
Characterization of Foot and Mouth Disease Virus Serotype SAT-2 in Swamp Water Buffaloes (Bubalus bubalis) under the Egyptian Smallholder Production System

 , ,
, ,  ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population and Sampling
2.2. Nucleic Acid Extraction and RT-PCR
2.3. DNA Sequencing and Phylogenetic Analysis
2.4. Biochemical Analysis
2.5. Histopathological Examination
2.6. Data Analysis
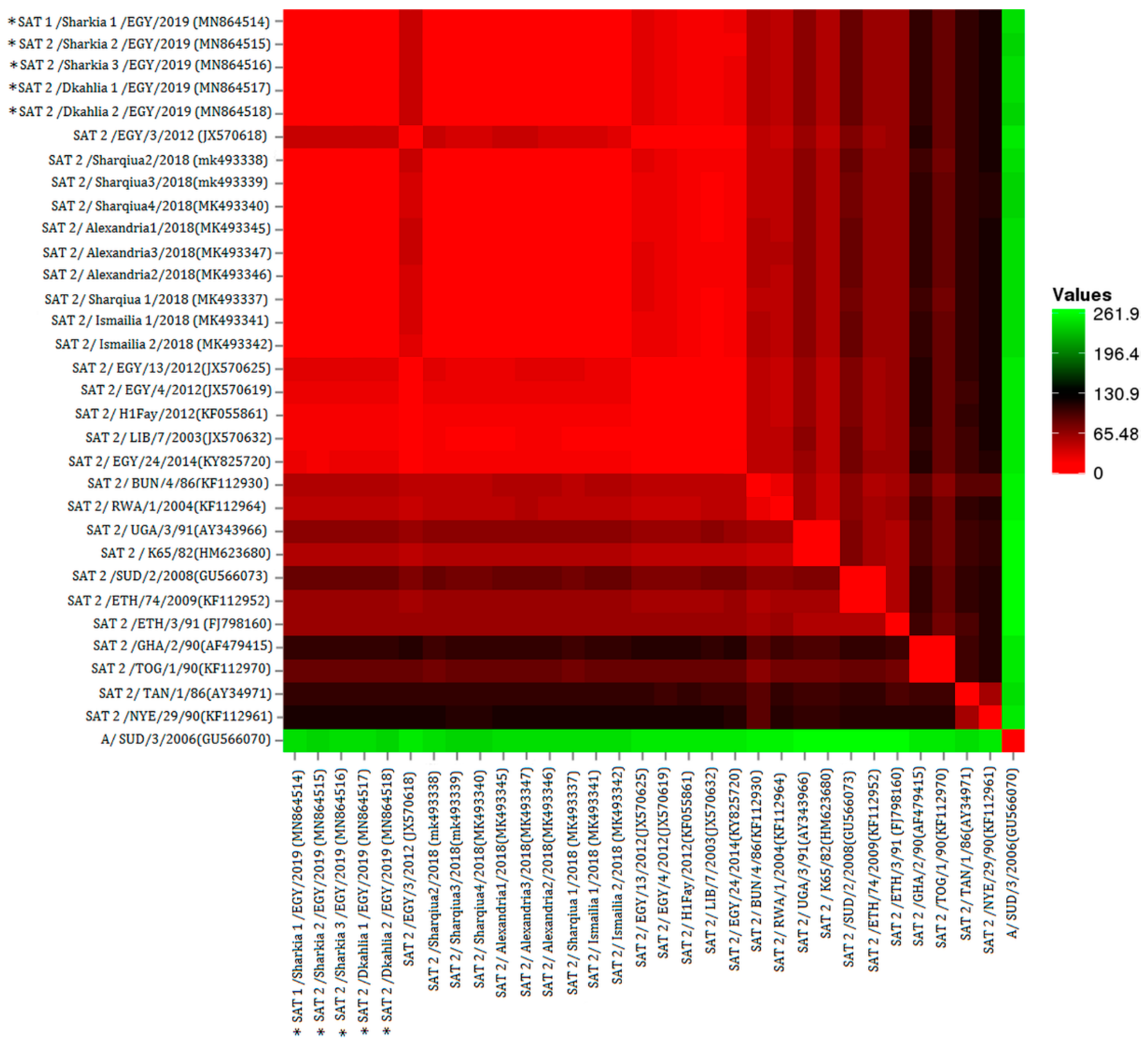
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lubroth, J. Foot-and-mouth disease. A review for the practitioner. Vet. Clin. N. Am. Food Anim. Pract. 2002, 18, 475–499. [Google Scholar] [CrossRef]
- Knight-Jones, T.; Rushton, J. The economic impacts of foot and mouth disease–What are they, how big are they and where do they occur? Prev. Vet. Med. 2013, 112, 161–173. [Google Scholar] [CrossRef] [Green Version]
- Reid, S.M.; Grierson, S.S.; Ferris, N.P.; Hutchings, G.H.; Alexandersen, S. Evaluation of automated RT-PCR to accelerate the laboratory diagnosis of foot-and-mouth disease virus. J. Virol. Methods 2003, 107, 129–139. [Google Scholar] [CrossRef]
- Knowles, N.; Nazem Shirazi, M.; Wadsworth, J.; Swabey, K.; Stirling, J.; Statham, R.; Li, Y.; Hutchings, G.; Ferris, N.; Parlak, Ü. Recent spread of a new strain (A-Iran-05) of foot-and-mouth disease virus type A in the Middle East. Transbound. Emerg. Dis. 2009, 56, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Jamal, S.M.; Ferrari, G.; Ahmed, S.; Normann, P.; Belsham, G.J. Molecular characterization of serotype Asia-1 foot-and-mouth disease viruses in Pakistan and Afghanistan; emergence of a new genetic Group and evidence for a novel recombinant virus. Infect. Genet. Evol. 2011, 11, 2049–2062. [Google Scholar] [CrossRef]
- Ahmed, H.A.; Salem, S.A.H.; Habashi, A.R.; Arafa, A.A.; Aggour, M.G.A.; Salem, G.H.; Gaber, A.S.; Selem, O.; Abdelkader, S.H.; Knowles, N.J.; et al. Emergence of Foot-and-Mouth Disease Virus SAT 2 in Egypt During 2012. Transbound. Emerg. Dis. 2012, 59, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Elhaig, M.M.; Elsheery, M.N. Molecular investigation of foot-and-mouth disease virus in domestic bovids from Gharbia, Egypt. Trop. Anim. Health Prod. 2014, 46, 1455–1462. [Google Scholar] [CrossRef]
- Mahmoud, E.A.; Neamat-Allah, A.N. Hemato-Biochemical Studies on Egyptian Buffaloes and Calves Naturally Infected with Foot and Mouth Disease Virus Serotype SAT2. Bull. Univ. Agric. Sci. Vet. Med. Cluj Napoca Vet. Med. 2016, 73, 230. [Google Scholar]
- El-Deen, N.; Neamat-Allah, A.; Rizk, L.G.; Fareed, R.S.G. Serological, hematological, biochemical and oxidative markers during foot and mouth disease serotype ‘O’infection, Egypt. Bull. Univ. Agric. Sci. Vet. Med. Cluj Napoca Vet. Med. 2017, 74, 218–226. [Google Scholar]
- Al-Hosary, A.A.; Kandeil, A.; El-Taweel, A.N.; Nordengrahn, A.; Merza, M.; Badra, R.; Kayali, G.; Ali, M.A. Co-infection with different serotypes of FMDV in vaccinated cattle in Southern Egypt. Virus Genes 2019, 55, 304–313. [Google Scholar] [CrossRef]
- Soltan, M.A.; Bazid, A.; Fawzy, M.; Wasfy, M.; Soliman, S.; Shahein, M.; El-Sayed, M. Genetic Characterization of Foot and Mouth Disease Virus (FMD) Serotypes in Egypt (2016–2017) and Identification of a New Lineage of Serotype O Topotype EA-3. Pak. Vet. J. 2019, 39. [Google Scholar] [CrossRef]
- Soltan, M.A.; Dohreig, R.M.; Abbas, H.; Ellawa, M.; Yousif, I.; Aly, A.E.; Wasfy, M.; El-Sayed, M.M. Emergence of foot and mouth disease virus, Lib-12 lineage of topotype VII, serotype SAT 2 in Egypt, 2018. Transbound. Emerg. Dis. 2019, 66, 1105–1106. [Google Scholar] [CrossRef] [PubMed]
- Baluka, S.A. Economic effects of foot and mouth disease outbreaks along the cattle marketing chain in Uganda. Vet. World 2016, 9, 544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knowles, N.; Samuel, A. Molecular epidemiology of foot-and-mouth disease virus. Virus Res. 2003, 91, 65–80. [Google Scholar] [CrossRef]
- El-Shehawy, L.I.; Abu-Elnaga, H.I.; Rizk, S.A.; Abd El-Kreem, A.S.; Mohamed, A.; Fawzy, H.G. Molecular differentiation and phylogenetic analysis of the Egyptian foot-and-mouth disease virus SAT2. Arch. Virol. 2014, 159, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Soltan, M.A.; Negmaldin, A.H.; El-Diasty, M.M.; Mansour, S.M.; Elbadry, M.A.; Wilkes, R.P. Molecular characterization of circulating Foot and mouth disease virus (FMDV) serotype O topotype EA-3 and serotype A (African topotype) genotype IV in Egypt, 2016. Vet. Microbiol. 2017, 208, 89–93. [Google Scholar] [CrossRef]
- Sulayeman, M.; Dawo, F.; Mammo, B.; Gizaw, D.; Shegu, D. Isolation, molecular characterization and sero-prevalence study of foot-and-mouth disease virus circulating in central Ethiopia. BMC Vet. Res. 2018, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- El Damaty, H.M.; Mahmmod, Y.S.; Gouda, S.M.; Sobhy, N.M. Epidemiological and ultrasonographic investigation of bovine fascioliasis in smallholder production system in Eastern Nile Delta of Egypt. Prev. Vet. Med. 2018, 158, 35–42. [Google Scholar] [CrossRef]
- Constable, P.; Hinchcliff, K.; Done, S.; Grünberg, W. Clinical Examination and Making A Diagnosis. In Veterinary Medicine, 11th ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–28. [Google Scholar]
- World Organization for Animal Health (OIE). Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Foot and Mouth Disease (Infection with Foot and Mouth Disease Virus) CHAPTER 3.1.8. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.01.08_FMD.pdf (accessed on 29 February 2020).
- Kandeil, A.; El-Shesheny, R.; Kayali, G.; Moatasim, Y.; Bagato, O.; Darwish, M.; Gaffar, A.; Younes, A.; Farag, T.; Kutkat, M.A.; et al. Characterization of the recent outbreak of foot-and-mouth disease virus serotype SAT2 in Egypt. Arch. Virol. 2013, 158, 619–627. [Google Scholar] [CrossRef]
- Othman, O.E.; Khodary, M.G.; El-Deeb, A.H.; Hussein, H.A. Five BoLA-DRB3 genotypes detected in Egyptian buffalo infected with Foot and Mouth disease virus serotype O. J. Genet. Eng. Biotechnol. 2018, 16, 513–518. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Guzy, P.M. Creatine phosphokinase-MB (CPK-MB) and the diagnosis of myocardial infarction. West. J. Med. 1977, 127, 455. [Google Scholar]
- Skinner, J.; Brown, R.; Roberts, L. Bovine haptoglobin response in clinically defined field conditions. Vet. Rec. 1991, 128, 147–149. [Google Scholar] [CrossRef]
- Hussein, H.A.; Staufenbiel, R. Analytical validation of bovine plasma ceruloplasmin measurement by p-phenylenediamine oxidation and effect of storage time and freezing temperature. Acta Vet. Scand. 2017, 59, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordatos, K. Theory and practice of histological techniques. Pathology 2002, 34, 384. [Google Scholar] [CrossRef]
- Chong, J.; Xia, J. MetaboAnalystR: An R package for flexible and reproducible analysis of metabolomics data. Bioinformatics 2018, 34, 4313–4314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Aly, M.; Nayel, M.; Salama, A.; Ghazy, E.; Elshahawy, I. Cardiac troponin I as a cardiac biomarker has prognostic and predictive value for poor survival in Egyptian buffalo calves with foot-and-mouth disease. Vet. World 2020, 13, 890–895. [Google Scholar] [CrossRef]
- Karapinar, T.; Dabak, D.O.; Kuloglu, T.; Bulut, H. High cardiac troponin I plasma concentration in a calf with myocarditis. Can. Vet. J. 2010, 51, 397–399. [Google Scholar]
- Ghuman, S.; Kumar, A.; Honparkhe, M. A preliminary study on plasma haptoglobin concentrations in buffaloes following spontaneous or assisted delivery. Buffalo Bull. 2009, 28, 127–130. [Google Scholar]
- Liu, X.; Shao, Y. A study on the normal levels of ceruloplasmin (Cp) in serum of domestic animals. AGRIS 1988, 10, 16–18. [Google Scholar]
- Knowles, N.J.; Wadsworth, J.; Reid, S.M.; Swabey, K.G.; El-Kholy, A.A.; Abd El-Rahman, A.O.; Soliman, H.M.; Ebert, K.; Ferris, N.P.; Hutchings, G.H. Foot-and-mouth disease virus serotype A in Egypt. Emerg. Infect. Dis. 2007, 13, 1593. [Google Scholar] [CrossRef] [PubMed]
- Nampanya, S.; Richards, J.; Khounsy, S.; Inthavong, P.; Yang, M.; Rast, L.; Windsor, P. Investigation of foot and mouth disease hotspots in northern Lao PDR. Transbound. Emerg. Dis. 2013, 60, 315–329. [Google Scholar] [CrossRef]
- Diab, E.; Bazid, A.-H.I.; Fawzy, M.; El-Ashmawy, W.R.; Fayed, A.A.; El-Sayed, M.M. Foot-and-mouth disease outbreaks in Egypt during 2013-2014: Molecular characterization of serotypes A, O, and SAT2. Vet. World 2019, 12, 190. [Google Scholar] [CrossRef] [Green Version]
- Arzt, J.; Baxt, B.; Grubman, M.J.; Jackson, T.; Juleff, N.; Rhyan, J.; Rieder, E.; Waters, R.; Rodriguez, L.L. The Pathogenesis of Foot-and-Mouth Disease II: Viral Pathways in Swine, Small Ruminants, and Wildlife; Myotropism, Chronic Syndromes, and Molecular Virus–Host Interactions. Transbound. Emerg. Dis. 2011, 58, 305–326. [Google Scholar] [CrossRef]
- Sobhy, N.M.; Bayoumi, Y.H.; Mor, S.K.; El-Zahar, H.I.; Goyal, S.M. Outbreaks of foot and mouth disease in Egypt: Molecular epidemiology, evolution and cardiac biomarkers prognostic significance. Int. J. Vet. Sci. Med. 2018, 6, 22–30. [Google Scholar] [CrossRef] [Green Version]
- David Eckersall, P. Chapter 5-Proteins, Proteomics, and the Dysproteinemias A2-Kaneko. In Clinical Biochemistry of Domestic Animals, 6th ed.; Kaneko, J.J., Harvey, J.W., Bruss, M.L., Eds.; Academic Press: San Diego CA, USA, 2008; pp. 117–155. [Google Scholar]
- Cassat, J.E.; Skaar, E.P. Iron in infection and immunity. Cell Host Microbe 2013, 13, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Parrow, N.L.; Fleming, R.E.; Minnick, M.F. Sequestration and scavenging of iron in infection. Infect. Immun. 2013, 81, 3503–3514. [Google Scholar] [CrossRef] [Green Version]
- Byomi, A.M. SAT2 Foot and Mouth Disease (FMD) outbreak in a mixed farm in Egypt. Berliner Münch. Tierärztl. Wochenschr. 2015, 128, 188–192. [Google Scholar]




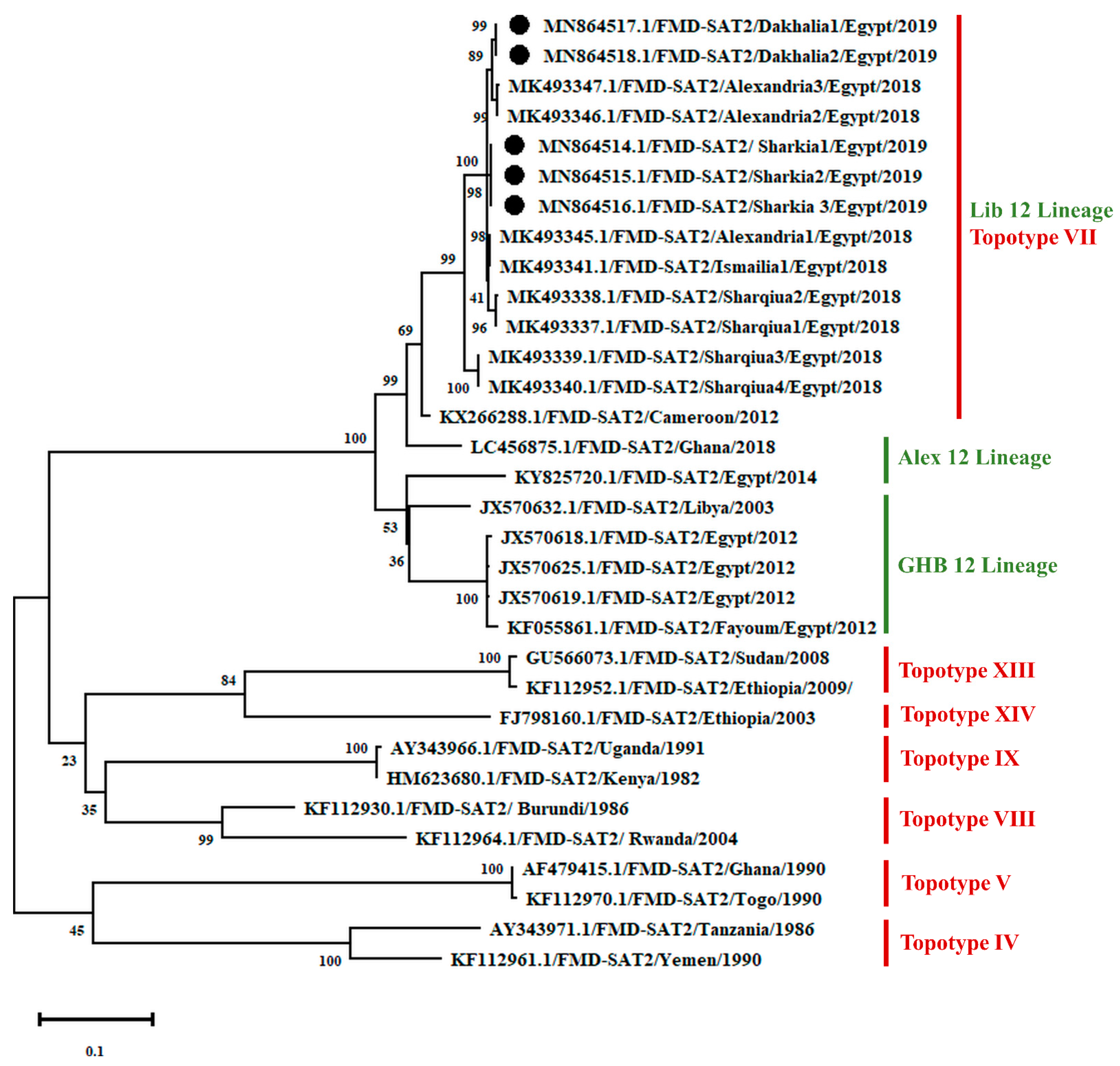{kind=link}
{kind=link}
{kind=link}
| Serotype | Target Genes | Primer Sequence (5′–3′) | Length of Amplified Product (bp) | Reference |
|---|---|---|---|---|
| FMD | 5′UTR | GCCTGGTCTTTCCAGGTCT | 326 | [20] |
| CCAGTCCCCTTCTCAGATC | ||||
| FMD A | VP3-2B | TACCAAATTACACACGGGAA | 863–866 | [21] |
| GACATGTCCTCCTGCATCTG | ||||
| FMD O | VP3-2B | ACCAACCTCCTTGATGTGGCT | 1301 | |
| GACATGTCCTCCTGCATCTG | ||||
| FMD SAT-2 | VP3-2B | TGAACTACCACTTCATGTACACAG | 1279 | [22] |
| ACAGCGGCCATGCACGACAG |
| Parameters | Standard Concentration Ranges | Mean (±SE) | References |
|---|---|---|---|
| cTnI (µg/L) | 0.021–0.12 | 0.61 ± 0.02 | [31,32] |
| CK-MB (U/L) | 100.56–165.64 | 263.73± 5.56 | [31] |
| Haptoglobin (g/dL) | 0.191–0.260 | 0.493 ± 0.051 | [33] |
| Ceruloplasmin (mg/dL) | 8.65–10.86 | 17.72 ± 1.23 | [34] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Damaty, H.M.; Fawzi, E.M.; Neamat-Allah, A.N.F.; Elsohaby, I.; Abdallah, A.; Farag, G.K.; El-Shazly, Y.A.; Mahmmod, Y.S. Characterization of Foot and Mouth Disease Virus Serotype SAT-2 in Swamp Water Buffaloes (Bubalus bubalis) under the Egyptian Smallholder Production System. Animals 2021, 11, 1697. https://doi.org/10.3390/ani11061697
El Damaty HM, Fawzi EM, Neamat-Allah ANF, Elsohaby I, Abdallah A, Farag GK, El-Shazly YA, Mahmmod YS. Characterization of Foot and Mouth Disease Virus Serotype SAT-2 in Swamp Water Buffaloes (Bubalus bubalis) under the Egyptian Smallholder Production System. Animals. 2021; 11(6):1697. https://doi.org/10.3390/ani11061697
Chicago/Turabian StyleEl Damaty, Hend M., Elshaima M. Fawzi, Ahmed N. F. Neamat-Allah, Ibrahim Elsohaby, Abdelmonem Abdallah, Gamelat K. Farag, Yousry A. El-Shazly, and Yasser S. Mahmmod. 2021. "Characterization of Foot and Mouth Disease Virus Serotype SAT-2 in Swamp Water Buffaloes (Bubalus bubalis) under the Egyptian Smallholder Production System" Animals 11, no. 6: 1697. https://doi.org/10.3390/ani11061697
APA StyleEl Damaty, H. M., Fawzi, E. M., Neamat-Allah, A. N. F., Elsohaby, I., Abdallah, A., Farag, G. K., El-Shazly, Y. A., & Mahmmod, Y. S. (2021). Characterization of Foot and Mouth Disease Virus Serotype SAT-2 in Swamp Water Buffaloes (Bubalus bubalis) under the Egyptian Smallholder Production System. Animals, 11(6), 1697. https://doi.org/10.3390/ani11061697