
Hepatitis E Virus RNA Presence in Wild Boar Carcasses at Slaughterhouses in Italy


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
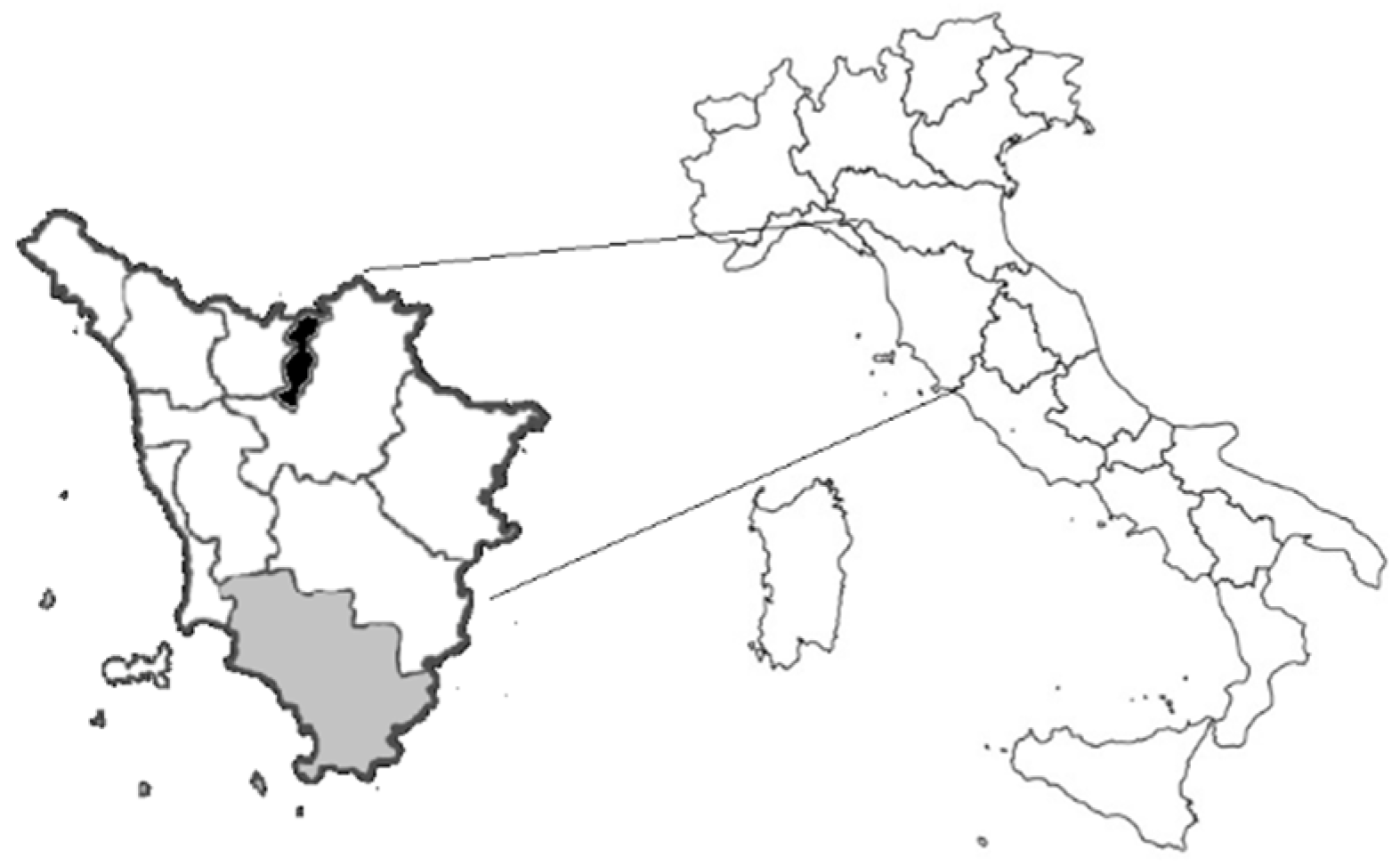
2.1. Samples Collection
2.2. RNA Extraction
2.3. Generation of HEV Standard Curve
2.4. RT-qPCR
2.5. PCR
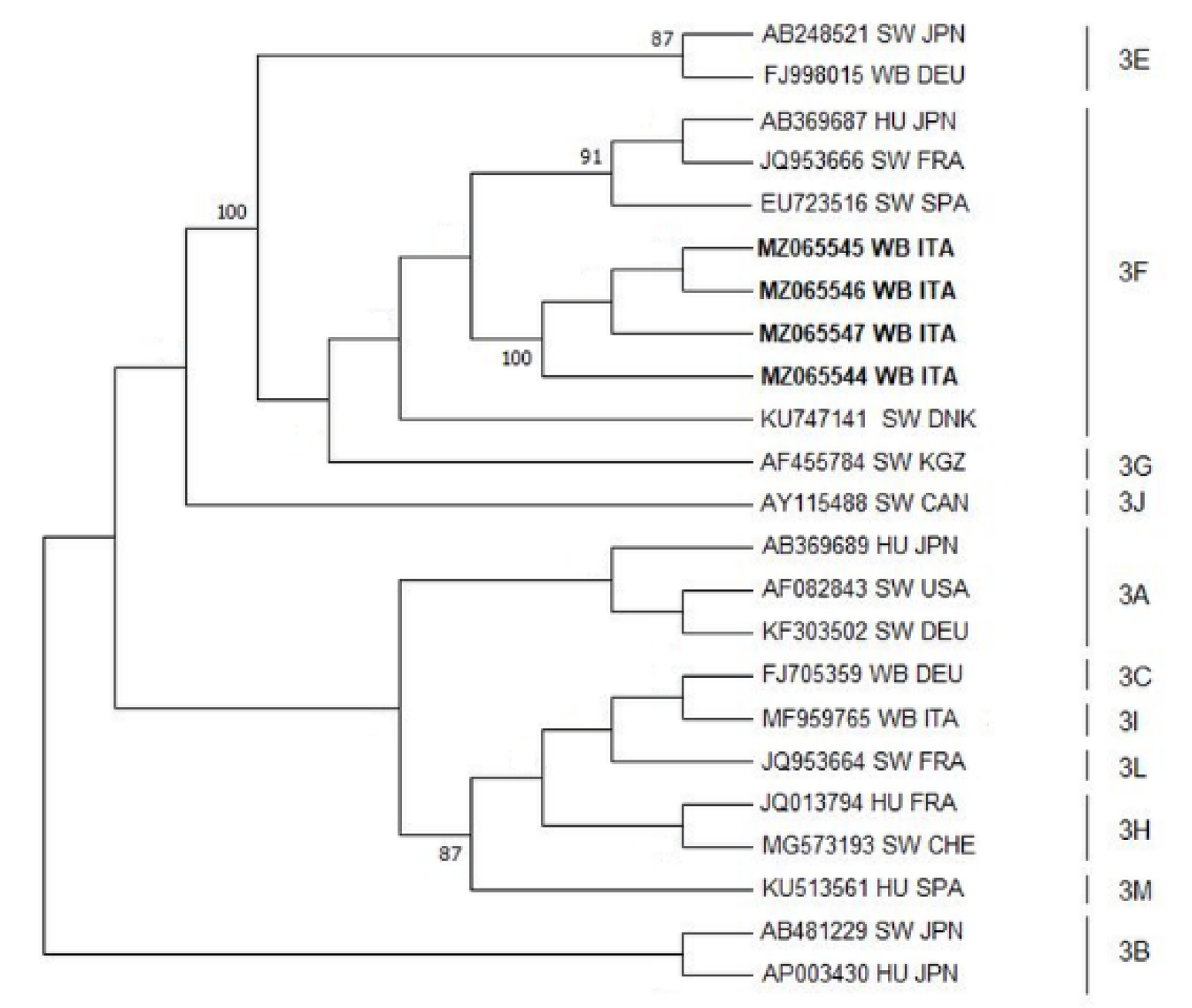
2.6. Sequence and Phylogenetic Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Rein, D.B.; Stevens, G.A.; Theaker, J.; Wittenborn, J.S.; Wiersma, S.T. The global burden of hepatitis E virus genotypes 1 and 2 in 2005. Hepatology 2012, 55, 988–997. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.J.; Norder, H.; Okamoto, H.; van der Poel, W.H.M.; Reuter, G.; et al. Update: Proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692–698. [Google Scholar] [CrossRef]
- Yin, X.; Li, X.; Feng, Z. Role of envelopment in the HEV life cycle. Viruses 2016, 8, 229. [Google Scholar] [CrossRef] [Green Version]
- Donnelly, M.C.; Scobie, L.; Crossan, C.L.; Dalton, H.; Hayes, P.C.; Simpson, K.J. Review article: Hepatitis E-a concise review of virology, epidemiology, clinical presentation and therapy. Aliment. Pharmacol. Ther. 2017, 46, 126–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aggarwal, R. The Global Prevalence of Hepatitis E Virus Infection and Susceptibility: A Systematic Review; WHO Document Production Services: Geneva, Switzerland, 2010. [Google Scholar]
- Doceul, V.; Bagdassarian, E.; Demange, A.; Pavio, N. Zoonotic hepatitis E virus: Classification, animal reservoirs and transmission routes. Viruses 2016, 8, 270. [Google Scholar] [CrossRef]
- Hartl, J.; Otto, B.; Madden, R.G.; Webb, G.; Woolson, K.L.; Kriston, L.; Vettorazzi, E.; Lohse, A.W.; Dalton, H.R.; Pischke, S. Hepatitis E seroprevalence in Europe: A meta-analysis. Viruses 2016, 8, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernandez Escamez, P.S.; Herman, L.; Koutsoumanis, K.; Lindqvist, R.; Nørrung, B.; et al. Public health risks associated with hepatitis E virus (HEV) as a food-borne pathogen. EFSA J. 2017, 15. [Google Scholar] [CrossRef]
- Ferri, G.; Vergara, A. Hepatitis E Virus in the Food of Animal Origin. Foodborne Pathog. Dis. 2021. [Google Scholar] [CrossRef]
- Lange, H.; Overbo, J.; Borgen, K.; Dudman, S.; Hoddevik, G.; Urdahl, A.M.; Vold, L.; Sjurseth, S.K. Hepatitis e in Norway: Seroprevalence in humans and swine. Epidemiol. Infect. 2017, 145, 181–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, J.; Mesquita, J.R.; Pereira, S.S.; Oliveira, R.M.S.; Abreu-Silva, J.; Rodrigues, A.; Myrmel, M.; Stene-Johansen, K.; Overbo, J.; Goncalves, G.; et al. Prevalence of hepatitis E virus antibodies in workers occupationally exposed to swine in Portugal. Med. Microbiol. Immunol. 2017, 206, 77–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Gageldonk-Lafeber, A.B.; van der Hoek, W.; Borlée, F.; Heederik, D.J.J.; Mooi, S.H.; Maassen, C.B.M.; Yzermans, C.J.; Rockx, B.; Smit, L.A.M.; Reimerink, J.H.J. Hepatitis E virus seroprevalence among the general population in a livestock-dense area in the Netherlands: A cross-sectional population-based serological survey. BMC Infect. Dis. 2017, 17, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Bartolo, I.; Ponterio, E.; Castellini, L.; Ostanello, F.; Ruggeri, F.M. Viral and antibody HEV prevalence in swine at slaughterhouse in Italy. Vet. Microbiol. 2011, 149, 330–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berto, A.; Martelli, F.; Grierson, S.; Banks, M. Hepatitis e virus in pork food chain, United Kingdom, 2009–2010. Emerg. Infect. Dis. 2012, 18, 1358–1360. [Google Scholar] [CrossRef] [PubMed]
- Di Bartolo, I.; Diez-Valcarce, M.; Vasickova, P.; Kralik, P.; Hernandez, M.; Angeloni, G.; Ostanello, F.; Bouwknegt, M.; Rodríguez-Lázaro, D.; Pavlik, I.; et al. Hepatitis E virus in pork production chain in Czech Republic, Italy, and Spain, 2010. Emerg. Infect. Dis. 2012, 18, 1282–1289. [Google Scholar] [CrossRef] [PubMed]
- Chelli, E.; Suffredini, E.; De Santis, P.; De Medici, D.; Di Bella, S.; D’amato, S.; Gucciardi, F.; Guercio, A.; Ostanello, F.; Perrone, V.; et al. Hepatitis E Virus Occurrence in Pigs Slaughtered in Italy. Animals 2021, 11, 277. [Google Scholar] [CrossRef]
- Sooryanarain, H.; Heffron, C.L.; Hill, D.E.; Fredericks, J.; Rosenthal, B.M.; Werre, S.R.; Opriessnig, T.; Meng, X.J. Hepatitis e virus in pigs from slaughterhouses, United States, 2017–2019. Emerg. Infect. Dis. 2020, 26, 354–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Intharasongkroh, D.; Sa-nguanmoo, P.; Tuanthap, S.; Thongmee, T.; Duang-in, A.; Klinfueng, S.; Chansaenroj, J.; Vongpunsawad, S.; Theamboonlers, A.; Payungporn, S.; et al. Hepatitis E Virus in Pork and Variety Meats Sold in Fresh Markets. Food Environ. Virol. 2017, 9, 45–53. [Google Scholar] [CrossRef]
- Leblanc, D.; Poitras, E.; Gagné, M.J.; Ward, P.; Houde, A. Hepatitis E virus load in swine organs and tissues at slaughterhouse determined by real-time RT-PCR. Int. J. Food Microbiol. 2010, 139, 206–209. [Google Scholar] [CrossRef]
- Nantel-Fortier, N.; Letellier, A.; Lachapelle, V.; Fravalo, P.; L’Homme, Y.; Brassard, J. Detection and Phylogenetic Analysis of the Hepatitis E Virus in a Canadian Swine Production Network. Food Environ. Virol. 2016, 8, 296–304. [Google Scholar] [CrossRef]
- Wichmann, O.; Schimanski, S.; Koch, J.; Kohler, M.; Rothe, C.; Plentz, A.; Jilg, W.; Stark, K. Phylogenetic and case-control study on hepatitis E virus infection in Germany. J. Infect. Dis. 2008, 198, 1732–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozyra, I.; Jabłoński, A.; Bigoraj, E.; Rzeżutka, A. Wild boar as a sylvatic reservoir of hepatitis E virus in Poland: A cross-sectional population study. Viruses 2020, 12, 1113. [Google Scholar] [CrossRef] [PubMed]
- Izopet, J.; Tremeaux, P.; Marion, O.; Migueres, M.; Capelli, N.; Chapuy-Regaud, S.; Mansuy, J.M.; Abravanel, F.; Kamar, N.; Lhomme, S. Hepatitis E virus infections in Europe. J. Clin. Virol. 2019, 120, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Juarez, A.; Frias, M.; Martinez-Peinado, A.; Risalde, M.A.; Rodriguez-Cano, D.; Camacho, A.; García-Bocanegra, I.; Cuenca-Lopez, F.; Gomez-Villamandos, J.C.; Rivero, A. Familial Hepatitis E Outbreak Linked to Wild Boar Meat Consumption. Zoonoses Public Health 2017, 64, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Sommerkorn, F.M.; Schauer, B.; Schreiner, T.; Fickenscher, H.; Krumbholz, A. Performance of Hepatitis E Virus (HEV)-antibody tests: A comparative analysis based on samples from individuals with direct contact to domestic pigs or wild boar in Germany. Med. Microbiol. Immunol. 2017, 206, 277–286. [Google Scholar] [CrossRef]
- Di Pasquale, S.; De Santis, P.; La Rosa, G.; Di Domenico, K.; Iaconelli, M.; Micarelli, G.; Martini, E.; Bilei, S.; De Medici, D.; Suffredini, E. Quantification and genetic diversity of Hepatitis E virus in wild boar (Sus scrofa) hunted for domestic consumption in Central Italy. Food Microbiol. 2019, 82, 194–201. [Google Scholar] [CrossRef]
- Mazzei, M.; Nardini, R.; Verin, R.; Forzan, M.; Poli, A.; Tolari, F. Serologic and molecular survey for hepatitis E virus in wild boar (Sus scrofa) in Central Italy. New Microbes New Infect. 2015, 7, 41–47. [Google Scholar] [CrossRef]
- Dalton, H.R.; Seghatchian, J. Hepatitis E virus: Emerging from the shadows in developed countries. Transfus. Apher. Sci. 2016, 55, 271–274. [Google Scholar] [CrossRef] [PubMed]
- de Deus, N.; Peralta, B.; Pina, S.; Allepuz, A.; Mateu, E.; Vidal, D.; Ruiz-Fons, F.; Martín, M.; Gortázar, C.; Segalés, J. Epidemiological study of hepatitis E virus infection in European wild boars (Sus scrofa) in Spain. Vet. Microbiol. 2008, 129, 163–170. [Google Scholar] [CrossRef]
- Boadella, M.; Ruiz-Fons, J.F.; Vicente, J.; Martín, M.; Segalés, J.; Gortazar, C. Seroprevalence evolution of selected pathogens in iberian wild boar. Transbound. Emerg. Dis. 2012, 59, 395–404. [Google Scholar] [CrossRef]
- Lewis, H.C.; Wichmann, O.; Duizer, E. Transmission routes and risk factors for autochthonous hepatitis E virus infection in Europe: A systematic review. Epidemiol. Infect. 2010, 138, 145–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schielke, A.; Ibrahim, V.; Czogiel, I.; Faber, M.; Schrader, C.; Dremsek, P.; Ulrich, R.G.; Johne, R. Hepatitis E virus antibody prevalence in hunters from a district in Central Germany, 2013: A cross-sectional study providing evidence for the benefit of protective gloves during disembowelling of wild boars. BMC Infect. Dis. 2015, 15, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Sáez-Royuela, C.; Gomariz, R.P.L.T. Age Determination of European Wild Boar Wildl. Soc. Bull. 1989, 17, 326–329. [Google Scholar]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods 2006, 131, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H.; Ling, R.; Li, H.; Harrison, T.J. The complete sequence of hepatitis E virus genotype 4 reveals an alternative strategy for translation of open reading frames 2 and 3. J. Gen. Virol. 2000, 81, 1675–1686. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Pittiglio, C.; Khomenko, S.; Beltran-Alcrudo, D. Wild boar mapping using population-density statistics: From polygons to high resolution raster maps. PLoS ONE 2018, 13, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Mughini-Gras, L.; Angeloni, G.; Salata, C.; Vonesch, N.; D’Amico, W.; Campagna, G.; Natale, A.; Zuliani, F.; Ceglie, L.; Monne, I.; et al. Hepatitis e virus infection in North Italy: High seroprevalence in swine herds and increased risk for swine workers. Epidemiol. Infect. 2017, 145, 3375–3384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spada, E.; Pupella, S.; Pisani, G.; Bruni, R.; Chionne, P.; Madonna, E.; Villano, U.; Simeoni, M.; Fabi, S.; Marano, G.; et al. A nationwide retrospective study on prevalence of hepatitis E virus infection in Italian blood donors. Blood Transfus. 2018, 16, 413–421. [Google Scholar]



{kind=link}
{kind=link}
| Animal | Age Category | Sex | Hunting Area | Slaughtering Date | RT-qPCR Viral Load | |
|---|---|---|---|---|---|---|
| Liver | Swab | |||||
| 1 | Adult | F | Semproniano GR | 11 February 2021 | 4.28 × 102 | - |
| 2 | Adult | F | 1.52 × 101 | - | ||
| 3 | Adult | M | 1.13 × 101 | - | ||
| 4 | Adult | M | 5.37 × 101 | - | ||
| 5 | Adult | F | Orbetello GR | 19 February 2021 | - | 3.84 × 101 |
| 6 | Adult | F | - | 9.9 × 102 | ||
| 7 | Sub-adult | M | Roccalbegna GR | 25 February 2021 | 1.31 × 103 | 1.42 × 102 |
| 8 | Sub-adult | M | 6.12 × 102 | 5.16 × 103 | ||
| 9 | Adult | F | 7.33 × 103 | - | ||
| 10 | Adult | M | Vicchi PO | 31 February 2021 | 2.08 × 103 | - |
| Mean | 1.48 × 103 | 1.57 × 103 | ||||
| Standard Deviation | 2.47 × 103 | 2.42 × 103 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Forzan, M.; Pacini, M.I.; Periccioli, M.; Mazzei, M. Hepatitis E Virus RNA Presence in Wild Boar Carcasses at Slaughterhouses in Italy. Animals 2021, 11, 1624. https://doi.org/10.3390/ani11061624
Forzan M, Pacini MI, Periccioli M, Mazzei M. Hepatitis E Virus RNA Presence in Wild Boar Carcasses at Slaughterhouses in Italy. Animals. 2021; 11(6):1624. https://doi.org/10.3390/ani11061624
Chicago/Turabian StyleForzan, Mario, Maria Irene Pacini, Marcello Periccioli, and Maurizio Mazzei. 2021. "Hepatitis E Virus RNA Presence in Wild Boar Carcasses at Slaughterhouses in Italy" Animals 11, no. 6: 1624. https://doi.org/10.3390/ani11061624
APA StyleForzan, M., Pacini, M. I., Periccioli, M., & Mazzei, M. (2021). Hepatitis E Virus RNA Presence in Wild Boar Carcasses at Slaughterhouses in Italy. Animals, 11(6), 1624. https://doi.org/10.3390/ani11061624