r) | |||||
|---|---|---|---|---|---|---|
| Worm Species | CON | IVM | MOX | PYR | Treatment | MOX |
| CO. coronatus | 1.00 a | 0.77 ac | 0.20 b | 0.64 c | 3.36 × 10−6 *** | −0.44 |
| CO. labiatus | 0.17 a | 0.08 a | 0.06 a | 0.04 a | 0.403 | |
| CO. labratum | 0.19 a | 0.04 a | 0.24 a | 2.7 × 10−17 a | 0.038 * | 0.36 |
| CS. catinatum | 1.00 a | 0.83 a | 0.56 a | 0.83 a | 0.066 | |
| CS. tetracanthum | 0.02 a | 0.0 a | 0.06 a | 0.00 a | 0.281 | |
| CY. ashworthi | 1.00 a | 0.83 a | 0.47 b | 0.75 a | 0.0004 *** | −0.38 |
| CY. auriculatus | 0.42 a | 0.31 a | 0.21 a,b | 0.24 a | 0.045 * | |
| CY. insigne | 0.57 a | 0.52 a,c | 0.21 b | 0.46 b,c | 9.2 × 10−5 *** | −0.31 |
| CY. leptostomus | 0.76 a | 0.67 a | 0.27 b | 0.44 b | 1.13 × 10−5 *** | −0.32 |
| CY. nassatus | 0.85 a | 0.83 a | 0.47 b | 0.79 a | 3.04 × 10−5 *** | −0.34 |
| CY. radiatus | 0.97 a | 0.71 a | 0.33 b | 0.69 a | 0.0004 *** | −0.41 |
| CS. pateratum | 0.91 a | 0.83 a | 0.67 a | 0.81 a | 0.00159 *** | |
| CY. elongatus | 0.56 a | 0.31 a | 0.22 a | 0.38 a | 0.106 | |
| CD. bicoronatus | 0.48 a | 0.42 a | 0.16 b | 0.36 a | 0.108 | |
| CT. calicatus | 0.97 a | 0.77 a,c | 0.36 b | 0.61 b,c | 0.0005 *** | −0.31 |
| CT. goldi | 0.92 a | 0.79 a | 0.74 a | 0.78 a | 0.189 | |
| CT. longibursatus | 1.00 a | 0.83 a,b | 0.71 b | 0.89 a,b | 0.025 * | |
| CT. minutus | 0.57 a | 0.54 a | 0.13 b | 0.46 a | 0.074 | −0.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, A.C.B.; Biddle, A.S. The Use of Molecular Profiling to Track Equine Reinfection Rates of Cyathostomin Species Following Anthelmintic Administration. Animals 2021, 11, 1345. https://doi.org/10.3390/ani11051345
Johnson ACB, Biddle AS. The Use of Molecular Profiling to Track Equine Reinfection Rates of Cyathostomin Species Following Anthelmintic Administration. Animals. 2021; 11(5):1345. https://doi.org/10.3390/ani11051345
Chicago/Turabian StyleJohnson, Alexa C. B., and Amy S. Biddle. 2021. "The Use of Molecular Profiling to Track Equine Reinfection Rates of Cyathostomin Species Following Anthelmintic Administration" Animals 11, no. 5: 1345. https://doi.org/10.3390/ani11051345
APA StyleJohnson, A. C. B., & Biddle, A. S. (2021). The Use of Molecular Profiling to Track Equine Reinfection Rates of Cyathostomin Species Following Anthelmintic Administration. Animals, 11(5), 1345. https://doi.org/10.3390/ani11051345