
Expression of Transforming Growth Factor Beta Isoforms in Canine Endometrium with Cystic Endometrial Hyperplasia–Pyometra Complex

 , ,
, ,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Uteri Sample Collections and Classification of the Study Groups
2.1.1. Animals
2.1.2. Clinical Evaluation of Animals
2.2. Hematoxylin and Eosin Staining of Tissue Sections and Histological Examination
2.3. RNA Isolation and cDNA Synthesis
2.4. Real-Time PCR Quantification
2.5. Western Blotting
2.6. Statistical Analysis
3. Results
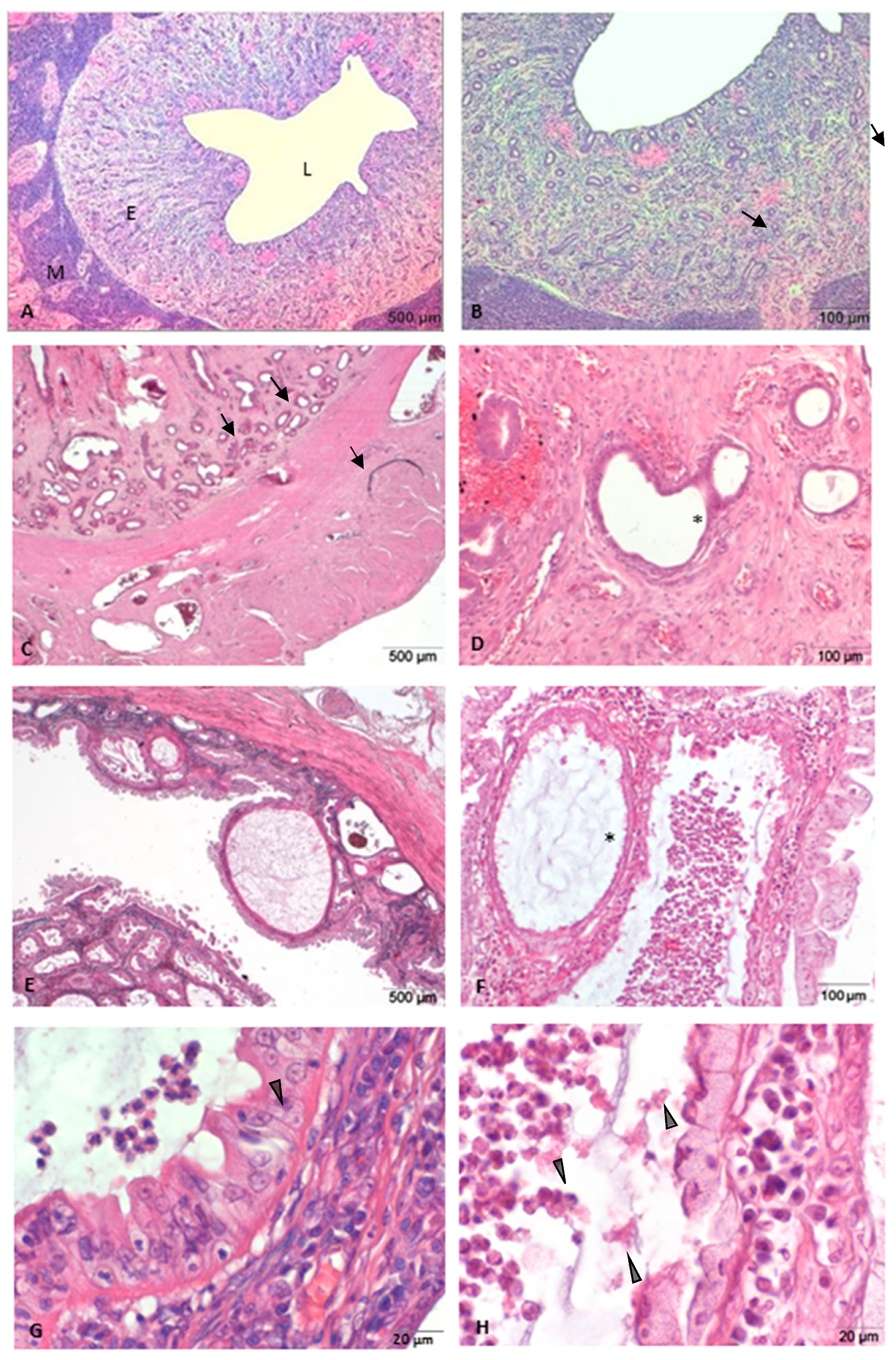
3.1. Histopathological Analysis of Uteri: Normal and with Morphological Abnormalities
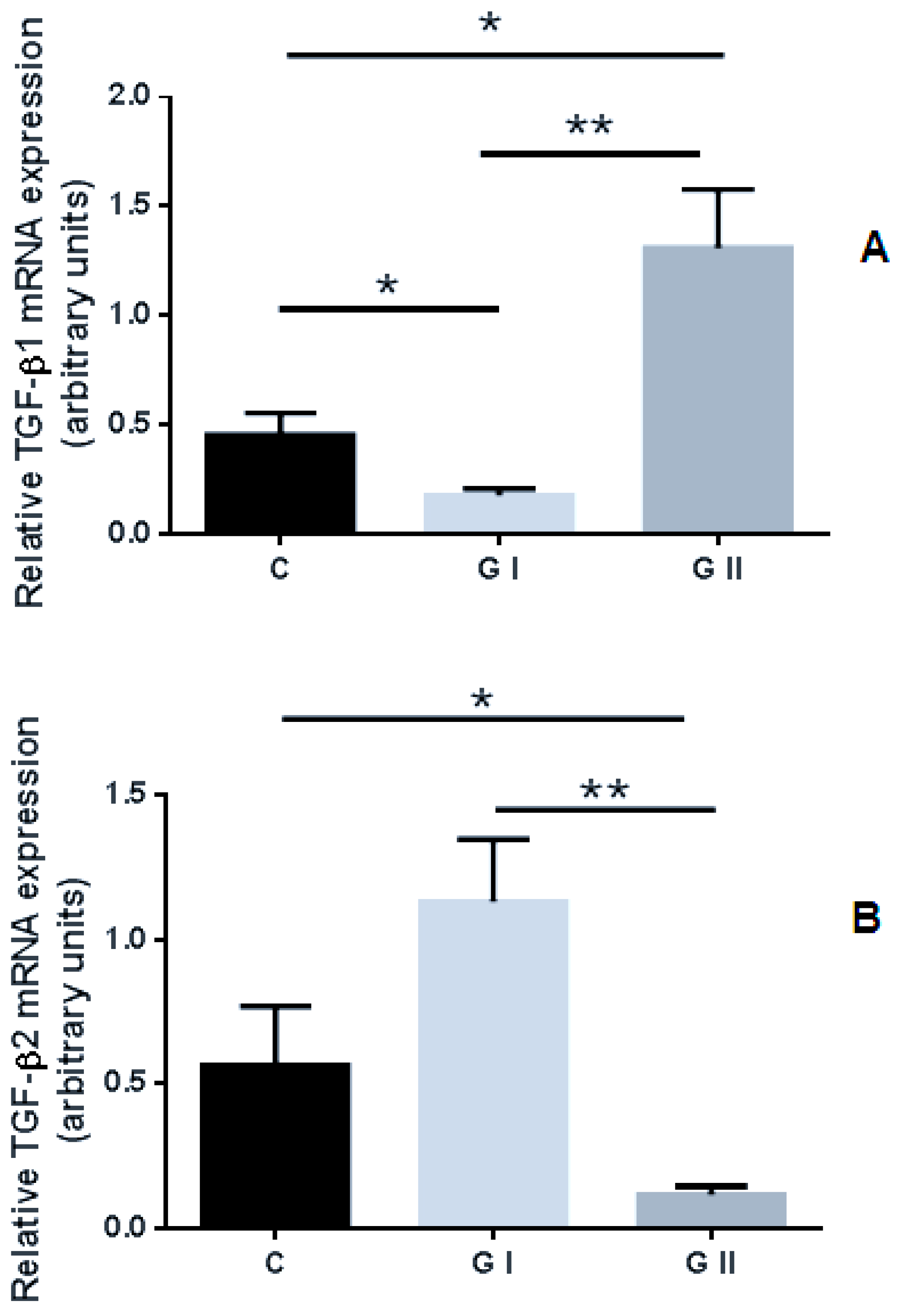
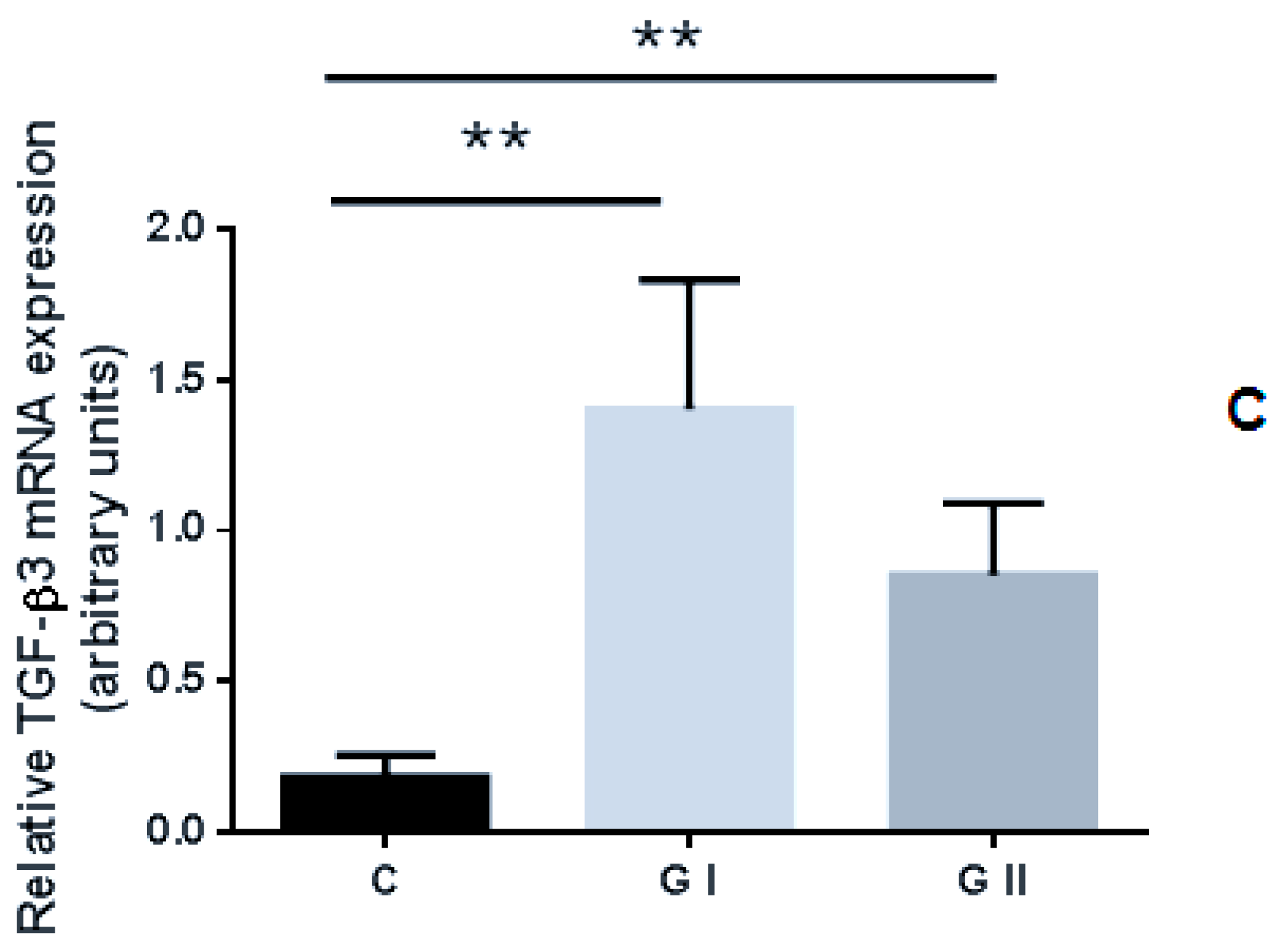
3.2. mRNA Expression Profile of TGF-βs in the Canine Endometrium
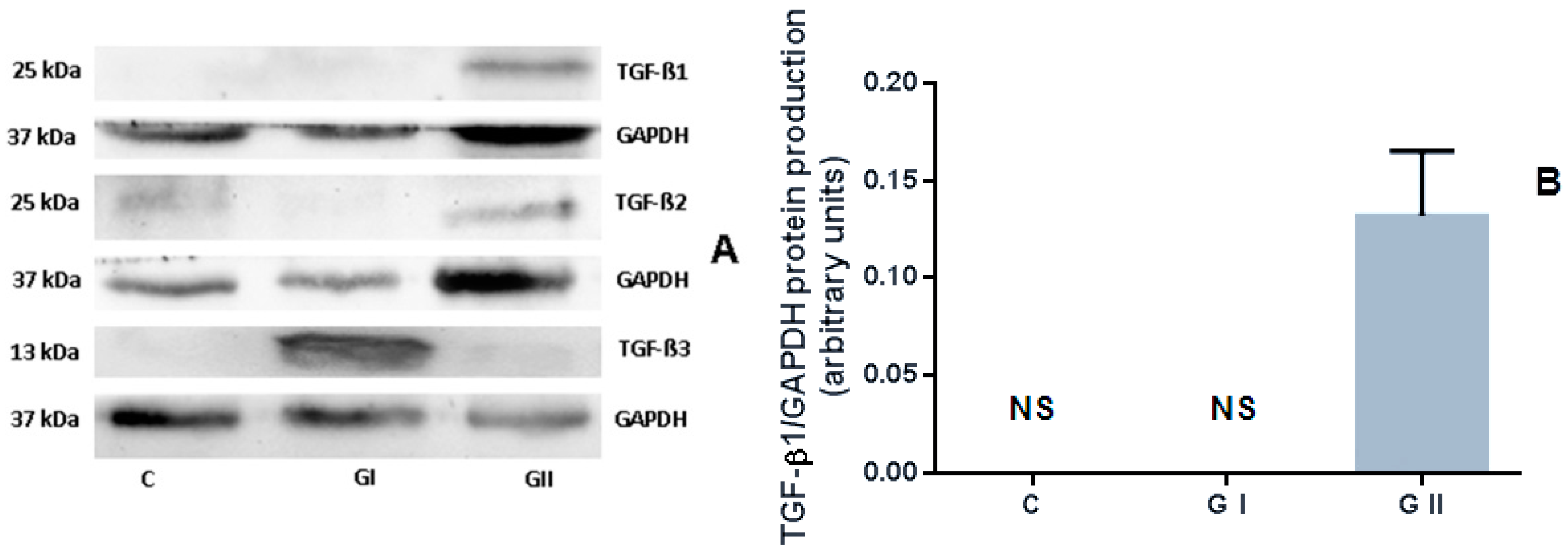
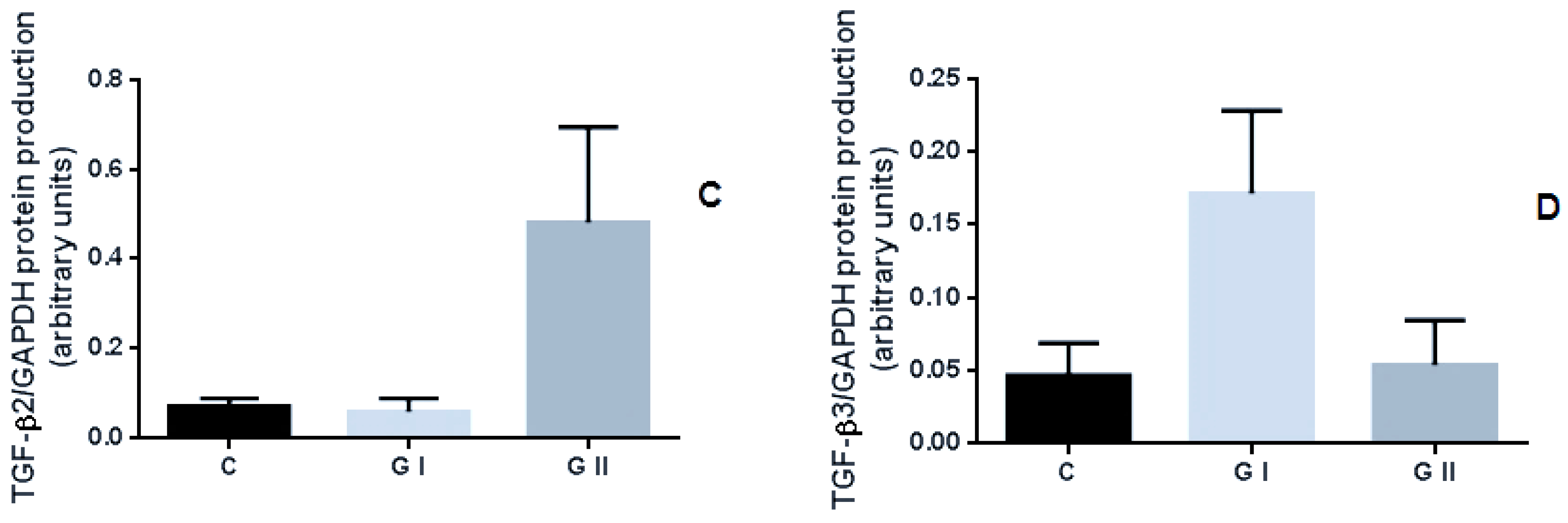
3.3. The Protein Production of TGF-β Isoforms in the Canine Endometrium
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BMP-2 | Bone morphogenetic protein 2 |
| CE | Chronic endometritis |
| CEH | Cystic endometrial hyperplasia |
| CL | Corpus luteum |
| ECM | Extracellular matrix |
| FGF10 | Fibroblast growth factor 10 |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| Gf | Growth factors |
| IL | Interleukin |
| NK | Natural killer |
| P4 | Progesterone |
| PF | Peritoneal fluid |
| PGF2α | Prostaglandin F2α |
| PI3-K/Akt | Phosphoinositide-3-kinase/Protein kinase B |
| RIF | Repeated implantation failure |
| TGF-β | Transforming growth factor beta |
| Tgfbr1 | Transforming growth factor beta receptor1 |
| UFs | Uterine fibroids |
References
- Bigliardi, E.; Parmigiani, E.; Cavirani, S.; Luppi, A.; Bonati, L.; Corradi, A. Ultrasonography and cystic hyperplasia-pyometra complex in the bitch. Reprod. Domest. Anim. 2004, 39, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Radwińska, J.; Domosawska, A.; Pomianowski, A.; Zarczyńska, K.; Jurczak, A. Implications of blood coagulation and fibrinolytic disorders in severe endometritis-pyometra complex in bitches. Bull. Vet. Inst. Pulawy 2012, 56, 293–297. [Google Scholar] [CrossRef] [Green Version]
- Batista, P.R.; Gobello, C.; Rube, A.; Corrada, Y.A.; Tórtora, M.; Blanco, P.G. Uterine blood flow evaluation in bitches suffering from cystic endometrial hyperplasia (CEH) and CEH-pyometra complex. Theriogenology 2016, 85, 1258–1261. [Google Scholar] [CrossRef] [PubMed]
- Bukowska, D.; Kempisty, B.; Zawierucha, P.; Jopek, K.; Piotrowska, H.; Antosik, P.; Ciesiółka, S.; Woźna, M.; Brüssow, K.P.; Jaśkowski, J.M. Microarray analysis of inflammatory response-related gene expression in the uteri of dogs with pyometra. J. Biol. Regul. Homeost. Agents 2014, 28, 637–648. [Google Scholar] [PubMed]
- Hagman, R. Diagnostic and prognostic markers for uterine diseases in dogs. Reprod. Domest. Anim. 2014, 49, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Kida, K.; Maezono, Y.; Kawate, N.; Inaba, T.; Hatoya, S.; Tamada, H. Epidermal growth factor, transforming growth factor-α, and epidermal growth factor receptor expression and localization in the canine endometrium during the estrous cycle and in bitches with pyometra. Theriogenology 2010, 73, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Veiga, G.A.L.; Miziara, R.H.; Angrimani, D.S.R.; Papa, P.C.; Cogliati, B.; Vannucchi, C.I. Cystic endometrial hyperplasia-pyometra syndrome in bitches: Identification of hemodynamic, inflammatory, and cell proliferation changes. Biol. Reprod. 2017, 96, 58–69. [Google Scholar] [CrossRef] [Green Version]
- Giudice, L.C. Growth factors and growth modulators in human uterine endometrium: Their potential relevance to reproductive medicine. Fertil. Steril. 1994, 61, 1–17. [Google Scholar] [CrossRef]
- Govinden, R.; Bhoola, K.D. Genealogy, expression, and cellular function of transforming growth factor-β. Pharmacol. Ther. 2003, 98, 257–265. [Google Scholar] [CrossRef]
- Caron, P.L.; Fréchette-Frigon, G.; Shooner, C.; Leblanc, V.; Asselin, E. Transforming growth factor beta isoforms regulation of Akt activity and XIAP levels in rat endometrium during estrous cycle, in a model of pseudopregnancy and in cultured decidual cells. Reprod. Biol. Endocrinol. 2009, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bierie, B.; Moses, H.L. Tumour microenvironment-TGFΒ: The molecular Jekyll and Hyde of cancer. Nat. Rev. Cancer 2006, 6, 506–520. [Google Scholar] [CrossRef] [PubMed]
- Knight, P.G.; Glister, C. Local roles of TGF-β superfamily members in the control of ovarian follicle development. Anim. Reprod. Sci. 2003, 78, 165–183. [Google Scholar] [CrossRef]
- Gao, Y.; Li, S.; Li, Q. Uterine epithelial cell proliferation and endometrial hyperplasia: Evidence from a mouse model. Mol. Hum. Reprod. 2014, 20, 776–786. [Google Scholar] [CrossRef] [PubMed]
- Massagué, J. TGFβ signalling in context. Nat. Rev. Mol. Cell Biol. 2012, 13, 616–630. [Google Scholar] [CrossRef] [PubMed]
- Galvão, A.M.; Skarzynski, D.; Ferreira-Dias, G. Luteolysis and the Auto-, Paracrine Role of Cytokines from Tumor Necrosis Factor α and Transforming Growth Factor β Superfamilies. Vitam. Horm. 2018, 107, 287–315. [Google Scholar] [CrossRef]
- Hoffmann, B.; Büsges, F.; Engel, E.; Kowalewski, M.P.; Papa, P. Regulation of corpus luteum-function in the bitch. Reprod. Domest. Anim. 2004, 39, 232–240. [Google Scholar] [CrossRef]
- Mize, D.; Saikia, P.J.; Sarma, H.N. Expression of Insulin-like Growth Factor-I in Uterus is Ovarian Steroid Dependent: An in situ Immunohistochemical Study in Rat. J. Reprod. Contracept. 2012, 23, 222–236. [Google Scholar] [CrossRef]
- Jones, R.L.; Stoikos, C.; Findlay, J.K.; Salamonsen, L.A. TGF-β superfamily expression and actions in the endometrium and placenta. Reproduction 2006, 132, 217–232. [Google Scholar] [CrossRef]
- Chatzaki, E.; Kouimtzoglou, E.; Margioris, A.N.; Gravanis, A. Transforming growth factor β1 exerts an autocrine regulatory effect on human endometrial stromal cell apoptosis, involving the FasL and Bcl-2 apoptotic pathways. Mol. Hum. Reprod. 2003, 9, 91–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eritja, N.; Felip, I.; Dosil, M.A.; Vigezzi, L.; Mirantes, C.; Yeramian, A.; Navaridas, R.; Santacana, M.; Llobet-Navas, D.; Yoshimura, A.; et al. A Smad3-PTEN regulatory loop controls proliferation and apoptotic responses to TGF-β in mouse endometrium. Cell Death Differ. 2017, 24, 1443–1458. [Google Scholar] [CrossRef] [PubMed]
- Shooner, C.; Caron, P.-L.; Fréchette-Frigon, G.; Leblanc, V.; Déry, M.-C.; Asselin, E. TGF-beta expression during rat pregnancy and activity on decidual cell survival. Reprod. Biol. Endocrinol. 2005, 3, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bukowska, D.; Kempisty, B.; Jackowska, M.; Woźna, M.; Antosik, P.; Piotrowska, H.; Jaśkowski, J.M. Differential expression of epidermal growth factor and transforming growth factor beta isoforms in dog endometrium during different periods of the estrus cycle. Pol. J. Vet. Sci. 2011, 14, 259–264. [Google Scholar] [CrossRef] [Green Version]
- Hagman, R. Molecular aspects of uterine diseases in dogs. Reprod. Domest. Anim. 2017, 52, 37–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Bosschere, H.; Ducatelle, R.; Vermeirsch, H.; Van Den Broeck, W.; Coryn, M. Cystic endometrial hyperplasia-pyometra complex in the bitch: Should the two entities be disconnected? Theriogenology 2001, 55, 1509–1519. [Google Scholar] [CrossRef]
- Woźna-Wysocka, M.; Rybska, M.; Błaszak, B.; Jaśkowski, B.M.; Kulus, M.; Jaśkowski, J.M. Morphological changes in bitches endometrium affected by cystic endometrial hyperplasia–pyometra complex–the value of histopathological examination. BMC Vet. Res. 2021, 17, 174. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Antosik, P.; Kempisty, B.; Jackowska, M.; Bukowska, D.; Woźna, M.; Lianeri, M.; Brüssow, K.P.; Jaśkowski, J.M. Differential expression of genes encoding EGF, IGF-I, TGF |β1, TGF β2 and TGF β3 in porcine endometrium during estrus cycle at different ages. Med. Weter. 2010, 66, 618–621. [Google Scholar]
- Groppetti, D.; Pecile, A.; Arrighi, S.; Di Giancamillo, A.; Cremonesi, F. Endometrial cytology and computerized morphometric analysis of epithelial nuclei: A useful tool for reproductive diagnosis in the bitch. Theriogenology 2010, 73, 927–941. [Google Scholar] [CrossRef]
- Kelly, A.; Houston, S.A.; Sherwood, E.; Casulli, J.; Travis, M.A. Regulation of Innate and Adaptive Immunity by TGFβ. Adv. Immunol. 2017, 134, 137–233. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, H.; Chen, Z.; Zhang, W.; Liu, X.; Fang, J.; Liu, F.; Kwak-Kim, J. Endometrial TGF-β, IL-10, IL-17 and autophagy are dysregulated in women with recurrent implantation failure with chronic endometritis. Reprod. Biol. Endocrinol. 2019, 17, 2. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Duran, S.; Lydon, J.P.; DeMayo, F.J.; Burghardt, R.C.; Bayless, K.J.; Bartholin, L.; Li, Q. Constitutive activation of transforming growth factor beta receptor 1 in the mouse uterus impairs uterine morphology and function. Biol. Reprod. 2015, 92, 1–13. [Google Scholar] [CrossRef]
- Pizzo, A.; Salmeri, F.M.; Ardita, F.V.; Sofo, V.; Tripepi, M.; Marsico, S. Behaviour of Cytokine Levels in Serum and Peritoneal Fluid of Women with Endometriosis. Gynecol. Obstet. Invest. 2002, 54, 82–87. [Google Scholar] [CrossRef]
- Lee, H.J.; Kim, H.; Ku, S.-Y.; Kim, S.H.; Kim, J.G. Transforming Growth Factor-β1 Gene Polymorphisms in Korean Women With Endometriosis. Am. J. Reprod. Immunol. 2011, 66, 428–434. [Google Scholar] [CrossRef]
- Young, V.J.; Brown, J.K.; Saunders, P.T.K.; Duncan, W.C.; Horne, A.W. The Peritoneum Is Both a Source and Target of TGF-β in Women with Endometriosis. PLoS ONE 2014, 9, e106773. [Google Scholar]
- Kajdaniuk, D.; Marek, B.; Borgiel-Marek, H.; Kos-Kudła, B. Transforming growth factor beta1 (TGFbeta1) in physiology and pathology. Endokrynol. Pol. 2013, 64, 384–396. [Google Scholar] [CrossRef] [Green Version]
- Sikora, J.; Smycz-Kubańska, M.; Mielczarek-Palacz, A.; Bednarek, I.; Kondera-Anasz, Z. The involvement of multifunctional TGF-β and related cytokines in pathogenesis of endometriosis. Immunol. Lett. 2018, 201, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Marek, A.; Brodzicki, J.; Liberek, A.; Korzon, M. TGF-β (transforming growth factor-β) in chronic inflammatory conditions—A new diagnostic and prognostic marker? Med. Sci. Monit. 2002, 8, 145–152. [Google Scholar]
- Loverro, G.; Maiorano, E.; Napoli, A.; Selvaggi, L.; Marra, E.; Perlino, E. Transforming growth factor-beta 1 and insulin-like growth factor-1 expression in ovarian endometriotic cysts: A preliminary study. Int. J. Mol. Med. 2001, 7, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Liu, X.; Zhang, H.; Guo, S.W. Transforming growth factor β1 signaling coincides with epithelial-mesenchymal transition and fibroblast-to-myofibroblast transdifferentiation in the development of adenomyosis in mice. Hum. Reprod. 2016, 31, 355–369. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Kishi, Y.; Matsubara, S. Mechanisms Underlying Adenomyosis-Related Fibrogenesis. Gynecol. Obstet. Investig. 2020, 85, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.Y.; Ku, B.J.; Kim, T.H.; Il Ahn, J.; Ahn, J.Y.; Yang, W.S.; Lim, J.M.; Taketo, M.M.; Shin, J.H.; Jeong, J.W. β-catenin activates TGF-β-induced epithelial–mesenchymal transition in adenomyosis. Exp. Mol. Med. 2020, 52, 1754–1765. [Google Scholar] [CrossRef] [PubMed]
- Ciebiera, M.; Włodarczyk, M.; Wrzosek, M.; Męczekalski, B.; Nowicka, G.; Łukaszuk, K.; Ciebiera, M.; Słabuszewska-Jóźwiak, A.; Jakiel, G. Role of transforming growth factor β in uterine fibroid biology. Int. J. Mol. Sci. 2017, 18, 2435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norian, J.M.; Malik, M.; Parker, C.Y.; Joseph, D.; Leppert, P.C.; Segars, J.H.; Catherino, W.H. Transforming Growth Factor β3 regulates the versican variants in the extracellular matrix-rich uterine leiomyomas. Reprod. Sci. 2009, 16, 1153–1164. [Google Scholar] [CrossRef] [Green Version]
- Arici, A.; Sozen, I. Transforming growth factor-beta3 is expressed at high levels in leiomyoma where it stimulates fibronectin expression and cell proliferation. Fertil. Steril. 2000, 73, 1006–1011. [Google Scholar] [CrossRef]
- Thompson, N.L.; Flanders, K.C.; Smith, J.M.; Ellingsworth, L.R.; Roberts, A.B.; Sporn, M.B. Expression of transforming growth factor-beta 1 in specific cells and tissues of adult and neonatal mice. J. Cell Biol. 1989, 108, 661–669. [Google Scholar] [CrossRef]
- Okamura, T.; Morita, K.; Iwasaki, Y.; Inoue, M.; Komai, T.; Fujio, K.; Yamamoto, K. Role of TGF-β3 in the regulation of immune responses. Clin. Exp. Rheumatol. 2015, 33, S63–S69. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Control Group | Group I CEH | Group II CEH–Pyometra |
|---|---|---|---|
| Number of bitches | n = 20 | n = 20 | n = 20 |
| Age (years) | |||
| max | 3 | 10 | 10 |
| min | 1 | 3 | 2 |
| mean ± SD | 1.92 ± 0.97 | 6.4 ± 2.4 | 7.9 ± 2.2 |
| Body weight (kg) | |||
| max | 40 | 42 | 46 |
| min | 2 | 3 | 5 |
| mean ± SD | 19.8 ± 11.1 | 20.4 ± 9.8 | 18.4 ± 14.1 |
| Breed | |||
| crossbreed | 12 | 8 | 11 |
| in breed type | 8 | 12 | 9 |
| Gene Name | Primer Sequences 5′–3′ | Gene Accession No. | Catalog No. | Product Size (bp) |
|---|---|---|---|---|
| TGF-β1 | F: ATGAGCCCAAGGGTTACCA R: GTCCAGGCTCCAAATGTAGG | NM_001003309 | 04688546001 | 65 |
| TGF-β2 | F: GCAGCAAGACGATAATCACG R: TCTTGTCGCTGTCGTCCTC | XM_545713.3 | 04688678001 | 67 |
| TGF-β3 | F: CTGGCCCTTTACAACAGCAC R: CGACTCGGTGTTTTCCTGAG | DQ310186.1 | 04686993001 | 87 |
| GAPDH | F:GCTGGGGCTCACTTGAAA R:GTTCACGCCCATCACAAAC | NM_001003142.1 | 04688678001 | 87 |
| ACTB | F:CTGGACTTCGAGCAGGAGA R:CCGTCGGGTAGTTCGTAGC | AF021873.2 | 04689038001 | 73 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rybska, M.; Woźna-Wysocka, M.; Wąsowska, B.; Skrzypski, M.; Kubiak, M.; Błaszak, B.; Łukomska, A.; Nowak, T.; Jaśkowski, J.M. Expression of Transforming Growth Factor Beta Isoforms in Canine Endometrium with Cystic Endometrial Hyperplasia–Pyometra Complex. Animals 2021, 11, 1844. https://doi.org/10.3390/ani11061844
Rybska M, Woźna-Wysocka M, Wąsowska B, Skrzypski M, Kubiak M, Błaszak B, Łukomska A, Nowak T, Jaśkowski JM. Expression of Transforming Growth Factor Beta Isoforms in Canine Endometrium with Cystic Endometrial Hyperplasia–Pyometra Complex. Animals. 2021; 11(6):1844. https://doi.org/10.3390/ani11061844
Chicago/Turabian StyleRybska, Marta, Magdalena Woźna-Wysocka, Barbara Wąsowska, Marek Skrzypski, Magdalena Kubiak, Beata Błaszak, Anna Łukomska, Tomasz Nowak, and Jędrzej M. Jaśkowski. 2021. "Expression of Transforming Growth Factor Beta Isoforms in Canine Endometrium with Cystic Endometrial Hyperplasia–Pyometra Complex" Animals 11, no. 6: 1844. https://doi.org/10.3390/ani11061844
APA StyleRybska, M., Woźna-Wysocka, M., Wąsowska, B., Skrzypski, M., Kubiak, M., Błaszak, B., Łukomska, A., Nowak, T., & Jaśkowski, J. M. (2021). Expression of Transforming Growth Factor Beta Isoforms in Canine Endometrium with Cystic Endometrial Hyperplasia–Pyometra Complex. Animals, 11(6), 1844. https://doi.org/10.3390/ani11061844